高寒生態脆弱區不同擾動生境草地植被及土壤無機氮變化特征
2014-03-26 08:38:28張玉霞姚拓王國基馬文文馬文彬
草業學報 2014年4期
張玉霞,姚拓,王國基,馬文文,馬文彬
(甘肅農業大學草業學院 草業生態系統教育部重點實驗室,甘肅 蘭州730070)
生態脆弱區(生態交錯區)是兩種不同類型生態系統交界過渡區域。高寒生態脆弱區則是不同類型生態系統交界過渡區域出現在了海拔較高地區[1]。高寒生態脆弱區生態系統結構簡單、穩定性差、生產力水平低、易受外界因子干擾、修復能力弱,植被主要優勢種以寒旱叢生禾草為主,并混生有不同數量的高原灌叢、墊伏植物及毒草,總蓋度較低[2]。人類活動的干擾亦可造成高寒草地的退化,導致地表下墊面的改變,區域水熱循環過程失衡,加速草地的退化演替進程[3]。長期持續性過度放牧干擾造成了不同程度草地退化,地上植物量和草地植被總蓋度隨著草地退化程度加劇而逐漸降低。植物群落優勢種嵩草(Kobresiaspp.)、珠芽蓼(Polygonumviviparum)等逐漸被多莖委陵菜(Potentillaemulticaulis)、棘豆(Oxytropisspp.)、龍膽(Gentianaspp.)和馬先蒿(Pedicularisspp.)等毒雜草代替,同時其他毒草伴隨生長。
土壤含氮化合物主要以有機態存在,無機態氮一般占全氮1%~5%。但有機態氮大部分必須經過土壤微生物的轉化作用形成無機態氮,才能被植物吸收利用。無機態氮主要存在形式是硝態氮(NO3--N)、銨態氮(NH4+-N),有時有少量亞硝態氮存在,是植物直接吸收利用的重要地下氮素寶庫,在農、林業可持續發展中扮演重要角色。有機氮經微生物氨化作用,可以釋放出氨,氨可以進一步轉化為銨,形成銨態氮。同時,在適宜條件下,銨態氮可以經微生物硝化作用轉化為硝酸鹽,形成硝態氮。已有研究表明,植物能吸收利用的氮素形態有硝態氮、銨態氮、亞硝態氮、單質態氮以及一些可溶性有機含氮化合物[4]。土壤中氮素經根系吸收進入植物體內,但作物能否高效吸收氮素與根系性狀有密切聯系[5]。無論低氮或高氮環境中,植物根系都形成了特有的適應機制[6]。土壤銨態氮、硝態氮不斷被植物吸收利用,養分被轉移,植被蓋度越大,被利用的養分也越多。李貴才等[7]認為不同生態系統的植被類型、微生物種類差異是導致土壤礦化量變化的原因。硝態氮含量季節變化趨勢差異性大,隨土層加深差異越來越不明顯,反映了植被類型對表層土壤氮素季節動態變化起到決定性作用[8]。因此其土壤銨、硝態氮含量高低受有機質含量、微生物種類和土壤環境條件如土壤溫度、濕度和鹽堿度等諸多因素影響[9]。顏淑云等[10]對瑪曲高寒草地不同利用方式下土壤氮素含量特征進行了研究,土壤無機氮與全氮線性相關,土壤全氮與銨態氮、硝態氮也呈線性關系。史作民等[11]對內蒙古鄂爾多斯地區4個植物群落類型的土壤碳氮特征進行研究。李榮等[12]對沙丘固定過程中土壤銨態氮和硝態氮的時空變化進行分析,發現土壤緊實可以減少硝態氮的淋失,但影響了微生物活動,間接影響銨、硝態氮含量。胡璐等[8]對中國北方蒙古干旱半干旱區土壤銨態氮分布及其環境控制因素進行分析,發現隨著土壤pH值的升高,其表層銨態氮含量減少。施用氮肥可以顯著影響土壤中有機碳類物質的含量,隨施氮濃度增加,各土層中總有機氮含量、土壤微生物量氮含量呈上升趨勢,水溶性有機氮含量呈下降趨勢[13]。梁飛等[14]通過田間人工種植試驗證明追施氮肥可以提高鹽地堿蓬(Suaeda salsa)整株生物量,促進鹽地堿蓬對鹽漬地的生物修復。在草地生態系統中,氮是牧草生長的重要限制因素之一,而有關高寒草地植被及無機氮素變化特征研究報道相對較少。本研究通過對天祝高寒草地植被的調查及不同干擾生境下土壤無機氮含量動態變化研究,揭示不同干擾生境下土壤無機氮素含量的時空變化特征,分析造成土壤無機氮素含量變化的原因,為高寒生態脆弱區植被恢復和可持續利用提供理論依據。
