戊糖乳桿菌發酵產乳酸及高產菌株誘變選育
2014-03-21 11:47:24王義強王啟業馬國輝林麗云
生物技術通報 2014年11期
王義強王啟業馬國輝林麗云
(1.中南林業科技大學生物技術實驗室,長沙 410004;2.經濟林培育與保護教育部重點實驗室,長沙 410004;3.國家林業局生物乙醇研究中心,長沙 410004)
戊糖乳桿菌發酵產乳酸及高產菌株誘變選育
王義強1,2,3王啟業1馬國輝1林麗云1
(1.中南林業科技大學生物技術實驗室,長沙 410004;2.經濟林培育與保護教育部重點實驗室,長沙 410004;3.國家林業局生物乙醇研究中心,長沙 410004)
戊糖乳桿菌(Lactobacillus pentosus)是能利用木質纖維素水解液發酵產乳酸的潛力菌株,發酵條件優化與高產菌株的選育是提高乳酸產量的重要手段。通過單因素試驗、Plackett-Burman設計與響應面試驗,對戊糖乳桿菌ATCC 8041產乳酸的發酵培養基及發酵條件進行了優化。結果表明,該菌株發酵培養基的最佳組合為葡萄糖93.11 g/L、酵母浸粉5.19 g/L、碳酸鈣29.43 g/L、蛋白胨10.00 g/L、Na2HPO4·12H2O 5.00 g/L、MgSO40.20 g/L、MnSO450 mg/L;最佳發酵條件為37℃、pH6.5、接種量6%、裝液量80%。在此優化條件下,該菌株發酵產乳酸為54.12 g/L。進一步以戊糖乳桿菌ATCC 8041為出發菌株,通過原生質體進行紫外誘變,經多重篩選,最終獲得一株遺傳穩定性好的高產乳酸突變株,命名為戊糖乳桿菌Lactic UVC-02,由中國典型培養物保藏中心保存,注冊號為CCTCC M 2013209。該突變株Lactic UVC-02經葡萄糖發酵,乳酸產量達64.17 g/L,比出發菌株ATCC 8041(54.12 g/L)提高18.6%。
戊糖乳桿菌 乳酸發酵 紫外誘變 響應面試驗
乳酸(Lactic acid),又名丙醇酸,是一種在人體、動植物、微生物體內普遍存在的有機酸[1]。乳酸及其衍生物可廣泛應用于食品、化工、紡織、制革、電子、醫藥以及農業等諸多領域[2,3]。隨著環境污染問題
日益嚴重,應用新技術開發新型塑料,從而解決傳統塑料造成的“白色污染”問題,已成為各國研究的熱點[4]。乳酸可聚合生產聚乳酸來制造生物降解塑料、綠色包裝材料及農業薄膜。聚乳酸無毒,無刺激性,具有較好的生物相容性和生物降解性。以聚乳酸來加工成塑料可解決日益嚴重的環境污染問題,引起世界的廣泛關注。乳酸的生產方法有酶合成法、化學合成法、微生物發酵法3種[5]。工業化生產多采用前兩種方法,但它們目前面臨原料來源有限、成本急劇增加的困境,最主要的是生產過程中帶來嚴重的環境污染。因此,通過微生物發酵法生產乳酸將成為未來發展的主要趨勢。
目前,能應用到工業生產乳酸的微生物只有霉菌中的根霉屬和細菌中的乳酸菌類[6-8]。利用根霉屬的米根霉生產乳酸在國內外研究很多,但其發酵產乳酸存在糖轉化率低(理論值75%)、有氧混合發酵成本高和副產物多等不足[5,9]。乳酸細菌能利用葡萄糖(或可利用的碳水化合物)發酵產乳酸,糖轉化率可達90%以上,且發酵成本低,產物主要為L-型乳酸和D-乳酸混合物[10]。部分乳酸菌甚至可利用木質纖維素的水解產物,如木糖、阿拉伯糖等五碳糖,發酵產生乳酸[11-13]。
