抗蟲轉基因銀河楊的生長特性和抗蟲性
2014-03-21 11:47:16陳虞超張麗鞏檑甘曉燕石磊宋玉霞
生物技術通報 2014年11期
關鍵詞:生長
陳虞超 張麗 鞏檑 甘曉燕 石磊 宋玉霞
(寧夏農業生物技術重點實驗室,銀川 750002)
抗蟲轉基因銀河楊的生長特性和抗蟲性
陳虞超 張麗 鞏檑 甘曉燕 石磊 宋玉霞
(寧夏農業生物技術重點實驗室,銀川 750002)
以非轉基因銀河楊(Populus alba×Populus hopeiensis)為對照,對抗蟲轉基因銀河楊抗蟲基因Cry3A的穩定性,轉基因銀河楊的物候期、生長狀況和抗蟲性進行研究。結果發現,Cry3A基因整合穩定,在根、莖、葉中RNA和蛋白水平均有表達,不同組織中Bt蛋白水平差異顯著,莖中Bt蛋白水平最高。抗蟲轉基因銀河楊的物候期、生長狀況未發生顯著變化,但表現出顯著的抗蟲性。
抗蟲轉基因銀河楊 Cry3A基因 物候期 生長狀況 抗蟲性
楊樹(Populus L.)生長快,適應性強,用途廣,是我國人工速生用材基地建設的主要造林樹種,影響楊樹豐產林發展最大的障礙為病蟲危害,其中蟲害尤甚。楊樹的害蟲主要分為食葉害蟲和蛀干害蟲,食葉害蟲以鱗翅目(Lepidoptera)的楊尺蠖(Apocheima cinerarius)和舞毒蛾(Lymantria dispar)、楊扇舟蛾(Clostera anachoreta)為主,蛀干害蟲以鞘翅目(Coleoptera)的光肩星天牛(Anoplophora glabripennis)、桑天牛(Apriona germari)、云斑天牛(Batocera horsfieldi)為主[1,2]。防治這些害蟲的傳統方法如化學藥物防治和人工捕捉等具有一定局限性,因此從20世紀80年代開始,我國開展了抗蟲轉基因楊樹培育的研究。眾多研究者通過轉基因技術將蘇云金芽孢桿菌(Bacillus thuringiensis,簡稱Bt)等外源抗蟲基因,導入不同楊樹品種中,成功培育出一系列性狀優良的抗蟲轉基因楊樹,其中部分已進入環境釋放或商品化階段[3-8]。Cry3A為Bt蛋白編碼基因之一,對鞘翅目昆蟲具有專一的毒殺作用,在抗蟲轉基因楊樹的培育中應用較多[9-11]。目前有關抗蟲轉基因楊樹培育和利用的研究較多,針對抗蟲轉基因楊樹生長特性和抗蟲性變化的跟蹤研究較少。鑒于此,本研究以普通銀河楊為對照,對已育成的轉Cry3A抗蟲基因銀河楊抗蟲基因的穩定性,物候期、生長狀況和抗蟲性變化進行較系統
的對比研究,旨在為抗蟲基因銀河楊下一步推廣應用奠定基礎。
1 材料與方法
1.1 材料
轉Cry3A抗蟲基因銀河楊[銀白楊(Populus alba L.)×河北楊(Populus hopeiensis Hu et chow)]以CaMV35S為Cry3A基因的啟動子,由寧夏農業生物技術重點實驗室培育,并于2004年種植于寧夏回族自治區銀川市寧夏農林科學院蘆花臺園林場(東經10608'、北緯3838')。測試昆蟲為人工飼養的鞘翅目(Coleoptera)天牛科(Cerambycidae)的光肩星天牛(Anoplophora glabripennis)。DNA提取試劑盒購自TIANGEN公司(貨號:DP305-02),RNA提取試劑盒購自全式金公司(貨號:ET121-01),RTPCR試劑盒GoScriptTMReverse Transeription System購自Promega公司,Bt-Cry3A的ELISA測定試劑盒購自于Agdia公司(貨號:PSA05900),其他試劑為國產分析純。
