Fe3+對小球藻的生長及油脂含量的影響
2014-03-17 06:56:08孫遠劉文彬周鐵柱謝通慧梁斌張永奎
生物技術通報 2014年4期
孫遠 劉文彬 周鐵柱 謝通慧 梁斌 張永奎
(四川大學化學工程學院制藥與生物工程系,成都 610065)
Fe3+對小球藻的生長及油脂含量的影響
孫遠 劉文彬 周鐵柱 謝通慧 梁斌 張永奎
(四川大學化學工程學院制藥與生物工程系,成都 610065)
傳統化石能源儲量日益減少,生物柴油因其環保可再生性成為優質的石化柴油替代品。利用小球藻生產生物柴油速度快、油脂含量高,受到了廣泛關注。為進一步提高小球藻生產生物柴油效率,分別探究了Fe3+的濃度及添加時間對自養和異養小球藻生長及產油的影響,獲得最優Fe3+培養條件為:自養小球藻延滯期添加10-3g/L Fe3+,生物量及油脂含量達2.80 g/L及30.90%;異養小球藻指數期添加10-5g/L Fe3+,生物量及油脂含量達3.30 g/L及29.05%。經脂肪酸分析,以上條件獲得的微藻油脂均可作為生物柴油生產原料。
小球藻 Fe3+自養 異養 生長 油脂
傳統化石能源儲量的日益減少,溫室氣體排放的加劇及油價的飆升使得全球對尋找可再生燃料的關注度越來越高[1]。生物柴油可減少57%-86%的溫室氣體排放,被廣泛研究及開發。至今,生物柴油的原料主要來自油料作物(如大豆、玉米、麻瘋樹等)、動物脂肪和餐飲廢油[2],由這些原料生產的生物柴油成本較高,且遠遠不能滿足運輸所需。微藻作為高光合生物有其特殊的原料成本優勢,以微藻作為載體生產的可再生生物柴油可以滿足全球需求[3]。微藻發酵具有生長速度快、油脂含量高、不與糧爭田、環保等優點[5]。并且,微藻生物燃料無毒,可生物降解[6]。
微藻細胞可進行自養和異養生長,自養微藻利用CO2進行光合作用,可在一定程度減少溫室氣體的排放,利于環境保護。但自養微藻產量較低[7],對光照及空氣含量具有依賴性。異養微藻可同時實現高細胞濃度及油脂含量[8],并且不依賴于光照和空氣。但其昂貴的原料成本制約著異養微藻生產商業化[9]。在自然條件下微藻中的油脂含量,一般是生物量的14%-30%(W/W)[9,11]。
大量研究表明,眾多無機元素,如氮、磷、鉀、鎂、鈣、硫、鐵、銅、錳及鋅等都對微藻生長有