傳統的乳酸發酵常以糧食作物為底物,不僅浪費糧食,且使乳酸價格高昂。因此,尋找和選育能夠利用木質纖維素水解液為原料的高產乳酸菌株迫在眉睫。因傳統誘變具有誘變效率高且不易回復突變等特性,工業生產上乳酸菌種的選育多數是以傳統的誘變選育為主[14]。戊糖乳桿菌(Lactobacillus pentosus)ATCC 8041不僅能利用六碳糖也能利用五碳糖發酵生產乳酸,是能利用木質纖維素水解液發酵產乳酸的潛力菌株[15]。本研究首次采用紫外線對出發菌株戊糖乳桿菌ATCC 8041原生質體進行誘變處理,期望篩選出高產乳酸突變株。
1 材料與方法
1.1 材料
1.1.1 菌種 本試驗所用的戊糖乳桿菌(Lactobacillus pentosus)ATCC 8041購于美國典型培養物保藏中心。
1.1.2 培養基 MRS肉湯活化培養基:眎蛋白胨10 g/L,酵母提取物5 g/L,牛肉膏10 g/L,葡萄糖20 g/L,山梨醇酐單油酸1 g/L,檸檬酸銨2 g/L,MnSO4·H2O 0.05 g/L,CH3COONa·3H2O 7.73 g/L,Na2HPO4·12H2O 5.04 g/L,H2O 1 000 mL,固體培養基添加15 g/L的瓊脂,pH6.5。
發酵培養基:葡萄糖90 g/L,眎蛋白胨10 g/L,酵母粉5 g/L,Na2HPO4·12H2O 5 g/L,MgSO40.2 g/L,MnSO4·H2O 0.05 g/L,CaCO330 g/L。
種子培養基(改良的MRS培養基):眎蛋白胨10 g/L、酵母提取物5 g/L、牛肉膏10 g/L、葡萄糖20 g/L、山梨醇酐單油酸1 g/L、檸檬酸銨2 g/L、MnS04·H2O 0.05 g/L、三水合乙酸鈉7.73 g/L、十二水合磷酸氫二鈉5.04 g/L,pH6.5。
斜面培養基:在種子培養基的基礎上,添加瓊脂20 g/L。
篩選培養基:改良的MRS培養基中加入0.01%溴鉀酚綠。
高滲再生培養基:采用雙層瓊脂法,用0.7 mol/L NaCl配制MRS培養基,上層含2%瓊脂,下層含3%的瓊脂。
滲透壓穩定劑:0.7 mol/L NaCl溶液。
1.2 方法
1.2.1 種子培養 取1 mL MRS肉湯培養基于安瓿管中溶解菌體,稀釋后涂布平板,培養3 d;取平板培養菌落接種于含10 mL種子培養基的25 mL試管,37℃靜置培養10 h為一級種子液。取1 mL一級種子液接種于含100 mL種子培養基的250 mL錐形瓶,37℃恒溫搖床中150 r/min培養10 h后用于試驗。
1.2.2 發酵培養基單因素試驗 以乳酸產量為考察指標,對發酵培養基中的7個組分:葡萄糖(60、75、90、105、120和135 g/L)、蛋白胨(0、5、10、15和20 g/L)、酵母粉(0、2.5、5、7.5和10 g/L)、Na2HPO4·12H2O(0、2.5、5、7.5和10 g/L)、MgSO4(0、0.1、0.2、0.3、0.4和 0.5 g/L)、MnSO4·H2O(0、25、50、75和100 mg/L)、CaCO3(0、10、20、30、40、50和60 g/L)進行單因素試驗,除目標因素外,其余培養基組分均按發酵培養基和已篩選因素中的較佳條件進行試驗,每組3個平行,37℃厭氧發酵72 h,液相色譜檢測產物,結果取平均值。
1.2.3 Plackett-Burman試驗設計與響應面試驗 在