1.2 方法
1.2.1 抗蟲轉基因銀河楊Cry3A基因的PCR與RT-PCR檢測 提取抗蟲轉基因銀河楊DNA,利用PCR方法檢測Cry3A基因的穩定性,引物序列為:5'-ATGACTGCTGATAACAACACGGA-3'和 5'-TTAATTCACTGGAATGAACTCA-3',擴增片段大小為1 794 bp。PCR體系為10×buffer 2.5 μL,dNTP 2.0 μL,引物各1.0 μL,Taq 1.0 μL,DNA 1.5 μL,ddH2O 17 μL,總體積25 μL。擴增程序為:94℃預變性5 min;94℃變性40 s,53℃復性40 s,72℃延伸1 min,循環35次;72℃保溫10 min。7月中旬,采集轉基因銀河楊根、莖、葉,采用試劑盒法提取抗蟲轉基因銀河楊的總RNA,以楊樹的ubiquitin基因(Gene ID:7464783)為內標,利用RT-PCR檢測抗蟲基因Cry3A的反轉錄情況;Cry3A的RTPCR引物同PCR引物一致,內標基因ubiquitin的引物序列為:5'-TGAGGCTTAGGGGAGGAACT-3'和5'-TGTAGTCGCGAGCTGTCTTG-3',擴增片段大小為195 bp;擴增程序參照試劑盒說明書。
1.2.2 抗蟲轉基因銀河楊Bt蛋白的檢測 于7月中旬,采集抗蟲轉基因銀河楊根、莖、葉組織,然后測定根、莖、葉中Bt蛋白的含量,具體過程參照試劑盒說明書進行,用Bio-Rad xMarkTM酶標儀測定結果。
1.2.3 抗蟲轉基因銀河楊生長特性調查 采用隨機抽樣法,選取抗蟲轉基因銀河楊(簡稱TG)和普通銀河楊(簡稱CK)調查樣株各30株,從2012-2013年,連續2年對所有樣株的生長物候期(包括芽萌動期、芽開放期、花序孕期、展葉始期、開花期、展葉盛期、封頂期、葉始黃期、葉黃盛期、落葉初期、落葉盛期)和形態特性(包括株高、胸徑、地徑、分支數、郁閉度)進行觀測。
1.2.4 抗蟲轉基因銀河楊室內抗蟲性鑒定 于7月中旬,分別采集抗蟲轉基因銀河楊、普通銀河楊植株的幼嫩枝葉,以容量為1 L的帶透氣蓋塑料培養瓶為飼養容器,飼喂光肩星天牛成蟲,每瓶3頭,每個處理組10瓶,每2 d換一次新鮮枝葉,重復3次,統計兩個試驗組光肩星天牛成蟲15 d內總死亡率和死亡指數。
總死亡率(%)=(飼養末期死亡總數/初期飼養總數)×100%
死亡指數=(∑天數×當天死亡數)/(總死亡數×試驗總天數)
2 結果
2.1 抗蟲轉基因銀河楊Cry3A基因的PCR檢測
通過PCR方法檢測Cry3A基因在抗蟲轉基因銀河楊植株中的穩定性,結果如圖1所示,抗蟲轉基因銀河楊植株的PCR擴增產物中,均存在一條1 800 bp左右大小的條帶,經測序比對與目的基因Cry3A同源性為100%。結果表明,Cry3A基因在抗蟲轉基因銀河楊植株中穩定存在,未發生基因丟失的現象。

圖1 抗蟲轉基因銀河楊Cry3A基因的PCR擴增產物電泳圖
2.2 抗蟲轉基因銀河楊Cry3A基因的RT-PCR檢測
通過RT-PCR方法檢測Cry3A基因在不同組織中的RNA表達情況,結果如圖2所示,在抗蟲轉基因銀河楊植株根、莖、葉的RT-PCR擴增產物中,均存在一條1 800 bp左右大小的條帶,經測序比對與目的Cry3A基因同源性為100%。結果表明,Cry3A基因在根、莖、葉中均能表達。

圖2 抗蟲轉基因銀河楊Cry3A基因RT-PCR擴增產物電泳圖
2.3 抗蟲轉基因銀河楊Bt蛋白檢測