著一定的影響。此外,氯、鉬等微量元素對微藻的生命活動也起著重要作用,常常被包括在藻類的微量元素配方之中。經過12年的現場試驗研究,Behrenfeld等[12]發現在高氮低葉綠素及貧營養水域中,鐵對浮游植物的生長與油脂積累起到了關鍵作用。很多科學家也對其機理做了研究,但遺憾的是至今科學界還沒有得出一個公認的結果,但普遍認為可能是鐵離子在脂質積累過程中調節了某些與脂質合成相關的酶的表達,主要是Fe3+能影響細胞色素氧化酶的鐵卟啉的合成。因此,本研究主要進行Fe3+對小球藻自養和異養生長及產油影響的分析,獲得利于小球藻自養和異養生長及產油的最優Fe3+濃度及添加時間,旨在為進一步提高小球藻生產生物柴油效率奠定基礎。
1 材料與方法
1.1 材料
1.1.1 試樣藻種 本課題組前期從水樣中篩選獲得的普通小球藻(Chlorella vulgaris)[4]。
1.1.2 培養基 種子液培養在添加了10 g/L葡萄糖的SE培養基中,其組成如下:0.75 g/L NaNO3,0.025 g/L CaCl2·2H2O,0.075 g/L MgSO4·7H2O,0.075 g/L K2HPO4·3H2O,0.175 g/L KH2PO4,0.025 g/L NaCl,0.005 g/L FeCl3·6H2O,1 mL/L Fe-EDTA,1 mL/L A5,40 mL/L Soil extract。發酵培養基為改良的SE培養基,根據試驗設計含有不同鐵離子濃度(Fe-EDTA)。自養培養基中不含有葡萄糖,異養培養基中含有10 g/L葡萄糖,接種量均有10%(V/V)。
1.1.3 培養條件 取新鮮培養基裝入100 mL/250 mL三角瓶中,高壓滅菌后(115℃,30 min),接入種子液,轉速180 r/min,溫度28℃。
1.2 方法
1.2.1 生長情況測定
1.2.1.1 光密度法 利用紫外/可見分光光度計測定680 nm處藻液的吸光度值(OD值),蒸餾水作參比。根據藻液的OD680值監測微藻細胞的生長狀況。
1.2.1.2 細胞干重法 發酵結束后,取一定體積藻液將藻液于4 800 r/min下離心10 min,水洗3次,去除上清液。烘干下層藻體至恒重,獲得微藻細胞的生物量(g/L)。
1.2.2 油脂含量及脂肪酸含量測定 發酵結束后,對干藻粉進行傳統的氯仿-甲醇提取[13],細胞內部油脂被提取出來,從而獲得小球藻油脂產量(g/L)。并根據下列公式計算得到小球藻油脂含量:
油脂含量(%)=油脂產量(g/L)/生物量(g/L)
提出的微藻油脂甲酯化后進行氣質聯用(GC/MS-QP2010,Shimadzu,Japan)分析其脂肪酸成分[4,13]。