單因素試驗的基礎上確定低水平(-1)值,高水平(1)為低水平的1.25倍,選取影響乳酸產量較大的7個因素(葡萄糖、蛋白胨、酵母粉、Na2HPO4、MgSO4、MnSO4、CaCO3)進行全面考察,選用N=12的PB設計,以乳酸產量(Y)為響應值,利用試驗設計和數據分析軟件Minitab15進行Plackett-Burman設計[16,17],根據軟件給出的試驗矩陣表確定各試驗組的編碼與取值進行試驗,再根據試驗結果進行數據分析,篩選出重要影響因素。
根據Plackett-Burman設計試驗結果,采用Box-Behnken設計進行試驗[18,19],所得結果利用軟件以乳酸產量(Y)為響應值做響應面分析,從而獲得培養基中重要因子的最佳組合。
1.2.4 發酵條件單因素試驗 在發酵培養基優化的基礎上,以乳酸產量為考察指標,分別對溫度(31、33、35、37、39、41和43℃)、初始pH(4.0、4.5、5.0、5.5、6、6.5和7.0)、種子液接種量(2%、4%、6%、8%、10%和12%)及100 mL發酵瓶裝液量(65%、70%、75%、80%、85%、90%和95%) 依次進行單因素試驗。除目標因素外,其余培養條件按已篩選的最佳條件進行試驗,每組3個平行,進行厭氧發酵,液相色譜檢測,結果取平均值。
1.2.5 原生質體制備 取10 mL液體種子培養物于滅過菌的離心管中,4 000 r/min 離心15 min,去上清液,用無菌生理鹽水洗滌收集的菌體兩次,加入3 mL含溶菌酶的滲透壓穩定劑中,將制備的菌酶混合液在30℃的條件下酶解2 h,將酶解液3 000 r/min離心15 min,取上清液收集原生質體,將原生質體懸浮于10 mL滲透壓穩定劑中備用。
1.2.6 原生質體紫外誘變 取5 mL經高滲溶液稀釋后原生質體溶液,混勻,倒入直徑9 mm的培養皿中,在磁力攪拌下距功率20 W的紫外燈35 cm處進行0、20、40、60、80、100和120 s紫外照射。誘變結束后,取不同時間的處理液100 μL涂布在高滲再生培養基上,每個處理3個重復,37℃條件下恒溫避光培養72 h。以時間為橫坐標,致死率為縱坐標,繪制致死曲線。致死率(%)=(對照1 mL菌液中活菌數-處理后1 mL菌液中活菌數)/對照1 mL菌液中活菌數×100%。
1.2.7 高產突變株的篩選 將最佳處理原生質體稀釋涂布于原生質體再生培養基上,72 h后用無菌牙簽將菌落轉接在篩選培養基平板上。2 d后培養基為綠色,菌株的代謝產物乳酸會使菌落周圍pH降低,形成黃色顯色圈,挑取顯色圈較大的90株菌株進行搖瓶發酵,每個菌株接1瓶,37℃培養,72 h后測其乳酸產量。得到乳酸產量較高的菌株再次進行發酵復篩,每株3瓶,37℃培養,72 h后測其乳酸產量。
1.2.8 高產突變株遺傳穩定性試驗 對篩選獲得的高活性的突變株進行繼代培養,連續繼代8次,每代5個平行,測其乳酸產量,分析其穩定性。
1.2.9 乳酸的液相色譜法檢測
1.2.9.1 采用液相色譜[20]測定發酵液中乳酸的含量 色譜條件:色譜儀為安捷倫1100[含在線脫氣機,可變波長檢測器(VWD),agilent 1100四元泵],色譜柱為EDipse XDB-C18(150 mm×4.6 mm 1.d. 5 μm),流動相為0.005 mol/L H2SO4溶液,pH2.5(用NaOH調節),流速1.0 mL/min,檢測波長210 nm,進樣量20 μm,柱溫為室溫。
1.2.9.2 流動相配制 0.27 mL分析純濃硫酸用水稀釋并定容至1 000 mL(0.005 mol/L),用NaOH溶液調節pH值至2.5,然后用0.45 μm孔徑的合成纖維素酯膜過濾。