通過ELISA法檢測Cry3A基因在不同組織中的Bt蛋白表達情況,結果如圖3所示,抗蟲轉基因銀河楊植株根、莖、葉中Bt蛋白水平分別為16、28和6 ng/g。結果表明,Cry3A基因在根、莖、葉中均能進行Bt蛋白表達,莖中水平最高;不同組織中Bt蛋白水平存在顯著差異。
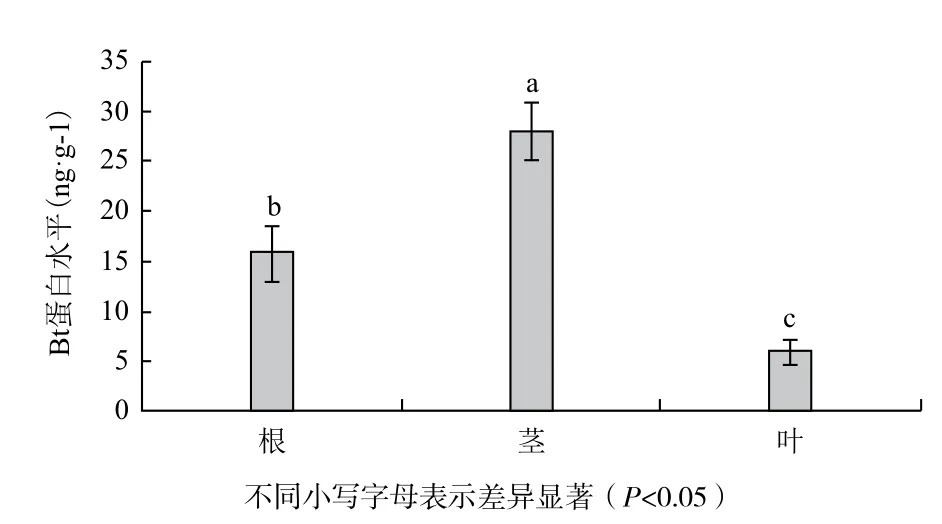
圖3 抗蟲轉基因銀河楊不同組織中Bt蛋白水平
2.4 抗蟲轉基因銀河楊生物學特性分析
通過對抗蟲轉基因銀河楊、普通銀河楊的物候期進行對比調查,結果發現二者的物候期無顯著差異(表1)。通過對抗蟲轉基因銀河楊、普通銀河楊的生長狀況進行對比調查,結果發現二者在胸徑年均生長量、地徑年均生長量、株高年均生長量、分支數、郁閉度方面均無顯著差異(表2)。

表1 轉基因銀河楊與普通銀河楊物候期對比表

表2 轉基因銀河楊與普通銀河楊生長狀況對比表
2.5 轉基因銀河楊抗蟲性分析
以普通銀河楊為對照,通過實驗室人工飼喂的方法,對比分析抗蟲轉基因銀河楊的抗蟲性,結果如表3所示,飼喂抗蟲轉基因銀河楊和普通銀河楊的光肩星天牛,總死亡率分別為86.7%、33.3%,死亡指數分別為0.62、0.24。表明抗蟲轉基因銀河楊對光肩星天牛具有顯著的抗性。

表3 轉基因銀河楊與普通銀河楊抗蟲性的對比分析
3 討論
外源基因在轉基因植株中丟失、沉默是植物基因工程應用中存在的主要問題。Meyer等[12]發現,轉基因移栽至大田后,由于光照、溫度等因素的影響,外源基因的失活程度更顯著,失活的轉基因植物數量也更多。但胡建軍等[13]通過田間調查發現,7年生的轉基因黑楊依然具有良好的抗蟲性,外源抗蟲基因表達正常。本研究對9年生的抗蟲轉基因銀河楊部分植株進行PCR檢測,發現Cry3A基因在轉基因楊樹植株中穩定存在,未發生基因丟失的現象。抗蟲轉基因銀河楊以CaMV35S作為Cry3A基因的啟動子,而CaMV35S屬異源組成型啟動子,在受體植株的不同組織器官中均可表達[14,15]。本研究發現,Cry3A基因在轉基因銀河楊植株的根、莖、葉均可表達,這可能是CaMV35S啟動子的特性所決定的。不同組織間的表達量差異顯著,這與姜志磊等[16]對抗蟲轉基因玉米的研究結論相似,推測這一現象可能與Bt蛋白的分泌方式、運輸途徑和方向等相關。
外源抗蟲基因導入受體植株基因組,使受體植株表現出較好的抗蟲性,但可能也會影響植株的新陳代謝,導致其生長特性等表型發生變化。呂淑平等[17]研究表明,外源Bt基因在增強轉基因棉花抗性的同時,還影響了轉基因棉花的株高等生長特性。但本研究發現,與普通銀河楊相比,抗蟲轉基因銀河楊在物候期、生長狀況等生長特性方面不存在顯著變化,這與劉海濤等[18]對抗蟲轉基因歐洲黑楊、楊敏生等[19]對轉雙抗蟲基因741楊的研究結果一致。本研究還發現,與普通銀河楊相比,抗蟲轉基因銀河楊對光肩星天牛具有顯著的抗性,這與張冰玉等[11]對轉Cry3A基因銀腺楊的相關研究結論一致。
4 結論
抗蟲轉基因銀河楊(Populus alba×Populus hopeiensis)的物候期、生長狀況未發生顯著變化,但抗蟲性顯著;其抗蟲基因Cry3A整合穩定,在根、莖、葉中RNA和蛋白水平均有表達,不同組織中Bt蛋白水平差異顯著,莖中Bt蛋白水平最高。
[1]王永俊. 我國楊樹害蟲防治研究概述[J]. 江蘇林業科技, 1991(4):47-48, 26.
[2]皮兵, 肖興翠, 曹建紅, 等. 楊樹抗蟲轉基因研究進展[J]. 湖南林業科技, 2012, 39(4):66-71.