1.2.3 統計學分析 所有試驗設有3個平行樣,計算平均值后繪制曲線進行分析,誤差通過平行樣與平均值的偏差獲得。試驗結果進行Origin 8的ANOVA顯著分析。
2 結果
2.1 Fe3+對小球藻自養生長及產油的影響
2.1.1 Fe3+濃度對小球藻自養生長及產油的影響為探究Fe3+對小球藻自養培養條件下的生長及產油的影響情況,小球藻在含有10-1、10-3、10-5和10-7g/L Fe3+的SE培養基中的自養生長被監測,結果(圖1)顯示,低濃度的Fe3+(10-3、10-5和10-7g/L)有利于小球藻的生長,并且Fe3+對小球藻生長的促進作用在此濃度范圍內(10-7-10-3g/L)隨濃度的增加而增強。當Fe3+濃度達到10-3g/L時,對小球藻生長的促進作用不再增強,反而出現抑制作用,小球藻在10-3g/L Fe3+培養條件下的生長甚至低于培養基中無Fe3+的情況。
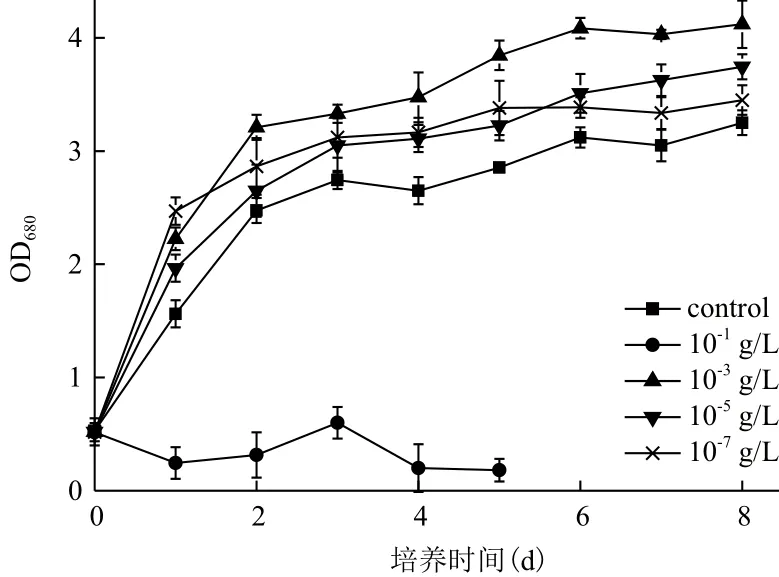
圖1 Fe3+濃度對小球藻自養生長的影響
發酵結束后,比較不同Fe3+濃度培養條件下小球藻的生長及油脂含量可知,微藻細胞密度、生物量及油脂含量均在10-3g/L Fe3+培養條件下取得最大
值(表1)。此培養條件下,小球藻生物量達到2.80 g/L,約為不含Fe3+培養條件下小球藻生物量的1.04倍。關于小球藻的油脂含量,在10-3g/L Fe3+自養培養條件下高達30.90%,較不含Fe3+培養條件下其油脂含量提高了42.66%。Fe3+濃度不同,小球藻油脂含量出現了顯著的區別,而且Fe3+對小球藻油脂含量的增強幅度大于Fe3+對小球藻生物量的增加幅度,說明Fe3+對小球藻的油脂積累有較大的影響作用。
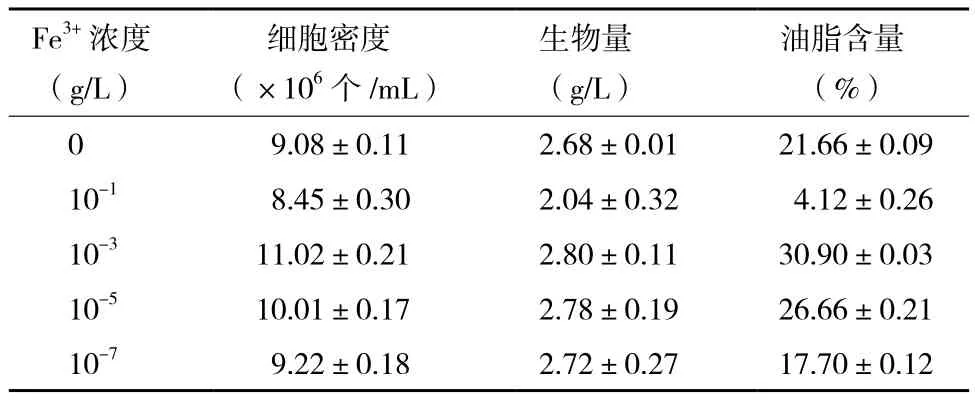
表1 不同Fe3+濃度培養條件下自養小球藻的細胞密度、生物量及油脂含量
2.1.2 Fe3+的添加時間對小球藻自養生長及產油的影響 根據上述試驗結果,選定10-3g/L作為Fe3+的添加濃度,分別在小球藻的延滯期、對數期及穩定期時添加Fe3+,對小球藻的生長及產油情況進行監測,結果(圖2,表2)顯示。延滯期添加Fe3+后的第1天,小球藻快速生長,相對于穩定期添加Fe3+小球藻的生長具有一定程度的提高(34%)。進入第2天后,小球藻的生長顯著加快,進入穩定期后其細胞密度約高于指數期及穩定期時添加Fe3+獲得細胞密度的1.5倍(圖2)。