1.2.9.3 對照品制備及標準曲線繪制 精密稱取乳酸1 g標準品放入10 mL容量瓶中,用超純水溶解并定容至10 mL,配成乳酸標準液,然后進行稀釋,分別配成0.5、1、2、4和5 g/L的標準溶液。用0.45 μm孔徑的合成纖維素酯膜過濾。再分別經液相色譜檢測,以濃度為橫坐標,峰高值為縱坐標,繪制出的標準曲線如圖1所示。

圖1 乳酸標準曲線
1.2.9.4 樣品的制備 取5 mL發酵液(5 000 r/min,10 min)除去碳酸鈣,加5 mL 0.01 mol/L的硫酸溶
液進行酸解10 min,離心除去硫酸鈣,以0.45 μm的濾膜過濾,取1 mL濾液適量稀釋即得待測液。
2 結果
2.1 戊糖乳桿菌ATCC 8041最佳發酵培養基確定
發酵培養基中的7個因素的單因素試驗結果為葡萄糖90 g/L、蛋白胨10 g/L、酵母粉5 g/L、Na2HPO4·12H2O 5 g/L、MgSO40.2 g/L、MnSO4·H2O 50 mg/L、CaCO330 g/L時對戊糖乳桿菌 ATCC8041發酵產乳酸的影響較大。在此基礎上,選取這7個對乳酸產量影響較明顯的因素進行Plackett-Burman設計試驗(表1),得到了2個對乳酸產量影響顯著的因素:葡萄糖質量濃度和酵母粉質量濃度,結果見圖2。在此基礎上對葡萄糖、酵母浸粉以及第三影響因子CaCO3做響應面試驗(表2),得到了預測乳酸產量的模型:
Y=54.00+1.62X1+0.33X2-0.020X3-0.075X1X2-0.020X1X3+0.020X2X3-2.58X12-0.79X22-0.22X32
其中X1為葡萄糖質量濃度,X2為酵母浸粉質量濃度,X3為CaCO3質量濃度。
對該模型進行方差分析和顯著性檢驗(表3)得出,葡萄糖質量濃度、酵母浸粉質量濃度兩個因素對乳酸產量影響均達到顯著水平。各因素影響乳酸產量的大小順序為:葡萄糖>酵母浸粉>CaCO3。二次回歸模型的R2為0.9997,方程P值<0.05,失擬檢驗P值為0.2500>0.05,失擬不顯著,試驗結果可靠。因此模型選擇正確,可用于預測乳酸產量。
Design-Expert軟件預測和驗證試驗表明,發酵培養基的最佳組合為:葡萄糖93.11 g/L、酵母浸粉5.19 g/L、碳酸鈣29.43 g/L、蛋白胨10.00 g/L、Na2HPO4·12H2O 5.00 g/L、MgSO40.20 g/L、MnSO450 mg/L。
2.2 戊糖乳桿菌ATCC 8041最佳發酵條件的確定
在發酵培養基優化的基礎上,對發酵條件做單因素試驗,結果(圖3-圖7)顯示,得到較佳發酵條件為:溫度37℃、初始pH6.5、接種量6%、裝液量80%。在培養基與發酵條件優化的基礎上,得到出發菌株發酵葡萄糖產乳酸產量為54.11 g/L。

表1 Plackett-Burman試驗設計與結果

圖2 七個因素的效應Pareto圖分析
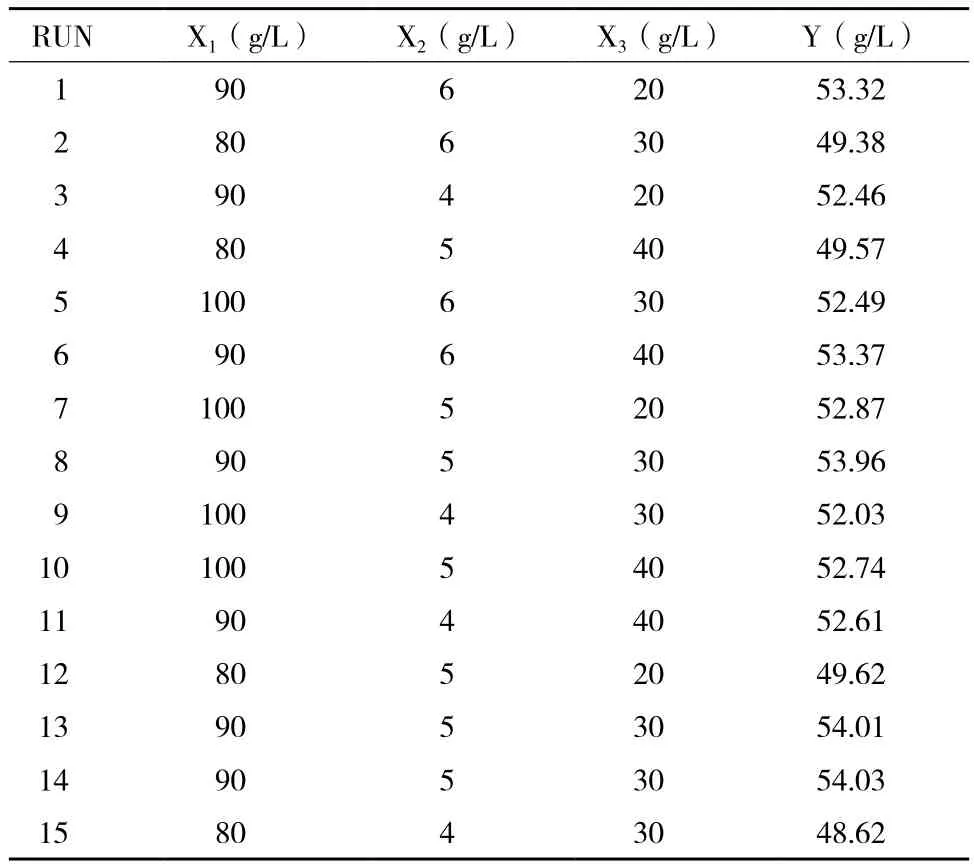
表2 響應面試驗設計及結果
2.3 戊糖乳桿菌ATCC 8041最佳誘變劑量的確定
戊糖乳桿菌ATCC 8041最佳紫外誘變劑量的確定結果(圖7)顯示,隨著紫外照射時間的增加,菌體的致死率有逐漸增加的趨勢。當照射時間為40
s時,菌體的致死率為85.54%,而60 s時紫外照射的致死率為97.84%。研究表明,當致死率為80%-90%之間時,菌體的誘變效果和生存能力的比例最好,從而確定在功率20 W的紫外燈,距離35 cm照射條件下,最佳誘變劑量為40 s。
