養殖密度對俄羅斯鱘幼魚的生長、生理和免疫指標的影響
2014-03-17 00:50:44陽李二超陳立僑龍麗娜崔超杜震宇孫盛明黎
水生生物學報 2014年5期
曹 陽李二超陳立僑龍麗娜崔 超杜震宇孫盛明黎 明
(1. 華東師范大學生命科學學院, 上海 200241; 2. 中國水產科學研究院漁業機械儀器研究所, 上海 200092; 3. 中國水產科學研究所淡水漁業研究中心, 無錫 214081; 4. 寧波大學海洋學院, 寧波 315211)
養殖密度對俄羅斯鱘幼魚的生長、生理和免疫指標的影響
曹 陽1李二超1陳立僑1龍麗娜2崔 超1杜震宇1孫盛明3黎 明4
(1. 華東師范大學生命科學學院, 上海 200241; 2. 中國水產科學研究院漁業機械儀器研究所, 上海 200092; 3. 中國水產科學研究所淡水漁業研究中心, 無錫 214081; 4. 寧波大學海洋學院, 寧波 315211)
密度是決定養殖魚類生長速度、產量和效益的主要因素之一。魚類集約化養殖系統中, 生產者往往盡可能地提高單位水體中的放養密度, 以獲得養殖產量和經濟效益的最大化[1]。但是, 密度過高除了直接加劇魚類對空間和餌料的競爭, 導致能量消耗增多和餌料利用率降低外,還會抑制魚類的正常生長, 甚至造成養殖動物的發病和死亡[2—6]。從代謝的角度來看, 在高密度養殖條件下, 魚類新陳代謝旺盛, 血液中的皮質醇、血糖和血紅蛋白水平也相應升高, 血液攜氧能力上升。短期內皮質醇和血糖水平的上升有利于緩解魚體所遭受的脅迫, 但持續過久會導致機體或組織對氨基酸的利用率下降, 并影響免疫系統的正常功能[7—10]。另有研究發現, 養殖系統中密度過高或過低, 都會在一定程度上抑制魚類的生長效率, 從而影響產量和經濟效益。孫大川等在對寶石鱸(Scortum barcoo)的研究發現, 養殖密度過高或者過低都對該魚的生長造成一定的負面影響[7]。已有的研究證實, 魚類種類、規格和養殖條件的不同, 密度脅迫所對其造成的負面影響也不盡相同[11—13]。因此, 探討特定養殖對象在不同密度下的生長性能和生理反應, 對于確定該養殖對象的適宜密度是十分必要的。
俄羅斯鱘(Acipenser gueldenstaedtii), 俗稱金龍王鱘,是一種營底棲生活的冷水性魚類。該魚體型大、生長快、繁殖能力強, 對不良環境具有較高的耐受性, 并且肉質鮮美, 用鱘魚卵所制成的魚子醬具有很高的經濟價值。隨著鱘魚的工廠化養殖業的發展日漸繁盛, 俄羅斯鱘作為鱘魚養殖業的重要品種之一, 具有廣闊的開發前景。然而, 迄今對于俄羅斯鱘適宜養殖密度的研究仍不多見。基于此, 本實驗研究不同養殖密度對俄羅斯鱘幼魚生長、免疫和抗應激性能等方面的差異, 以期為俄羅斯鱘幼魚工廠化養殖適宜密度的確定提供參考, 結果對促進俄羅斯鱘的工廠化養殖技術、提高經濟效益都具有重要的意義。
1 材料與方法
1.1 實驗動物及養殖管理
俄羅斯鱘幼魚購自浙江衢州市一養殖場, 經聚維酮碘(10 g/L浸泡5min后, 暫養14d后, 開始正式實驗。隨機選取體表無外傷、個體均勻的幼魚(4.4±0.5) g共 1090尾, 平均投放到9個網箱(0.75 m×0.75 m×1 m)中。所有網箱均安放在一個循環水池(5 m×1.6 m×1.2 m)中, 水深0.5 m。養殖用水為曝氣24h以上的除氯自來水, 日循環量1440 m3, 日凈化量為120 m3。養殖期全程充氧, 每天用YSI多參數水質分析儀(美國)對溫度、鹽度、溶氧量和pH進行監測。在整個養殖過程中, 水溫(24—26)℃, 溶解氧>6 mg/L, 總氨氮<0.1 mg/L, pH 7.44—8.12, 光照周期保持12h︰12h。實驗周期為56d。
暫養及實驗期間均投喂商業飼料, 營養參數為粗蛋白含量≥48%, 粗脂肪≥5%, 粗灰分≥15%。每天分三次(8:00、14:00和20:00)飽食投喂, 投喂30min后利用食物臺收集殘餌, 記錄投餌量和殘餌量。
1.2 實驗設計和取樣
實驗設置3個密度處理組, 分別為高密度組(3.2 kg/m3)、中密度組(1.6 kg/m3)和低密度組(0.8 kg/m3), 每個處理組設置三個平行。飼養實驗結束后, 饑餓 24h, 以濃度為120 mg/L的三卡因甲基磺酸鹽(MS-222)進行麻醉, 統計魚的存活率, 并稱量體重及測量體長, 以統計增重率和肥滿度。每個平行組隨機挑選 5尾魚, 解剖取肝臟稱重,以計算肝體指數。再隨機挑選10尾魚, 5尾用內涂有肝素鈉(100 IU/mL)的一次性注射器于尾靜脈取血, 用以血液學和免疫學分析, 另 5尾用未浸潤過肝素鈉的一次性注射器尾靜脈取血, 所取血樣置于 4℃過夜, 1200 r/s離心5min獲取血清, –80℃存儲用于生化分析。
與生長指標有關的指標計算公式如下:
增重率(Weight gain, WG, %)=100×(W2–W1)/W1
餌料系數(Feed conversion ratio, FCR)=F/[W2–W1+W0]攝食量(Food intake, FI, g/尾)=F/[(N2+N1)/2]
蛋白質效率(Protein efficiency rate, PER, %)=100×(W2–W1)/(F×48%)
肥滿度(Condition factor, K)=WB/L3
肝體指數(Hepatosomatic index, HSI, %)=100×WL/WB存活率(Survival rate, SR, %)=100×N2/N1
其中, W1、W2和W0分別為實驗開始、實驗結束和死亡個體總重量(g), F為各平行投喂的飼料量(g), N1為初始總數(尾), N2為終末總數(尾), WB為個體體重(g), L為體長(cm), WL為同一尾魚的肝臟鮮重(g)。
1.3 急性氨氮脅迫