發酵結束后,對比Fe3+不同添加時間獲得的小球藻的生物量及油脂含量可知,延滯期添加Fe3+,小球藻的生物量及油脂含量均高于其他兩個時期添加Fe3+獲得的生物量及油脂含量。
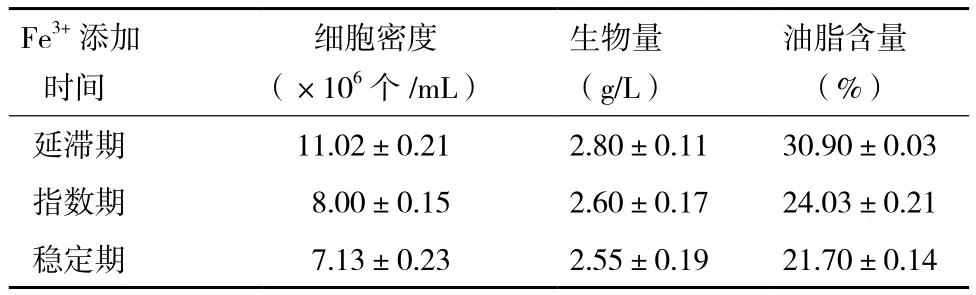
表2 不同時間添加Fe3+對自養小球藻細胞密度、生物量和油脂含量的影響
2.2 Fe3+對小球藻異養生長及產油的影響
2.2.1 Fe3+濃度對小球藻異養生長及產油的影響如圖3所示,培養基中Fe3+濃度不同,小球藻的生長差異較大。較高濃度的Fe3+不利于小球藻的生長,10-1和10-3g/L Fe3+條件下小球藻的生長狀況甚至低于不添加Fe3+時小球藻的生長。而低濃度的Fe3+(10-5和10-7g/L)有利于小球藻的生長。發酵結束后,根據小球藻的生物量及油脂含量(表3)得出,低濃度Fe3+條件下(10-5和10-7g/L)小球藻的生長大大優于高濃度Fe3+條件下(10-1和10-3g/L)小球藻的生長,前者約為后者的2.8倍。小球藻油脂含量在10-1g/L Fe3+的培養條件下最低,其他Fe3+濃度條件下小球藻的油脂含量差異不大。結合小球藻的異養生長,則10-5g/L Fe3+培養條件較優。
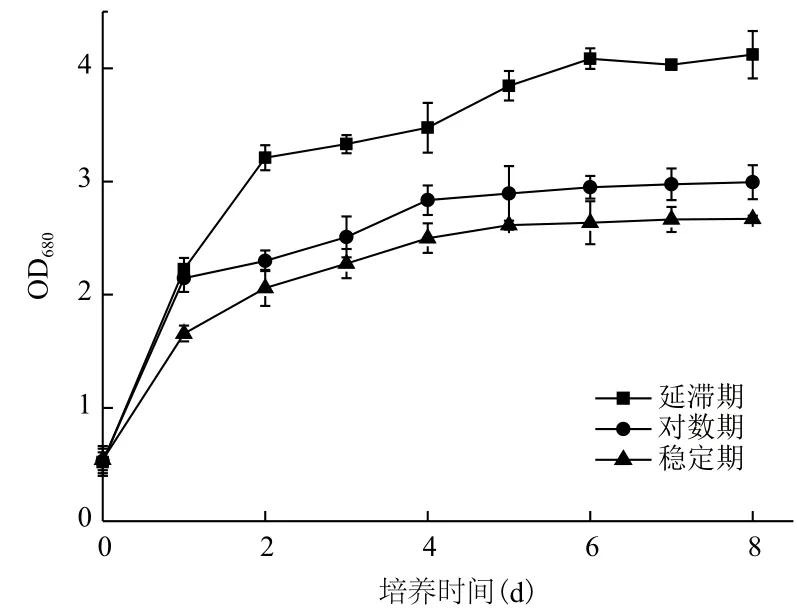
圖2 Fe3+添加時間對小球藻自養生長的影響
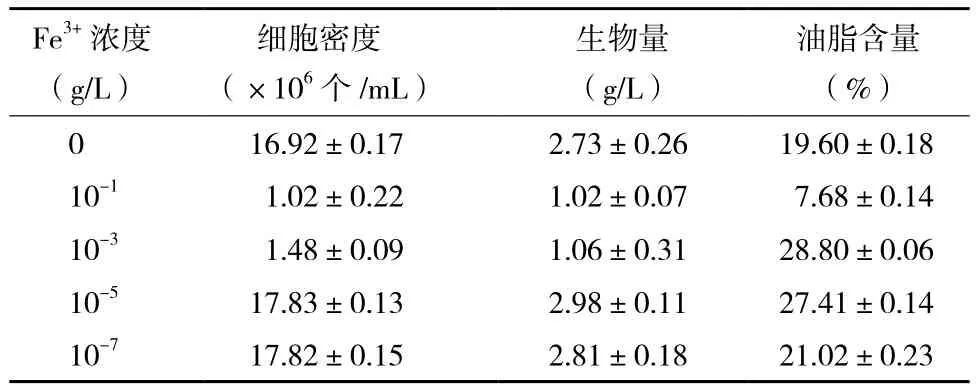
表3 不同Fe3+濃度培養條件下異養小球藻的細胞密度、生物量和油脂含量
2.2.2 Fe3+的添加時間對小球藻異養生長及產油的影響 分別對進入延滯期、對數期及穩定期的小球藻培養基中添加10-5g/L Fe3+,小球藻的生長及產油情況如圖4和表4所示。延滯期及對數期添加Fe3+,小球藻的生長約為穩定期添加Fe3+時小球藻生長的2倍。這表明,小球藻生長的前期對Fe3+的促進作用較敏感,待到小球藻生長至穩定期后,利用Fe3+來促進小球藻的生長則較為困難。對比延滯期與對
數期添加Fe3+時小球藻的油脂含量,對數期添加Fe3+獲得的小球藻的油脂含量略有優勢,達到了生物量的29.05%,約為延滯期添加Fe3+獲得的小球藻的油脂含量的1.06倍。綜合考慮小球藻的異養生長及產油,Fe3+的最佳濃度及添加時間分別為10-5g/L和指數期。