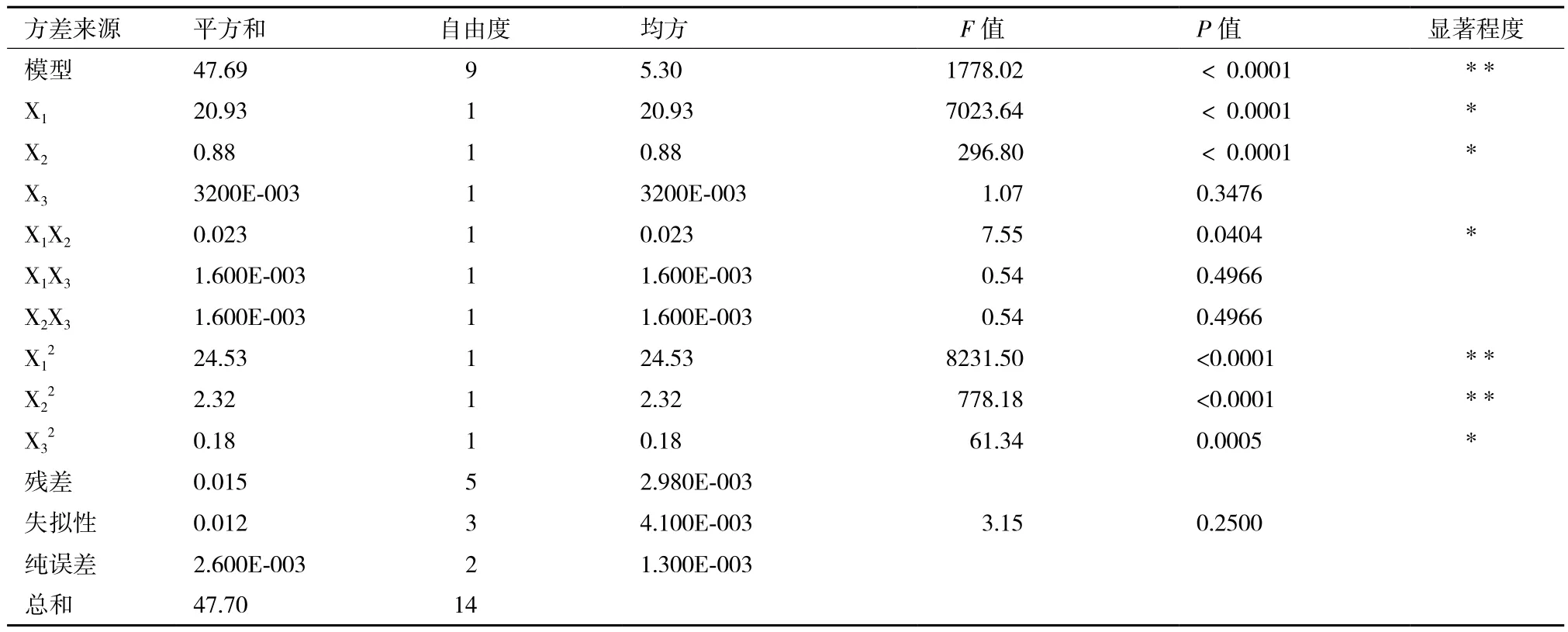
表3 方差分析及顯著性檢驗
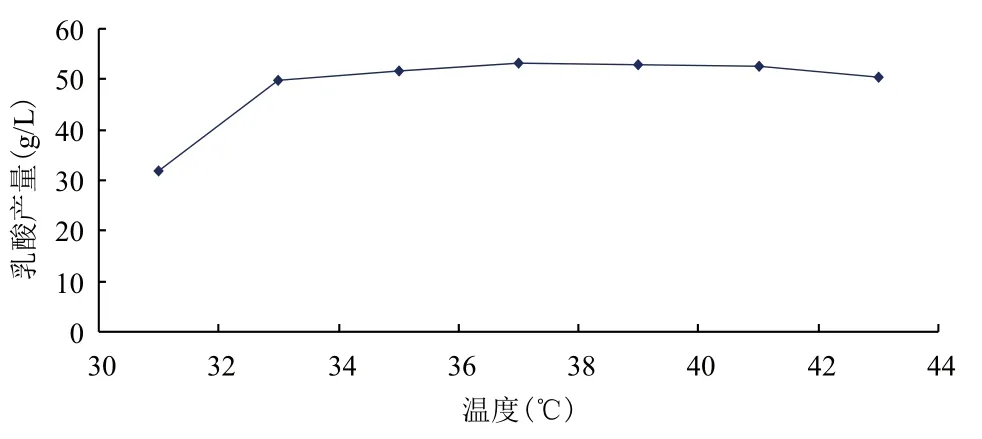
圖3 溫度對乳酸產量的影響
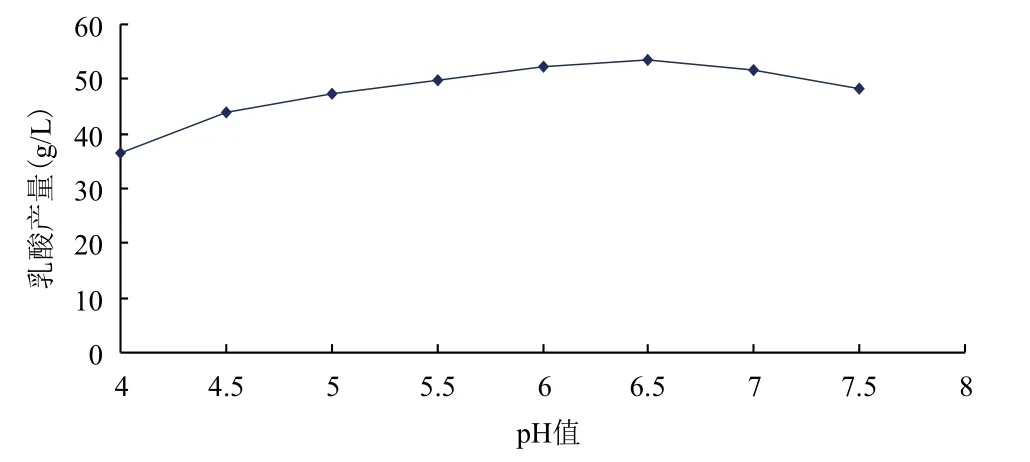
圖4 初始pH對乳酸產量的影響

圖5 接種量對乳酸產量的影響
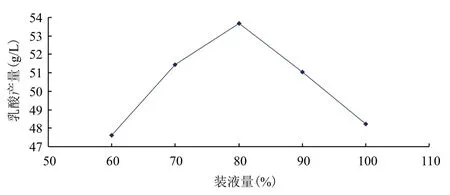
圖6 裝液量對乳酸產量的影響
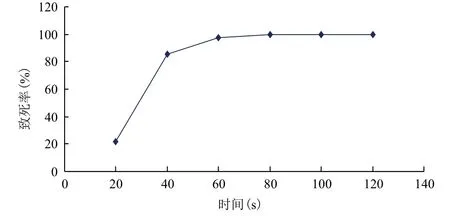
圖7 Lactobacillus pentosus ATCC 8041紫外致死率曲線
2.4 高產乳酸突變株的選育
出發菌株經紫外誘變后經篩選培養基初篩,觀察篩選平板,正常產酸菌落會產生黃色顯色圈(圖
8-B),挑取黃色較深的90株菌落進行復篩,結果(表4)顯示,紫外誘變后正突變為32.22%,負突變為28.89%,38.89%的菌株沒有明顯差異。在正突變菌株中,平均產量提高幅度為7.4%,最大產量提高率達到18.84%。進一步對正突變菌株進行搖瓶發酵測乳酸產量進行再復篩,結果(表5)顯示,經過再次復篩,突變株Lactic UVC-02和Lactic UVD-05產量較對照高15%以上。故對突變株Lactic UVC-02和Lactic UVD-05進行傳代培養,檢驗其遺傳穩定性。
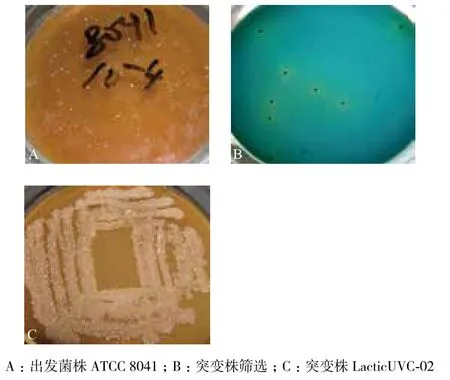
圖8 出發菌株和突變株菌落形態
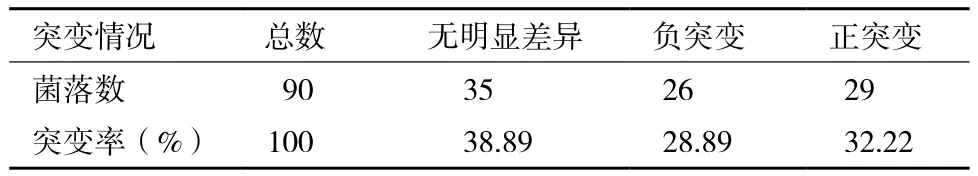
表 4 出發菌株紫外誘變結果
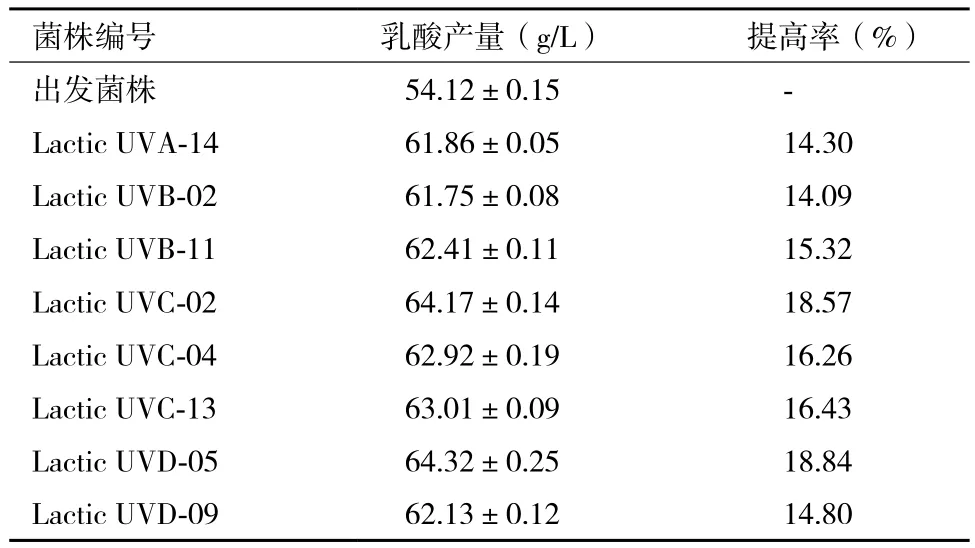
表5 突變株發酵復篩結果
2.5 突變菌株的遺傳穩定性
將突變株Lactic UVC-02進行傳代培養,傳代8次,測定其乳酸產量(每代重復5次)如表6所示。進一步對數據進行方差分析,結果見表6。由方差分析可以看出,F crit=2.31是α=0.05的F統計量臨界值,F=1.58是F統計量的計算值,1.58<2.31,P=0.177,故數據間無顯著差異,說明突變株Lactic UVC-02遺傳穩定性好。

表6 突變株Lactic UVC-02遺傳穩定性及方差分析
將突變株Lactic UVD-05進行傳代培養,傳代8次,其乳酸產量如表7所示(每代重復5次)。對數據進行方差分析,見表7。由方差分析可以看出,F crit=2.31,是α=0.05的F統計量臨界值,F=4.64是F統計量的計算值,4.64>2.31,P=0.001,數據間差異顯著,說明突變株Lactic UVD-05的遺傳穩定性差。
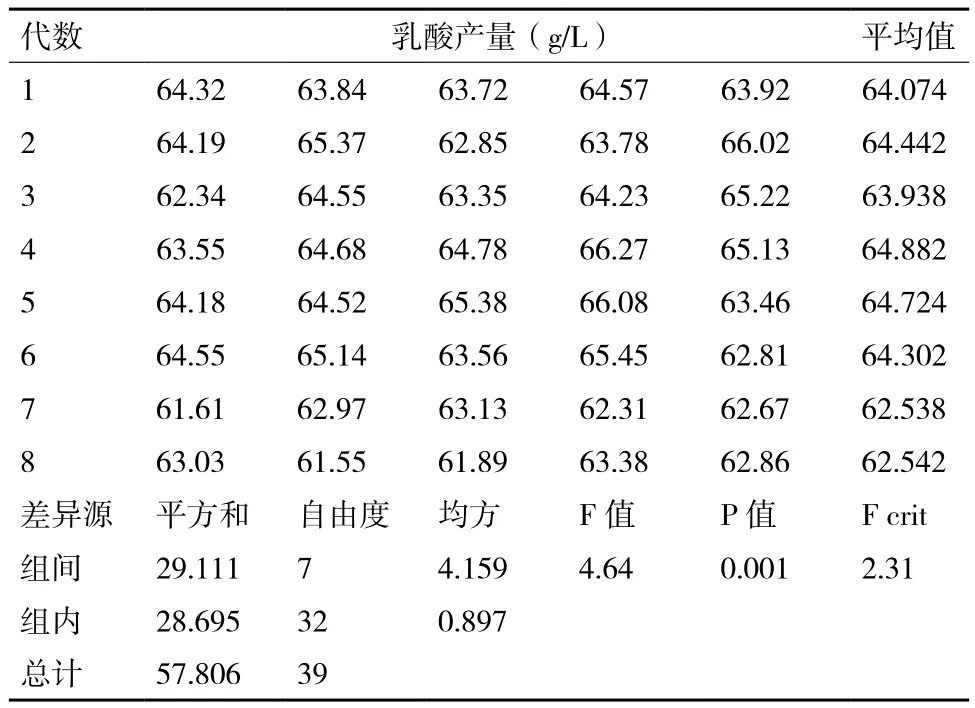
表7 突變株Lactic UVD-05遺傳穩定性及方差分析
通過對突變株Lactic UVC-02和Lactic UVD-05的遺傳穩定性比較表明,Lactic UVC-02的穩定性
優于Lactic UVD-05,故確定突變株Lactic UVC-02為一株高產乳酸突變株,命名為戊糖乳桿菌Lactic UVC-02,由中國典型培養物保藏中心保存,注冊號為CCTCC M 2013209。該突變株Lactic UVC-02經葡萄糖發酵后,乳酸產量達到64.17 g/L,比出發菌株(54.12 g/L)提高18.6%,為進一步利用木質纖維發酵產乳酸提供了研究基礎。