在養殖實驗結束后, 每個平行隨機選取10尾俄羅斯鱘, 分別投放于9個聚乙烯(0.6 m×0.4 m×0.4 m)水箱中,進行 96h氨氮急性脅迫實驗。實驗用水和養殖實驗相同,期間不投喂。
參照杜浩等[14]對中華鱘稚魚的研究結果并結合多次預實驗的結果, 確定用總氨17 mg/L的濃度進行96h氨氮脅迫實驗, 每隔7h進行氨氮濃度的校準。統計12h、24h、36h、48h、60h、72h、84h和96h的死亡率, 及時撈出死魚。
1.4 血液指標分析
全血樣品一式三份經磷酸緩沖液(PBS)稀釋(1︰10000), 采用血球計數板統計紅細胞數(RBC)和白細胞數(WBC)。血紅蛋白含量的測定采用雙波長比色法, 試劑盒購自南京建成生物工程研究所。
1.5 生化指標測定
皮質醇、超氧化物歧化酶(SOD)和丙二醛(MDA)的測定均選用南京建成生物工程研究所的試劑盒。皮質醇采取雙位點夾心酶聯免疫吸附法。血糖含量測定采用葡萄糖氧化酶-過氧化物酶法, 用分光光度計測定在波長505 nm處吸光值。超氧化物歧化酶(SOD)酶活性測定采用黃嘌呤氧化酶法, 其活力單位定義為: 每mg蛋白質在1 mL反應液中SOD 抑制率達50%時所對應的SOD 量為一個亞硝酸鹽單位(NU)。丙二醛(MDA)含量采用硫代巴比妥酸法, 利用MDA可與硫代巴比妥酸縮合形成紅色產物, 測定其在532 nm處的吸光值。每個樣品均重復測定三次。
1.6 免疫指標
溶菌酶的測定采用南京建成生物研究室的試劑盒,遵照空白對照法, 同一樣品一式三份測定530 nm處的吸光值, 以1min內吸光值下降0.001為1個活性單位(U)。
血液吞噬細胞的呼吸暴發水平(Respiratory burst activity)的測定方法參照[15]。同一樣品重復測定三次。向每個酶標孔中加入 0.1 mL的硝基四唑氮蘭(Nitro blue tetrazolium, NBT)溶液(sigma公司)0.1 mL, 再向當中加入0.1 mL的抗凝血, 使其在NBT溶液中溫浴至少30min, 然后加入50 μL的漢克緩沖液(Sigma公司), 3000 r/min離心5min, 記錄其在545 nm下的吸光值。根據NBT(mg/mL)= 40×(吸光值–0.0245)/5.8564得出呼吸暴發的水平。
1.7 統計分析
所有數據用平均值±標準誤(Mean±SE)的方式來表示。采用單因素方差分析(ANOVA)對實驗數據進行檢測,如果檢測到顯著性差異, 則采用 Duncan多重比較。P<0.05時被認為差異顯著。軟件在Windows 7操作系統中完成。
2 結果
2.1 生長性能
由表 1可以看出, 各組存活率無顯著差異(P>0.05);中密度組的增重率和肥滿度均顯著高于高密度組(P<0.05), 但與低密度組之間無顯著差異(P>0.05); 同時,低、中密度組之間的蛋白質效率差異不顯著(P>0.05), 但均顯著高于高密度組(P<0.05); 飼料系數的統計結果顯示,高密度組顯著高于中、低密度組(P<0.05), 后二者間則無顯著性差異(P>0.05); 中密度組攝食量顯著高于低密度組和高密度組(P<0.05), 且高密度組顯著高于低密度組(P<0.05); 此外, 高密度組的肝指數最低, 但在各組間無顯著性差異(P>0.05)。
2.2 血液參數
從表 2中可以看出, 高密度組的血紅蛋白含量最高(P<0.05), 相比之下, 中密度組與低密度組之間則無顯著性差異(P>0.05); 此外, 紅細胞數目隨著養殖密度的提高而升高, 但各個密度組之間并無顯著性差異(P>0.05); 相似地, 高密度組的白細胞數目最低, 但與中密度組、低密度組相比, 三者之間亦無顯著性差異(P>0.05)。高密度組呼吸爆發水平最低, 但各組間無顯著差異(P>0.05)。X SE± )X SE± )
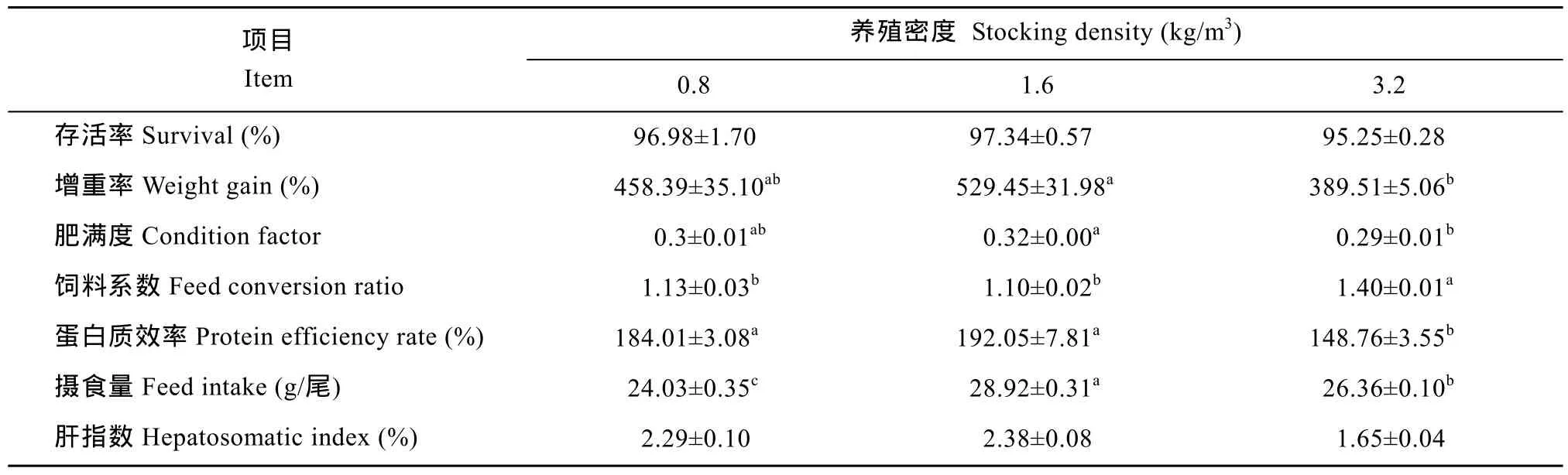
表1 不同養殖密度對俄羅斯鱘的生長指標的影響Tab. 1 Effects of stocking density on growth parameters of juvenile Russian sturgeon juvenile (n=5,
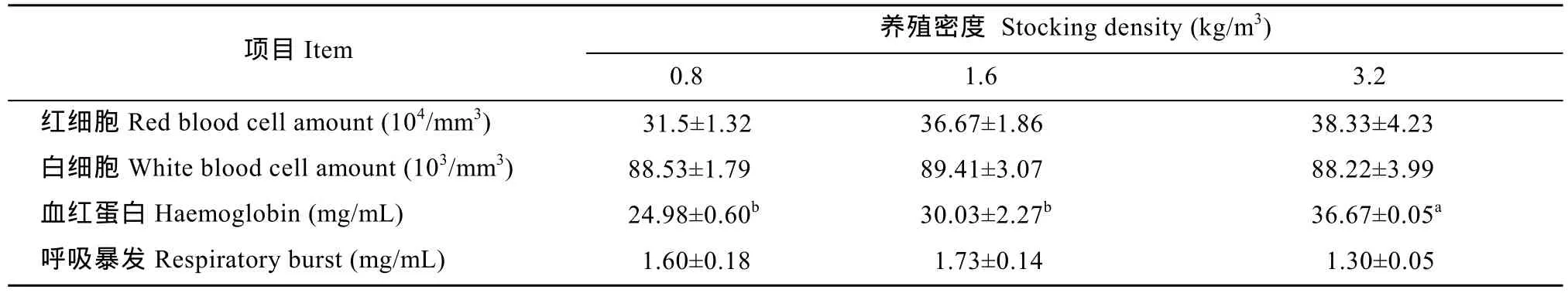
表2 不同養殖密度對俄羅斯鱘血液參數的影響Tab. 2 Effect of stocking density on some hematological parameters in juvenile Russian sturgeon juvenile (n=5,
2.3 血清生化和免疫參數
表 3顯示, 高密度組的皮質醇水平顯著高于中密度組、低密度組(P<0.05), 而低密度組和中密度組無顯著性差異(P>0.05); 血糖含量的趨勢與皮質醇類似, 但各組間無顯著差異(P>0.05); 低密度組的血清SOD活性最高, 高密度組的最低, 但三個密度組之間無顯著性差異(P>0.05);血清MDA的測定結果顯示, 高密度組血清MDA的含量最高, 中密度組的最低, 各組間無顯著性差異(P>0.05);此外, 低密度組與中密度組的溶菌酶活力無顯著性差異(P>0.05), 但均顯著高于高密度組(P<0.05)。
2.4 急性氨氮脅迫
從圖1可以看出, 在總氨濃度為17 mg/L條件下, 隨著氨氮脅迫時間的延長, 各組死亡率均呈逐漸上升趨勢,從實驗開始至 84h內, 同一時段不同組間之間的個體死亡率無顯著性差異(P>0.05), 但至 96h時, 高密度組的死亡率最高, 低密度組居中, 中密度組最低, 統計分析發現,高密度組的死亡率顯著高于中密度組(P<0.05), 而低密度組與中、高密度組之間則無顯著性差異(P>0.05)。
3 討論
3.1 生長性能