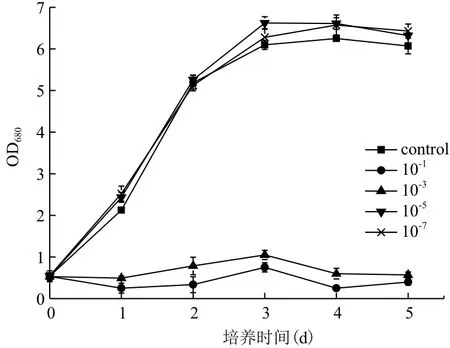
圖3 不同Fe3+濃度對小球藻異養生長的影響

表4 不同時間添加Fe3+對異養小球藻細胞密度、生物量和油脂含量的影響
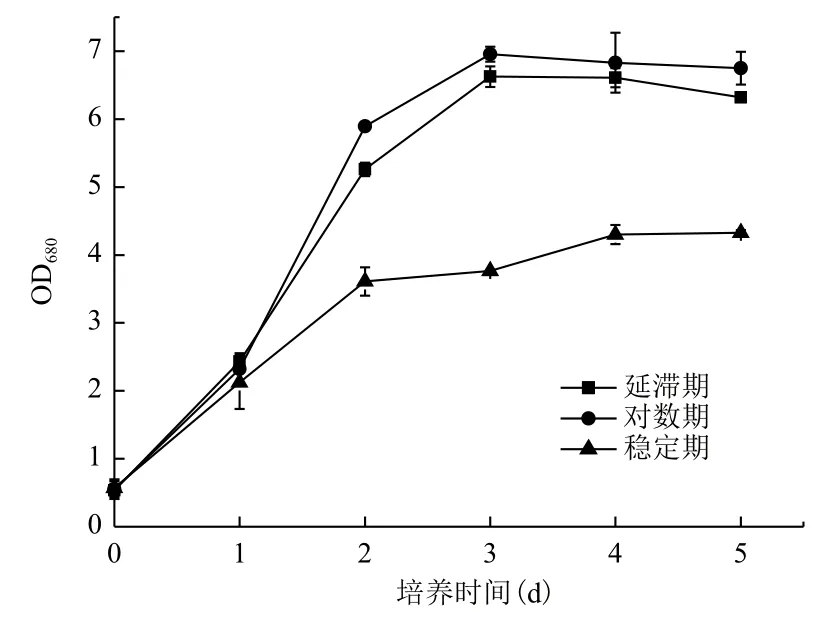
圖4 Fe3+添加時間對小球藻異養生長的影響
2.3 脂肪酸組成
分別對自養小球藻在延滯期添加10-3g/L Fe3+培養條件下獲得的油脂及異養小球藻在指數期添加10-5g/L Fe3+培養條件下獲得的油脂進行脂肪酸組成測定,結果如圖5所示。自養及異養小球藻脂肪酸組成主要為C16和C18類脂肪酸。本試驗中,自養及異養條件下C18:1占總脂肪酸的比例相當,異養條件下C16:1含量較自養條件下C16:1含量多。根據經驗公式計算[14],得到自養及異養條件下生長的小球藻生產出的油脂十六烷值分別為68.29和63.41。根據Pétursson[15]的研究,計算得到自養及異養條件下小球藻油脂的碘值分別為54.59 I2/ 100 g和71.78 I2/ 100 g。
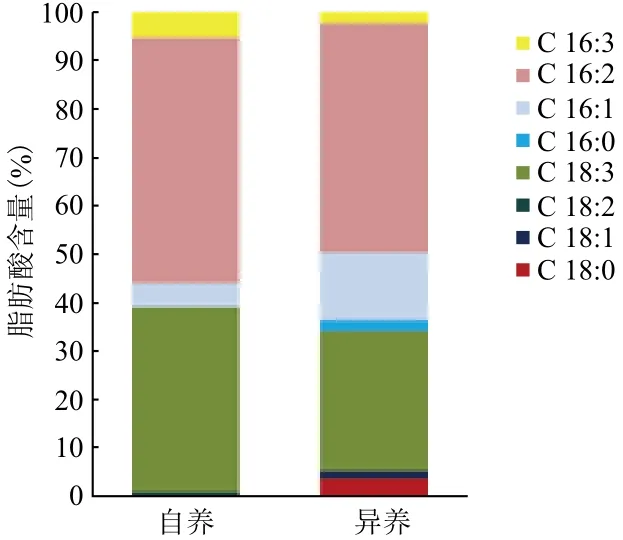
圖5 自養及異養培養條件下小球藻脂肪酸的主要成分
3 討論
小球藻自養生長時,Fe3+可以促進了與微藻生長繁殖有關的基因表達,從而提高微藻的細胞密度及生物量[16]。因此,當培養基中添加了Fe3+后,小球藻的生長優于其在不含Fe3+的培養環境下的生長。此外,Fe3+對小球藻的油脂合成途徑中某些關鍵基因的表達也具有影響,如Fe3+可以影響rbcL、accD和PAP1等基因的表達,從而誘導小球藻的油脂生成[12,17]。因此,小球藻在含有Fe3+的培養環境中生長,有利于其油脂積累。Liu等[18]以小球藻為研究對象,也發現了同樣的結果。但是,并非Fe3+在加入培養基以后,便立即對小球藻的生長發揮促進作用。而是在Fe3+加入培養基中一定時間后,其促進作用才得到充分發揮。因此,在小球藻生長的穩定期添加Fe3+,有利于Fe3+更好地發揮其對小球藻生長的促進作用。Fe3+對小球藻油脂積累的作用類似于其對小球藻生長的促進作用。在小球藻生長的穩定期向培養基中添加Fe3+,則有較為充裕的時間使Fe3+對小球藻的油脂合成途徑中關鍵基因的表達發生作用,從而生成更多的油脂。綜合以上因素,利于自養小球藻生長及產油的最佳Fe3+條件為:在小球藻生長的延滯期添加10-3g/L Fe3+。