3 討論
在發酵工業中,發酵培養基的優化對發酵水平的提高有著重大作用,而合理的試驗設計及統計方法的選擇在探索最佳發酵培養基的過程中尤為重要[19]。Plackett-Burman(P-B)法是一種以不完全平衡塊為原理的試驗設計,它能夠從眾多的過程變量中快速、有效地篩選出最為重要的幾個因素,為進一步研究提供基礎。響應面分析法是1951年Box Wilson開發的用于研究多因子系統中因子交互作用達到最大響應值時所對應的最佳條件,結合數學方法和統計分析,通過對重要因子響應過程變量進行數學建模分析,定向優化響應因子[21]。國內外很多學者通過P-B設計和響應面分析法在乳酸發酵培養基優化方面取得了很好的成就[22-27]。本研究通過P-B設計快速有效地從7個影響乳酸產量的發酵培養基組分中篩選出3個顯著性影響組分:葡萄糖、酵母浸粉和碳酸鈣,然后利用響應面分析法中Box-Behnken設計得出最佳培養基組合:葡萄糖93.11 g/L、酵母浸粉5.19 g/L、碳酸鈣29.43 g/L、蛋白胨10 g/L、Na2HPO4·12H2O 5 g/L、MgSO40.2 g/L、MnSO450 mg/L。在最佳培養基組合下,乳酸產量達到54.11 g/L,與預測值擬合較好。說明運用響應面法優化戊糖乳桿菌ATCC8041發酵產乳酸是合理可靠的。
微生物誘變育種是指利用各種誘變劑處理微生物細胞,以提高其基因突變頻率,再通過適當的篩選方法獲得所需的高產優質菌種的育種方法,誘變育種的速度快、成本低、耗時短、方法簡便,是一種高效的菌株選育方法。其中的紫外線輻射方法更具有操作簡便,經濟實惠,對實驗室條件要求低,且出現正突變的幾率較高,誘變的效果好,是一種非常實用的誘變劑[28]。在微生物發酵產乳酸方面,利用紫外誘變篩選優良菌株也取得了良好的效果。仲松等[29]以干酪乳桿菌ZZ-L為出發菌株,對其原生質體進行紫外誘變,篩選得到一株比原始菌株乳酸產量高20.7%的高產突變菌株ZZ-06。吳慧昊等[30]使用紫外照射和亞硝基胍復合誘變的方法選育出一株在低溫下生長良好、遺傳性狀穩定的菌株Q1-4-6。Yin等[31]采用紫外和亞硝基胍復合誘變的方法得到突變株Rhizopus oryzae LA-UN-1,乳酸產量達到59.5 g/L,比原始菌株提高54.5%。本研究以能同時利用六碳糖和五碳糖的戊糖乳桿菌ATCC 8041為出發菌,制備了原生質體,并對其原生質體懸液進行紫外誘變,期望獲得能利用木質纖維水解液發酵產乳酸的高產突變株。突變體經過初篩、復篩等一系列過程篩選出乳酸產量較高的2個菌株:Lactic UVC-02與Lactic UVD-05,對其進行了8代繼代培養檢測其遺傳穩定性,其中突變株Lactic UVC-02遺傳穩定性較好,各代之間無顯著差異。突變株菌落形態與原始菌株也出現一定程度的差異,引起這種差異的原因還需進一步在分子水平進行分析研究。由于木質纖維原材料處理過程復雜且周期較長,目前我們已初步得出突變株Lactic UVC-02能利用楊木水解液發酵產乳酸,但暫時還未能完全驗證該突變株是否能高效利用木質纖維水解液發酵產乳酸,這也是我們下一步工作的研究重點。
4 結論
戊糖乳桿菌ATCC 8041發酵產乳酸的最佳發酵培養基和發酵條件組合為:葡萄糖93.11 g/L、酵母浸粉5.19 g/L、碳酸鈣29.43 g/L、蛋白胨10.00 g/L、Na2HPO4·12H2O 5.00 g/L、MgSO40.20 g/L、MnSO450 mg/L;最佳發酵條件為溫度37℃、pH6.5、接種量6%、裝液量80%。進一步對戊糖乳桿菌ATCC8041進行原生質體誘變得到高產突變株Lactic UVC-02,保存在中國典型培養物保藏中心(注冊號:CCTCC M 2013209),其乳酸產量達到64.17 g/L,比原始菌株產量提高了18.6%。