在通常情況下, 隨著養殖密度的上升, 魚類的生長率和飼料效率均呈下降的趨勢[16]。Rafatnezha等對不同密度條件下歐洲鰉(Huso huso)的研究表明, 最高密度組的個體增重率和肥滿度顯著最低, 而飼料系數最高[8]。Christiansen等在對北極紅點鮭(Salvelinus alpinus)的研究發現, 攝食率和增重率均隨密度的升高而顯著降低[17]。并且,過高的養殖密度也在一定程度上抑制了銀鱸(Bidyanus bidyanus)、非洲鯰(Clarias gariepinus)、羅非魚(Oreochromis spilurus)和虹鱒(Oncorhynchus mykiss)等的生長[18—21]。高密度抑制魚類生長的原因有如下幾點: 其一, 過高的養殖密度會加劇群體內的競爭, 直接導致平均個體攝食量降低[22]。其二, 密度過高會對魚類造成一定程度的應激效應, 機體需消耗更多的能量來應對脅迫,以維持正常的代謝過程。在本實驗中, 高密度組的血紅蛋白含量顯著高于中、低密度組, 這與此前諸多報道的結果一致, 提示高密度組個體的血液攜氧能力提高, 新陳代謝相對旺盛[23—25]。其三, 高密度下的個體用于新陳代謝所消耗能量的比例,一般要高于密度適中或較低環境下的個體[26]。并且, 高密度組個體為了獲得必要的食物, 覓食游動頻繁, 消耗的能量也相應有所提高[27]。在本實驗中, 與中、低密度組相比, 高密度組的攝食量、蛋白質效率、飼料利用率均最低,表明高密度組條件下的魚攝入、吸收和同化的營養物質比例相對較低, 而消耗供能的比例偏高, 從而抑制了魚的生長性能和飼料利用效率。但也有一些學者發現, 高密度并未明顯影響某些魚類的生長性能, 如 Celikkale等的研究顯示, 高養殖密度對均重(280±31.27) g的俄羅斯鱘的特定生長率、飼料利用率和肥滿度未造成顯著影響[28]。造成這一差異的原因, 可能與魚的種類、生長階段以及實驗環境的不同有關, 具體原因尚需進一步深入研究。另有研究指出, 養殖密度偏低亦會抑制魚的生長[29]。逯尚尉等在對點帶石斑魚(Epinephelus malabaricus)的研究表明,中密度(2.1 kg/m3)組個體的增重率顯著高于低密度(1.1 kg/m3)組和高密度(3.2、4.2 kg/m3)組[5]。本研究中低密度組的攝食率和增重率均低于中密度組, 可能由于俄羅斯鱘幼魚具有集群效應(Cluster effect), 密度過低導致其攝食不積極, 且個體平均活動范圍過大, 覓食等活動所消耗能量相應增多。低密度組的飼料系數高于中密度組, 且蛋白質效率較低則從另一個側面驗證了這一點。
表3 不同養殖密度對俄羅斯鱘血清生化和免疫參數的影響Tab. 3 Effects of stocking density on the serum biochemical and immune parameters in juvenile Russian sturgeon juvenile (n=5,± SE)

表3 不同養殖密度對俄羅斯鱘血清生化和免疫參數的影響Tab. 3 Effects of stocking density on the serum biochemical and immune parameters in juvenile Russian sturgeon juvenile (n=5,± SE)
項目Item 0.8 1.6 3.2養殖密度 Stocking density (kg/m3)血清皮質醇水平Serum cortisol (ng/mL) 728.62±24.58b731.73±45.74b887.32±46.16a血糖水平Serum glucose (mg/dL) 45.05±1.28 46.10±1.41 52.08±3.35超氧化物歧化酶SOD (U/mL) 98.79±2.06 97.50±3.11 91.40±2.28丙二醛MDA (nmol/mL) 6.83±0.43 6.20±0.19 7.11±0.27溶菌酶Lysozyme (U/mL) 155.79±2.11a179.61±9.5a117.99±7.81b
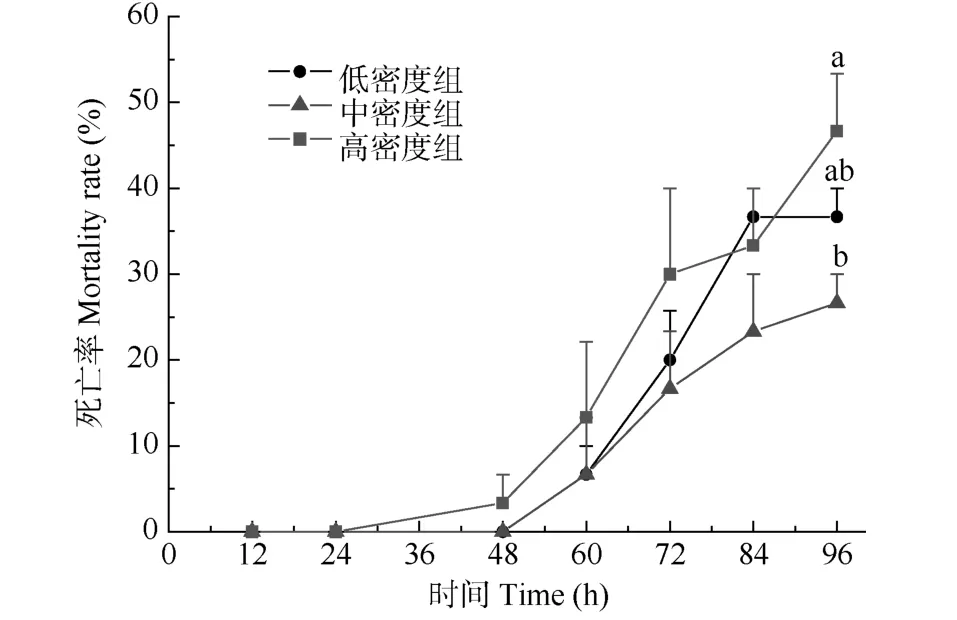
圖1 俄羅斯鱘幼魚氨氮脅迫死亡率Fig. 1 The mortality rate of Russian sturgeon juvenile under ammonia nitrogen stress
3.2 抗應激能力
皮質醇(Cortisol)是一項反映評估魚類應激反應強度的重要指標。當魚類處于急性應激狀態下, 魚體的皮質醇在幾小時之內急劇升高, 而處于慢性脅迫條件時, 皮質醇水平長期維持在較高水平。魚類由于高密度脅迫而長期處在應激狀態, 通過下丘腦—垂體—腎間組織產生皮質醇等類固醇, 釋放到血液中[30]。皮質醇短期的升高能夠促進體蛋白分解, 加速脂肪的氧化, 糖類的合成增多, 從而使機體獲得足夠的能量用以緩解抵御脅迫所需。但是, 皮質醇水平過高或長期持續較高水平, 則會造成魚體生長停滯, 脂肪合成減少和機體消瘦[31]。在本研究中, 高密度組個體長期處于某種應激狀態, 因此機體皮質醇持續處于較高水平, 這可能是抑制幼魚生長的原因之一。并且, 密度脅迫造成的皮質醇水平的升高還會引起魚類短期的急性適應性反應, 血糖水平因而升高[32]。在本研究中, 高密度組的血糖含量高于其他組, 但組間沒有達到顯著差異水平。Lupatsch等對歐洲鱸魚(Dicentrarchus labrax)[26]的研究支持了本實驗的結果。不僅如此,機體持續處于應激狀態導致了皮質醇水平長期過高, 進而抑制機體的特異性和非特異性免疫防御機制, 導致魚體對各類病原體敏感性的升高[33]。高養殖密度常引起水質惡化, 氨氮濃度過高, 進而導致魚類組織潰爛和呼吸功能受阻, 損傷魚類的抗氧化系統和免疫系統等[34]。本研究 96h急性氨氮脅迫實驗中, 高密度組的累計死亡率高于中、低密度組, 可能是由于密度脅迫導致了皮質醇水平過高, 進而降低了該組魚對高氨氮的耐受性和免疫性能的正常發揮。