小球藻異養生長時,Fe3+對小球藻發生促進作用具有一定的濃度限制,并非Fe3+濃度越高對小球藻生長的促進作用越好。如本實驗的結果,培養基中Fe3+濃度為10-5g/L和10-7g/L時,小球藻的生長優于高濃度Fe3+培養條件下小球藻的生長。并且,當小球藻生長至穩定期時利用Fe3+來促進小球藻的生長較為困難,可能由于此時小球藻的生長已達到一定水平,細胞的活躍程度不高,對Fe3+的敏感度不高,使得Fe3+無法較好地發揮其促進作用,因此得到小球藻的生物量及油脂產量較低。綜合考慮得到利于異養小球藻生長及產油的最佳Fe3+條件為:在小球藻生長的指數期添加10-5g/L Fe3+。
自養及異養小球藻油脂的主要成分為C16和C18類脂肪酸,此類脂肪酸是生物柴油生產利用的主要脂肪酸,與賈彬等[19]報道的結果一致。其中,十八碳三烯酸(C18:3)為細胞結構的主要構成成分[20]。C18:1和C16:1兩類脂肪酸有利于生物柴油的氧化穩定性及寒冷環境下的性能穩定性[21]。考慮生物柴油的品質,試驗獲得油脂十六烷值及碘值均符合歐洲的EN 14214標準。由此可知,實驗獲得的微藻油脂可以作為優質的原料用于生物柴油生產。
4 結論
探究了Fe3+對小球藻生長及產油的影響,獲得利于自養和異養小球藻生長及產油的最佳Fe3+濃度及添加時間分別為:延滯期添加10-3g/L Fe3+和指數期添加10-5g/L Fe3+。最優Fe3+培養條件下,獲得的自養及異養小球藻的生物量分別為2.80 g/L和3.30 g/L,油脂含量分別為30.90%和29.05%。經過對微藻油脂的脂肪酸組成分析,得出自養及異養小球藻在最優Fe3+培養條件下生產的油脂均可作為生物柴油的生產原料。
[1] Aubert S, Gout E, Bligny R, et al. Multiple effects of glycerol on plant cell metabolism. Phosphorus-31 nuclear magnetic resonance studies[J]. Int J Biol Chem, 1994, 269(34):21420-21427.
[2] Lin J, Shen H, Tan H, et al. Lipid production by Lipomyces starkeyi cells in glucose solution without auxiliary nutrients[J]. J Biotechnol, 2011, 152(4):184-188.