[1]曹本昌, 徐建林, 匡群.根霉發酵L-乳酸[J].食品與發酵工業, 1991(1):37-41.
[2]金其榮, 金豐收. 乳酸衍生物發展應用新動向[J]. 山西食品
工業, 2002, 3:2-4.
[3]錢志良, 胡軍, 雷肇祖. 乳酸的工業化生產應用和市場[J].工業微生物, 2001(6):49-54.
[4]曾煒, 陳豐秋, 詹曉力. 乳酸的生產技術及其研究進展[J].化工進展, 2006, 25(7):746-747.
[5]張剛, 乳酸細菌——基礎、技術和應用[M]. 北京:化學工業出版社, 2007.
[6]樂曉潔, 王昌祿, 顧曉波, 等. 細菌發酵生產L-乳酸高產菌株的選育[J]. 中國食品添加劑, 2004, 1:67-69.
[7]鄭艷, 薛景珍, 劉長江. L-乳酸發酵菌株的選育[J]. 微生物學雜志, 2004, 24(3):26-28.
[8]張秀鳳, 李正英, 江均平. 細菌L-乳酸發酵的研究[J]. 食品與機械, 2005, 21(5):74-76.
[9]白冬梅, 趙學明, 胡宗定. 反相HPLC雙檢測器法同時測定米根霉乳酸發酵液中的有機酸與葡萄糖[J]. 食品與發酵工藝, 2001, 27(1):13-17.
[10]付曉芬, 江均平, 張潔, 等. 微生物利用木糖發酵L—乳酸代謝途徑的研究[J]. 食品工業科技, 2009(8):359-362.
[11]Masayuki T, Masahiro H, Tanabe S, et al. Production of L-lactic acid by simultaneous saccharification and fermentation using unsterilized defatted rice bran as a carbon source and nutrient components[J]. Food Sci Technol Res, 2005, 11:400-406.
[12]Zhu Y, Lee YY, Elander RT. Conversion of aqueous ammoniatreated corn stover to lactic acid by simultaneous saccharification and co-fermentation[J]. Appl Biochem Biotechnol, 2007(1-12):137-140.
[13]Bustos G, Moldes AB, Cruz JM, et al. Production of fermentable media from trimming wastes and bioconversion into lactic acid by Lactobacillus pentosus[J]. Journal of the Science of Food and Agriculture, 2004, 84:2105-2112.
[14]張麗麗, 沈兆兵, 史吉平, 等. 紫外誘變和丁醇馴化復合選育高產丁醇菌株[J]. 中國釀造, 2013, 32(5):129-133.