3.3 免疫及抗氧化力
有研究表明, 高養殖密度作為脅迫會對魚類免疫性能和抗氧化性能造成一定影響[35,36]。溶菌酶是魚類重要的非特異性免疫因子。Demers和 Bayne在對虹鱒(Oncor-hynchus mykiss)的研究發現, 隨著養殖密度的升高, 皮質醇和溶菌酶水平顯著升高[37]。但 Yin等對錦鯉(Cyprinus carpio L.)研究的發現, 溶菌酶等非特異性免疫指標在脅迫初期顯著降低, 之后便維持較低水平[38]。王博文等在對鯽魚(Carassius auratus)的研究表示, 各密度組溶菌酶水平的變化趨勢與皮質醇不同, 高密度組的溶菌酶水平隨著脅迫時間的延長, 呈先升高后降低的趨勢[39]。在本研究中高密度組的溶菌酶水平顯著低于中、低密度組。在環境影響機體免疫器官的過程中皮質醇起介導作用,短期內的升高能幫助增強機體非特異性免疫, 但若長期處于較高水平就會抑制機體免疫功能[37]。這可能是導致本實驗高密度組俄羅斯鱘的溶菌酶水平受到顯著抑制,免疫功能下降的一個原因。呼吸暴發(Respiratory Burst, NBT)是另一個反映機體免疫性能的指標, 呼吸暴發活性越強, 說明吞噬細胞的殺菌能力越強[40]。在本研究中, 各密度下的俄羅斯鱘的呼吸暴發活性雖然差異不顯著, 但高密度組的 NBT活性最低, 這說明高密度脅迫在一定程度上影響了鱘魚的免疫性能, 該結果與Peters等的研究結果類似[41]。SOD活力和MDA含量的高低間接反映了機體的抗氧化能力[42]。本研究的結果顯示, 高密度組的血清SOD活性要低于中、低密度組, 且高密度組的MDA水平要稍高于中、低組, 組間差異均不顯著, 與劉松巖等[36]對中華鱘(Acipensersinensis)的研究結果相似, 提示高密度組個體受到密度脅迫時, 新陳代謝加快, 能量消耗升高, 這在一定程度上促進了氧自由基的產生, 進而導致MDA含量升高。高密度組的 SOD活力較其他組低, 說明養殖密度過高也可能導致機體內的自由基代謝失衡。
4 結論
本實驗初步研究表明, 低密度(0.8 kg/m3)組和中密度(1.6 kg/m3)組條件下的俄羅斯鱘幼魚的生長、生理及免疫性能較高密度(3.2 kg/m3)組更優。其中中密度組最優。在本實驗條件下, 體質量為(4—5) g左右的俄羅斯鱘幼魚的適宜放養密度約為 1.6 kg/m3, 該結果可為工廠化養殖提供參考依據。
[1] Zhu Y J, Yang D G, Chen J W, et al. An evaluation of stocking density in the cage culture efficiency of Amur sturgeon (Acipenser schrenckii) [J]. Journal of Applied Ichthyology, 2011, 27(2): 545—549
[2] Salas-Leiton E, Anguis V, Martin-Antonio B, et al. Effects of stocking density and feed ration on growth and gene expression in the senegalese sole (Solea senegalensis): potential effects on the immune response [J]. Fish & Shellfish Immunology, 2010, 28(2): 296—302
[3] Marchand F, Boisclair D. Influence of fish density on the energy allocation pattern of juvenile brook trout (Salvelius fontinalis) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(4): 796—805
[4] Shi X T, Li D P, Zhuang P, et al. Effects of rearing density on juvenile Acipenser schrenckii digestibility, feeding rate and growth [J]. Chinese Journal of Applied Ecology, 2006, 17(8): 1517—1520 [石小濤, 李大鵬, 莊平, 等. 養殖密度對史氏鱘消化率, 攝食率和生長的影響. 應用生態學報, 2006, 17(8): 1517—1520]
[5] Lu S W, Liu Z P, Yu Y. Effects of density stress on growth and metabolism of juvenile Epinephelus malabaricus [J]. Journal of Fishery Sciences of China, 2010, 18(2): 322—328 [逯尚尉, 劉兆普, 余燕. 密度脅迫對點帶石斑魚幼魚生長、代謝的影響. 中國水產科學, 2011, 18(2): 322—328]
[6] Han N N, Shi Y C. The application of blood indexes in ichthyological research [J]. Journal of Anhui Agricultural Science, 2010, 38(33):18877—18878, 18880 [韓娜娜, 史成銀. 血液指標在魚類學研究中的應用. 安徽農業科學, 2010, 38(33): 18877—18878, 18880]