[3] 宋東輝, 侯李君, 施定基. 生物柴油原料資源高油脂微藻的開發利用[J]. 生物工程學報, 2008, 24(3):341-348.
[4] Chisti Y. Biodiesel from microalgae[J]. Biotechnol Adv, 2007, 25(3):294-306.
[5] Xie T, Sun Y, Du K, et al. Optimization of heterotrophic cultivation of Chlorella sp. for oil production[J]. Bioresour Technol, 2012, 118:235-242.
[6] McPhee WA, Boucher M, Stuart J, et al. Demonstration of a liquid-tin anode solid-oxide fuel cell(LTA-SOFC)operating from biodiesel fuel[J]. Energy Fuels, 2009, 23(10):5036-5041.
[7] 繆曉玲, 吳慶余. 微藻生物質可再生能源的開發利用[J]. 可再生能源, 2003, 3(109):13-16.
[8] Qiao H, Wang G. Effect of carbon source on growth and lipid accumulation in Chlorella sorokiniana GXNN01[J]. Chin J Oceanol Limnol, 2009, 27:762-768.
[9] Hoque ME, Singh A, Chuan YL. Biodiesel from low cost feedstocks:The effects of process parameters on the biodiesel yield[J]. Biomass Bioenergy, 2011, 35(4):1582-1587.
[10] Spolaore P, Joannis-Cassan C, Duran E, et al. Commercial applications of microalgae[J]. J Biosci Bioeng, 2006, 101(2):87-96.