[3]伍寧豐, 范云六. 含蘇云金芽孢桿菌蛋白基因的楊樹工程植株的建立[J]. 科學通報, 1991(9):705-708.
[4]田穎川, 李太元, 莽克強, 等. 抗蟲轉基因歐洲黑楊的培育[J].生物工程學報, 1993, 9(4):291-297.
[5]王學聘, 韓一凡, 戴蓮韻, 等. 抗蟲轉基因歐美黑楊的培育[J].林業科學, 1997, 33(1):69-74.
[6]張雁, 郭同斌, 等. 轉Bt基因南林895楊抗蟲性及對土壤微生物影響分析[J]. 林業科學研究, 2012, 25(3):346-350.
[7]胡建軍, 楊敏生, 盧孟柱. 我國抗蟲轉基因楊樹生態安全性研究進展[J]. 生物多樣性, 2010, 18(4):336-345.
[8]蘇曉華, 張冰玉, 黃秦軍, 等. 我國林業基因工程研究進展及關鍵領域[J]. 林業科學, 2003, 39(5):111-118.
[9]甄志先, 李靜, 梁海永, 等. 轉BtCry3A基因楊樹毒蛋白表達及對桑天牛抗性的研究[J]. 蠶業科學, 2007, 33(4):538-542.
[10]王彥平, 李靜, 楊敏生, 等. 轉不同抗蟲基因741楊的抗蟲選擇性[J]. 林業科學, 2008, 44(8):67-71.
[11]張冰玉, 蘇曉華, 等. 轉抗鞘翅目害蟲銀腺楊的獲得及其抗蟲性的初步研究[J]. 北京林業大學學報, 2006, 28(2):102-105.
[12]Meyer P, Heidmann I. Epigenetic variants of a transgenic petunia line show hypermehhylation in transgene DNA:an indication for specific recognition of foreign DNA in transgenic plant[J]. Mol Gen Genet, 1994(243):390-399.
[13]胡建軍, 李淑梅, 盧孟柱, 等. 轉Bt基因歐洲黑楊抗蟲穩定性及其對天敵昆蟲的影響[J]. 林業科學研究, 2007, 20(5):656-659.
[14]張春曉, 王文棋, 蔣湘寧, 等. 植物基因啟動子研究進展[J].遺傳學報, 2004, 31(12):1455-1464.
[15]路靜, 趙華燕, 何奕昆, 等. 高等植物啟動子及其應用研究進展[J]. 自然科學進展, 2004, 14(8):856-862.
[16]姜志磊, 劉德璞, 李曉輝, 等. 轉基因抗蟲玉米Bt毒蛋白時空表達分析[J]. 吉林農業科學, 2008, 33(6):35-37.
[17]呂淑平, 郭小平, 等. 轉基因抗蟲棉Bt基因導入對受體材料農藝性狀的影響[J]. 農業生物技術科學, 2004, 20(3):36-37.
[18]劉海濤, 張川紅, 鄭勇奇, 等. 抗蟲轉基因歐洲黑楊苗期光合特征研究[J]. 北京林業大學學報, 2011, 33(1):36-43.
[19]楊敏生, 高寶嘉, 王進茂, 等. 轉雙抗蟲基因741楊基本特性分析[J]. 林業科學, 2005, 41(1):91-97.
(責任編輯 李楠)
Growth Characteristic and Insect-Resistance of Transgenic Populus alba Populus hopeiensis
Chen Yuchao Zhang Li Gong Lei Gan Xiaoyan Shi Lei Song Yuxia
(Key Laboratory of Agricultural Biotechnology of Ningxia,Yinchuan 750002)
The stability of insect-resistance gene Cry3A, phenophase, growth status and insect-resistance of transgenic poplar hybrid(Populus alba×Populus hopeiensis)was investigated, non- transgenic Populus alba×Populus hopeiensis as control. The results showed that the Cry3A gene in transgenic Populus alba×Populus hopeiensis integrated stably. The Cry3A gene could be expressed at RNA level and protein level in root, shoot and leaf. The Bt protein level was significantly different in different tissues, and was highest in shoot. The phenophase and growth status of transgenic Populus alba×Populus hopeiensis changed indistinctively. But it showed distinct insect-resistance.
Insect-resistance transgenic Populus alba×Populus hopeiensis Cry3A gene Phenophase Growth status Insectresistance
2014-03-07
國家林業轉基因生物安全性監測項目(JJ-2012-07)
陳虞超,男,碩士,助理研究員,研究方向:植物基因工程;E-mail:chenyuchao820321@163.com
宋玉霞,女,碩士,研究員,研究方向:植物分子生物學;E-mail:songyx666@163.com
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