[7] Tian Z H, Xu S G, Wang W, et al. Effect of acute thermal stress on Hsp70 Mrna, physiology and nonspecific immunity in Siberian sturgeon (Acipenser baerii) [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 344—350 [田照輝, 徐邵剛, 王巍, 等. 急性熱應激對西伯利亞鱘HSP70 mRNA表達、血清皮質醇和非異性免疫的影響. 水生生物學報, 2013, 37(2): 344—350]
[8] Rafatnezhad S, Falahatkar B, Gilani M H T. Effects of stocking density on haematological parameters, growth and fin erosion of great sturgeon (Huso huso) juveniles [J]. Aquaculture Research, 2008, 39(14): 1506—1513
[9] Pickering A D. Rainbow trout husbandry: management of the stress response [J]. Aquaculture, 1992, 100(1—3): 125—139
[10] Huan Y, Zhu X M, Xie S Q, et al. Compensmory growth in Acipenser baerii, following feed derivation at high tempemture: temporal patterns in growth, feed intake and body composition [J]. Acta Hydrobiologica Sinica, 2011, 34(6): 1113—1121 [黃瑩, 朱曉鳴, 解綬啟, 等. 西伯利亞鱘在高溫下饑餓后的補償生長. 水生生物學報, 2011, 34(6): l113—1121]
[11] Rotllant J, Tort L. Cortisol and glucose responses after acute stress by net handling in the sparid red porgy previously subjected to crowding stress [J]. Journal of Fish Biology, 1997, 51(1): 21—28
[12] Peng S M, Shi Z H, Sun P, et al. Effects of breeding density on the growth and tissues biochemical indices of juvenile silver pomfret (Pampus argenteus) [J]. Chinese Journal of Ecology, 2010, 29(7): 1371—1376 [彭士明, 施兆鴻, 孫鵬,等. 養殖密度對銀鯧幼魚生長及組織生化指標的影響. 生態學雜志, 2010, 29(7): 1371—1376]
[13] Yang D, Wei Q W, Wang K, et al. Three types of culture Chinese sturgeon (Acipenser sinesis) and the analysis of their effects by comparison [J]. South China Fisheries Science, 2006, 2: 1—5
[14] Du H, Wei Q W, Liu J Y, et al. Acute toxicity of phenol, copper, nitrite and total ammonia to juvenile Chinese sturgeon, Acipenser sinensis [J]. Journal of Dalian FisheriesUniversity, 2007, 22(2): 118—122 [杜浩, 危起偉, 劉鑒毅,等. 苯酚、Cu2+、亞硝酸鹽和總氨氮對中華鱘稚魚的急性毒性. 大連水產學院學報, 2007, 22(2): 118—122]
[15] Prusty A K, Sahu N P, Pal A K, et al. Effect of dietary tannin on growth and haemato- immunological parameters of Labeo rohita (Hamilton) fingerlings [J]. Animal Feed Science and Technology, 2007, 136: 96—108
[16] Shelbourn J E, Brett J R, Shirahata S. Effect of temperature and feeding regimes on the specific growth rate of sockeye salmon fry (Oncorhynchus nerka), with a consideration of size effect [J]. Fisheries Research Board of Canada, 2011, 30: 1191—1194