1 材料與方法
1.1 研究地自然概況
研究地位于甘肅省天祝藏族自治縣金強河流域甘肅農業大學高山草原試驗站,其地理坐標為37°11′~37°13′N、102°29′~102°33′E,平均海拔2700~3300m,氣候陰冷潮濕,空氣稀薄,紫外線輻射強烈。根據相關記錄,年均溫-0.1℃,1月均溫-18.3℃,7月均溫12.7℃,>0℃年積溫1380℃,水熱同期,年日照時數2600h;年降水量416mm,多為地形雨,集中于7-9月,年蒸發量1592mm。無絕對無霜期,僅分冷、熱兩季,春季常有旱象,并有暴風雪。土層厚約40~80cm。土壤pH值7.0~8.2,有機質含量10%~16%。土壤以亞高山草甸土、亞高山黑鈣土等為主。植被以嵩草、苔草(Carexspp.)、針茅(Stipacapillata)、莎草(Cyperusspp.)、珠芽蓼、金露梅(Potentillafruticosa)、棘豆、狼毒(Stellerachamaejasme)、委陵菜(Potentillachinensis)、杜鵑(Rhododendronspp.)、火絨草(Leontopodium)、黃芪(Astragalusspp.)、麻花艽(Gentianastraminea)、高山唐松草(Thalictrumalpinum)、甘肅馬先嵩(Pediculariskansuensis)、早熟禾(Poaspp.)等為主。
1.2 樣品采集
1.2.1 樣點選擇 為研究不同干擾生境下高寒生態脆弱區草地土壤無機氮動態變化,在海拔2970~3000m處,以目前高寒草地的幾種常見干擾生境為主,選取放牧強度小的圍欄內天然草地(圍欄內)、放牧強度大的圍欄外天然草地(圍欄外)及牧道3種干擾方式的草地,具體樣地特點見表1。

表1 樣地類型及干擾特點Table 1 Types and characteristics of sample site
1.2.2 植被調查[15]在研究樣地不同干擾生境內隨機選取5個1m×1m的樣方,辨別記錄樣方內植物種類、種數;以10cm為單位將樣方分為100個網格,采用針刺法在各交叉點刺探測試有無植物根系著生,有植物即算作1,調查植被蓋度;并將樣方內的所有植物齊地面刈割,除去粘附的土壤、礫石等雜質后稱取鮮重,記錄樣方地上生物量;在3種樣地內選擇5株有代表性的植株測量自然高度,重復10次,調查植被高度。
1.2.3 土樣采集 分別在2012年4月,6月,8月,10月,在上述3種干擾生境內采用土樣剖面取樣法取樣,相同月份盡量考慮地形基本一致,植物生長表現基本相同,隨機選取5個點,分0~10cm、10~20cm和20~30cm在同一垂直線上自下而上逐層采集土壤樣品,將土樣裝入采樣箱,放入冰袋暫存,帶回實驗室后存入4℃,并在一周內測定各指標。
1.3 土壤銨態氮(NH4+-N)和硝態氮(NO3--N)測定
1.3.1 土壤銨態氮測定方法——靛酚藍比色法[16]稱取過1mm篩的土樣10g,加KCl溶液50mL,振蕩0.5 h后過濾。取濾液5~10mL放入50mL容量瓶中,用KCl溶液補充至10mL,加入苯酚溶液和次氯酸鈉堿性溶液各5mL搖勻。在20℃左右的室溫下放置1h后,加掩蔽劑1mL并定容。在625nm波長處比色,記錄吸光度,根據標準曲線計算測定溶液質量濃度。結果計算:c=ρ×v×ts/m,式中,c為銨態氮濃度(mg/kg);ρ為銨態氮的質量濃度(μg/mL);v為顯色液的體積(mL);ts為分取倍數;m為干樣品質量(g)。