[11] 蔣漢明, 高坤山. 氮源及其濃度對三角褐指藻生長和脂肪酸組成的影響[J]. 水生生物學報, 2004, 28(5):545-551.
[12] Behrenfeld MJ, Worthington K, Sherrell RM, et al. Controls on tropical Pacific Ocean productivity revealed through nutrient stress diagnostics[J]. Nature, 2006, 442(7106):1025-1028.
[13] Wang W, Zhou W, Liu J, et al. Biodiesel production from hydrolysate of Cyperus esculentus waste by Chlorella vulgaris[J]. Bioresour Technol, 2013, 136:24-29.
[14] 陳林, 王忠, 李銘迪, 等. 生物柴油組分與十六烷值關系研究[J]. 中國油脂, 2012, 37(1):53-56.
[15] Pétursson S. Clarification and expansion of formulas in AOCS recommended practice Cd 1c-85 for the calculation of lodine value from FA composition[J]. J Am Oil Chem Soc, 2002, 79(7):737-738.
[16] 陳莉, 梁文艷, 王金麗, 等. 鐵對銅綠微囊藻生長活性及產毒的影響[J]. 安全與環境學報, 2009, 9(4):21-24.
[17] Michel KP, Pistorius EK. Adaptation of the photosynthetic electron transport chain in cyanobacteria to iron deficiency:the function of IdiA and IsiA[J]. Physiol Plant, 2004, 120(1):36-50.
[18] Liu Z, Wang G, Zhou B. Effect of iron on growth and lipid accumulation in Chlorella vulgaris[J]. Bioresour Technol, 2008, 99(11):4717-4722.
[19] 賈彬, 王亞南, 何蔚紅, 等. 生物柴油新原料——微生物油脂[J]. 生物技術通報, 2014(1):19-26.
[20] Garcí MC, Sevilla JF, Fernández FA, et al. Mixotrophic growth of Phaeodactylum tricornutum on glycerol:growth rate and fatty acid profile[J]. J Appl Phycol, 2000, 12(3-5):239-248.
[21] Pinzi S, Garcia I, Lopez-Gimenez F, et al. The ideal vegetable oilbased biodiesel composition:a review of social, economical and technical implications[J]. Energy Fuels, 2009, 23(5):2325-2341.
(責任編輯 李楠)
Effects of Fe3+on the Growth and Oil Content of Chlorella vulgaris for Biodiesel Production
Sun Yuan Liu Wenbin Zhou Tiezhu Xie Tonghui Liang Bin Zhang Yongkui
(Department of Pharmaceutical & Biological Engineering,School of Chemical Engineering,Sichuan University,Chengdu 610065)
Biodiesel is a clean renewable energy as the high quality substitute for traditional diesel. Chlorella vulgaris, used for biodiesel production, has been gaining the attention of the society because of its fast growth and high oil content. Iron ion could improve the growth and oil content of C. vulgaris. Effects of the concentration of Fe3+and the time to add Fe3+on the growth and oil content of autotrophic and heterotrophic C. vulgaris were explored in this study. The best conditions of Fe3+were as follow:(1)10-3g/L Fe3+added in the lag phase of autotrophic C. vulgaris, biomass of 2.80 g/L and oil content of 30.90% were obtained, (2)10-5g/L Fe3+added in the exponential phase of heterotrophic C. vulgaris, biomass of 3.30 g/L and oil content of 29.05% were obtained. And the oil produced by C. vulgaris under autotrophic and heterotrophic conditions was an ideal feedstock for biodiesel production.
Chlorella vulgaris Fe3+Autotrophy Heterotrophy Growth Oil content
2014-02-10
四川大學青年教師科研啟動基金項目(2014SC011058)
孫遠,女,碩士研究生,碩士,研究方向:微生物發酵;E-mail:suny88@163.com
張永奎,博士,教授,博士生導師,研究方向:應用微生物;E-mail:zhangyongkui@scu.edu.cn
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
中國糧油學報(2018年12期)2018-01-26 13:32:04