[17] Christiansen J S, Svendsen Y S, Jobling M, et al. The combined effects of stocking density and sustained exercise on the behaviour, food intake, and growth of juvenile Arctic charr (Salvelinus alpinus L.) [J]. Canadian Journal of Zoology, 1992, 70(1): 115—122
[18] Rowland S J, Mifsud C, Nixon M, et al. Effects of stocking densety on the performance of the Australian freshwater silver perch (Bidyanus bidyanus) in cages [J]. Aquaculture, 2006, 253(1—4): 301—308
[19] Haylor G S. Controlled hatchery production of Clarias gariepinus (Burchell, 1822): growth and survival of fry at high stocking density [J]. Aquaculture Research, 1992, 23(3): 405—422
[20] Cruz E M, Ridha M. Production of the tilapia Oreochromis spilurus Günter stocked at different densities in sea cages [J]. Aquaculture, 1991, 99: 95—103
[21] Bagley M J, Bentley B, Gall A E G. A genetic evaluation of the influence of stocking density on the early growth of rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 1994, 121(4): 313—326
[22] Sawcer S J, Maranian M, Singlehurst S, et al. Enhancing linkage analysis of complex disorders: an evaluation of highdensity genotyping [J]. Human Moecularl Genetics, 2004, 13(7): 1943—1949
[23] Trenzado C E, Morales A E, Palma J M, et al. Blood antioxidant defenses and hematological adjustments in crowded uncrowded rainbow trout (Oncorhynchus mykiss) fed on diets with different levels of antioxidant vitamins and HUFA [J]. Comparative Biochemistry and Physiology C, Toxicology & Pharmacology, 2009, 149(3): 440—447
[24] Ruane N M, Carballo E C, Komen J. Increased stocking density influences the acute physiological response of the common carp Cyrinus carpio (L) [J]. Aquaculture Research, 2002, 33(10): 777—784
[25] Docan A, Gristea V, Grecu I, et al. Haematological response of the European catfish, Silurus glanis reared at different densities in “flow-through” production system [J]. Archiva Zootechnica, 2010, 13: 63—70
[26] Lupatsch I, Santos G A, Schrama J W, et al. Effect of stocking density and feeding level on energy expenditure and stress responsiveness in European sea bass Dicentrarchus labrax [J]. Aquaculture, 2010, 298(1): 245—250
[27] Lambooij E, Vis J W V D, Kloosterboer R J, et al. Welfare aspects of live chilling and freezing of farmed eel (Anguilla anguilla L.): neurological and behavioural assessment [J]. Aquaculture, 2002, 210(1—4): 159—169