[15]Bustos G, Moldes AB, Cruz JM, et al. Influence of the metabolism pathway on lactic acid production from hemicellulosic trimming vine shoots hydrolyzates using Lactobacillus pentosus[J]. Biotechnol Prog, 2005, 21:793-798.
[16]楊承劍, 黃興國, 等. Plackett-Burman設計在益生菌生長主要影響因子篩選中的應用[J]. 飼料工業, 2007, 28(16):31-33.
[17]Pan CM, Fan YT, Xing Y, et al. Statistical optimization of process parameters on biohydrogen production from glucose by Clostridium sp. Fanp2[J]. Bioresour Technol, 2008, 99(8):3146-3154.
[18]Yuan LL, Li YQ, Wang Y, et al. Optimization of critical medium components using response surface methodology for phenazine-1-carboxylic acid production by Pseudomonas sp. M-18Q[ J]. J Biosci Bioeng, 2008, 105(3):232-237.
[19]代志凱, 張翠, 阮征. 試驗設計和優化及其在發酵培養基優化中的應用[J]. 微生物學報, 2010, 37(6):894-903.
[20]白冬梅, 趙學明, 胡宗定. 高效液相色譜手性流動相添加劑分離乳酸對應體[J]. 分析化學, 2001, 29(4):413-415.
[21]田泱源, 李瑞芳. 響應面法在生物過程優化中的應用[J]. 食品工程, 2010(2):8-11.
[22]潘麗軍, 龐銳, 吳學鳳, 等. 葡萄糖和木糖共發酵生產L-乳酸的培養基和培養條件響應面優化[J]. 食品科學, 2011, 32(9):140-145.
[23]丁涓, 魏敏, 張莉. 玉米漿發酵生產 L-乳酸的工藝優化[J].食品科學, 2011, 32(1):127-130.
[24]秦浩. 高產L-乳酸菌株的選育及其發酵條件的研究[D]. 無錫:江南大學, 2012.
[25]Naveena BJ, Altaf M, Bhadriah K, et al. Selection of medium components by Plackett-Burman design for production of l(+)lactic acid by Lactobacillus amylophilus GV6 in SSF using wheat bran[J]. Bioresource Technology, 2005, 96(4):485-490.
[26]Coelho LF, De Lima CJB, Rodovalho CM, et al. Lactic acid production by new Lactobacillus plantarum LMISM6 grown in molasses:optimization of medium composition[J]. Brazilian Journal of Chemical Engineering, 2011, 28(1):27-36.
[27]Gowdhaman D, Sugumaran KR, Ponnusami V. Optimization of lactic acid production from tea waste by Lactobacillus plantarum MTCC 6161 in solid state fermentation by central composite design[J]. Inter J Chemtech Res, 2012, 4(1):143-148.
[28]石彩蕊, 王義強, 陳介南, 等. 產β-葡萄糖苷酶微生物育種研究進展[J]. 生物技術通報, 2011(3):59-65.
[29]仲松, 張慶慶, 孫平平. 紫外誘變原生質體選育高產L-乳酸菌株的研究[J]. 安徽工程大學學報, 2011, 26(1):28-30.
[30]吳慧昊, 牛鋒. 乳酸菌低溫菌株的復合誘變選育[J]. 微生物學通報, 2013, 40(4):631-645.
[31]Yin L, Ruan Q, Fu Y. Strain improvement of Rhizopus oryzae for over-production of lactic acid by random mutations[J]. African Journal of Microbiology Research, 2013, 7(23):2970-2975.
(責任編輯 馬鑫)
Lactobacillus pentosus Fermentation Lactic Acid Production and High Yield Strain Mutation Breeding
Wang Yiqiang1,2,3Wang Qiye1Ma Guohui1Lin Liyun1
(1. Biotechnology Laboratory of Central South University of Forestry and Technology,Changsha 410004;2. Key Lab of Non-wood Forest Nurturing and Protection of National Ministry of Education,Changsha 410004;3. Bio-ethanol Research Center of State Forestry Administration,Changsha 410004)
Lactobacillus pentosus is a potential strain which can make use of the lignocellulose hydrolysate fermentation to produce lactic acid, optimization of fermentation conditions and the breeding of high yield strain are the important means to improve lactic acid production. Fermentation medium and fermentation conditions were optimized for strain Lactobacillus pentosus ATCC 8041, through single factor experiment, Plackett-Burman design and response surface experiment. The results showed that the best combination of fermentation medium is 93.11 g/L glucose, 5.19 g/L yeast powder, 29.43 g/L calcium carbonate, 10.00 g/L peptone, 5.00 g/L Na2HPO4·12H2O, 0.20 g/L MgSO4, 50 mg/L MnSO4. The best fermentation conditions is temperature 37℃, pH6.5, quantity of 6%, and fluid amount 80%. The lactic acid production is 54.12 g/L under the optimized fermentation medium and fermentation conditions. And then, Lactobacillus pentosus ATCC 8041 was taken as the original strain, treated with UV mutation ATCC 8041 protoplast, after through multiple screening, finally we have got a mutant which has high lactic acid production, named Lactobacillus pentosus lactic UVC-02 which conserved in China Center for Type Culture Collection, registration number for CCTCC M 2013209. The results for glucose fermentation showed that lactic UVC-02 produced 64.17 g/L lactic acid, which was 18.6% higher than the original strain.
Lactobacillus pentosus Lactic acid fermentation UV mutation Response surface experiment
2014-03-27
國家林業局“948“項目(2011-4-13)
王義強,男,教授,博士生導師,研究方向:細胞工程與生物能源;E-mail:wangyiqiang12@163.com
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
河北農機(2020年10期)2020-12-14 03:13:30
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
今日農業(2019年15期)2019-01-03 12:11:33
聚氯乙烯(2018年9期)2018-02-18 01:11:34
新農業(2017年22期)2018-01-03 05:46:00
現代農業(2016年5期)2016-02-28 18:42:33