[28] Celikkale M S, Memis D, Ercan E, et al. Growth performance of juvenile Russian sturgeon (Acipenser gueldenstaedtii Brandt & Ratzenburg,1833) at two stocking densities in net cages [J]. Journal of Applied Ichthyology, 2005, 21(1): 14—18
[29] Jorgensen E H, Christiansen J S, Jobling M. Effects of stocking density on food intake, growth performance and oxygen consumption in Arctic charr (Salvelinus alpinus) [J]. Aquaculture, 1993, 110: 191—204
[30] Vijayan M M, Leatherland J F. High stocking density affects cortisol secretion and tissue distribution in brook charr, Salvelinus fontinalis [J]. Journal of Endocrinology, 1990, 124(2): 311—318
[31] Gregory T R, Wood C M. The effects of chronic plasma cortisol elevation on the feeding behaviour, growth, competitive ability, and swimming performance of juvenile rainbow trout [J]. Physiological and Biochemical Zoology, 1999, 72(3): 286—295
[32] Bayunova L, Barannikova I, Semenkova T. Sturgeon stress reactions in aquaculture [J]. Journal of Applied Ichthyology, 2002, 18(4—6): 397—404
[33] Robertson O H, Hane S, Wexler D C, et al. The effect of hydrocortisone on immature rainbow trout (Salmo gairdneri) [J]. General and Comparative Endocrinology, 1963, 3(4—6): 422— 436
[34] Qiang J, Xu P, He J, et al. The combined effects of external ammonia and crowding stress on growth and biochemical activities in liver of (GIGT) Nile tilapia juvenile (Oreochromis niloticus) [J]. Journal of Fisheries of China, 2011, 35(12): 1837—1847 [強俊, 徐跑, 何杰, 等. 氨氮與擁擠脅迫對吉富品系尼羅羅非魚幼魚生長和肝臟抗氧化指標的聯合影響. 水產學報, 2011, 35(12): 1837—1848]
[35] Hegazi M M, Attia Z I, Ashour O A. Oxidative stress and antioxidant enzymes in liver and white muscle of Nile tilapia juveniles in chronic ammonia exposure [J]. Aquatic Toxicology, 2010, 99(2): 118—125
[36] Liu S Y. Effect of environmental stress on free radical level and antioxidant enzyme activities of Chinese sturgeon, Acipenser Sinensis [D]. Thesis for Master of Science. Huazhong Agricultural University, Wuhan. 2006 [劉松巖.環境脅迫對中華鱘體內自由基水平和抗氧化酶活力的影響. 碩士學位論文, 華中農業大學, 武漢. 2006]
[37] Demers N E, Bayne C J. The immediate effect of stress on hormones and plasma lysozyme in rainbow trout [J]. Developmental and Comparative Immunology, 1997, 21(4): 363—373
[38] Yin Z, Lam T J, Sin Y M. The effects of crowding stress on the non- specific immuneresponse in fancy carp (Cyprinus carpio L.) [J]. Fish and Shellfish Immunology, 1995, 5(7): 519—529
[39] Wang B W, Wang J G, Li A H, et al. Changes of cortisol an lysozyme levels in Carassius auratus blood after crowding stress and the fish sensitivity to pathogen [J]. Journal of Fishery Sciences of China, 2004, 11(4): 408—412 [王博文,王建國, 李愛華, 等. 擁擠脅迫后鯽魚血液皮質醇和溶菌酶水平的變化及對病原的敏感性. 中國水產科學, 2004, 11(4): 408—412]
[40] Sharp G J E, Secombes C J. The role of reactive oxygen species in the killing of the bacterial fish pathogen Aeromonas salmonicida by rainbow trout macrophages [J]. Fish and Shellfish Immunolog, 1993, 3(2): 119—129
[41] Peters G, Faisal M, Lang T, et al. Stress caused by social interaction and its effect on susceptibility to Aeromonas hydrophila infection in rainbow trout Salmo gairdneri [J]. Diseases of Aquatic Organisms, 1988, 4: 83—89
[42] Viarengo A, Canesi L, Martinez P G, et al. Pro-oxidant processes and antioxidant defence systems in the tissues of the Antarctic scallop (Adamussium colbecki) compared with the Mediterranean scallop (Pecten jacobeus) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1995, 111(1): 119—126
EFFECTS OF STOCKING DENSITY ON GROWTH, PHYSIOLOGICAL AND IMMUNE RESPONSES IN JUVENILE RUSSIAN STURGEON
CAO Yang1, LI Er-Chao1, CHEN Li-Qiao1, LONG Li-Na2, CUI Chao1, DU Zhen-Yu1, Sun Sheng-Ming3
and LI Ming4
(1. School of Life Sciences, East China Normal University, Shanghai 200241, China; 2. Institute of Fisheries Mechanical Equipment, Scientific Research Institute of China, Shanghai 200092, China; 3. Freshwater Fisheries Research Center, Chinese
Academy of Fishery Sciences, Wuxi 214081, China; 4. School of Marine, Ningbo University, Ningbo 315211, China)
俄羅斯鱘; 養殖密度; 生長; 生理; 免疫
Russian sturgeon (Acipenser gueldenstaedtii); Stocking density; Growth; Physiology; Immune
S965.2
A
1000-3207(2014)05-0968-07
10.7541/2014.143
2013-07-30;
2014-02-26
國家科技支撐計劃(2012BAD25B03); 國家基礎研究“973” 計劃項目(2009CB118702); 公益性行業(農業)科研專項項目(201003020, 201203065); 國家自然科學基金項目(31172422); 上海市教委水產E研究院(E03009)部分資助
曹陽(1988—)女, 遼寧開原人; 碩士研究生; 主要從事水生動物營養學研究。E-mail: caoyang3640@163.com
陳立僑, 教授; E-mail: lqchen@bio.ecnu.edu.cn
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52

- 水生生物學報的其它文章
- GILL MEDIATES IMMUNE RESPONSES AFTER GRASS CARP REOVIRUS CHALLENGE IN GRASS CARP (CTENOPHARYNGODON IDELLA)
- MORPHOLOGICAL OBSERVATION AND RBCL SEQUENCE ANALYSIS OF A NEW SPECIES FROM CHINA, GRATELOUPIA BOAOENSIS WANG ET LUAN SP. NOV.(HALYMENIACEAE, RHODOPHYTA)
- GILL MEDIATES IMMUNE RESPONSES AFTER GRASS CARP REOVIRUS CHALLENGE IN GRASS CARP (CTENOPHARYNGODON IDELLA)
- 一種快速構建集胞藻6803 petBD必需基因定點突變株的方法
- 大通湖環棱螺的次級生產力
- 黃鯽線粒體DNA控制區結構及長度多態性分析