1.3.2 土壤硝態氮測定方法——雙波長紫外分光光度法[17]稱取過1mm篩的土樣10g,加0.2g CaSO4和100mL二次蒸餾水,振蕩15min后靜置30min過濾。取濾液50mL于50mL比色管中,之后轉移到三角瓶中再加1mol/L的鹽酸溶液1mL,在波長220和275nm處讀取吸光度,根據標準曲線計算測定液質量濃度。結果計算:c=c0×V×D×1000/(M×103),式中,c為硝態氮濃度(mg/kg);c0為測定液質量濃度(μg/mL);V為測定溶液體積(mL);D為分取倍數;M為干試樣質量(g);1000與103為單位換算數量級。
1.4 數據處理
所有圖表使用Excel軟件進行繪制,試驗數據采用SPSS進行統計分析。
2 結果與分析
2.1 不同干擾生境植被特征
對研究樣地植被進行調查,結果(表2)表明,天祝高寒草地中,雖然圍欄內和圍欄外植物種類相似度較高,均包含有嵩草、苔草、針茅、珠芽蓼、金露梅等,蓋度相近,但兩種干擾生境下優勢種、植被高度與地上生物量明顯發生了變化,圍欄內的高度、地上生物量分別是圍欄外的2.75和2.49倍,而且圍欄外草地的可食牧草減少,并以棘豆、狼毒等毒草為優勢種;牧道植物種數、優勢種、蓋度、高度及地上生物量相比于圍欄內外草地均明顯減少。
2.2 土壤銨態氮(NH4+-N)和硝態氮(NO3--N)空間變化
2.2.1 不同干擾生境銨態氮含量空間變化 3種干擾生境下土壤銨態氮含量的空間變化如圖1所示。結果表明,3種干擾生境下土壤銨態氮含量均隨著土層深度的增加而逐漸減小,不同生境類型NH4+-N含量均表現出0~10cm>10~20cm>20~30cm的趨勢。圍欄內0~10cm銨態氮含量最高(16.357mg/kg),顯著高于其10~20cm、20~30cm層,10~20cm與20~30cm層的銨態氮含量差異顯著;圍欄外0~10cm土層的銨態氮含量與10~20cm層的差異不顯著,與20~30cm層的差異顯著,10~20cm與20~30cm層差異不顯著;牧道各土層的銨態氮含量均低于10mg/kg,20~30cm層含量最低(4.328mg/kg),且各土層間差異顯著。
2.2.2 不同干擾生境硝態氮含量空間變化 圖2為3種不同干擾生境下土壤硝態氮含量空間變化。結果表明,3種干擾生境下土壤硝態氮含量亦隨著土層深度增加而逐漸減小,各干擾生境下NO3--N含量變化趨勢與NH4+-N的基本相似,也表現出0~10cm>10~20cm>20~30cm的趨勢。圍欄內0~10cm層硝態氮含量與20~30cm層的差異顯著,與10~20cm層的差異不顯著,10~20cm與20~30cm層差異不顯著;圍欄外0~10 cm層硝態氮含量最高(23.924mg/kg),顯著高于10~20cm和20~30cm層,10~20cm與20~30cm層的硝態氮含量差異顯著;牧道20~30cm層含量最低(3.029mg/kg),且各土層間差異顯著。
2.3 土壤銨態氮(NH4+-N)和硝態氮(NO3--N)季節變化
對圍欄內天然草地、圍欄外天然草地和牧道3種干擾生境下土壤表層(0~10cm)銨態氮和硝態氮的季節動態進行了研究,結果表明,3種干擾生境下土壤銨態氮和硝態氮含量具有明顯的季節變化特征(圖3、圖4)。
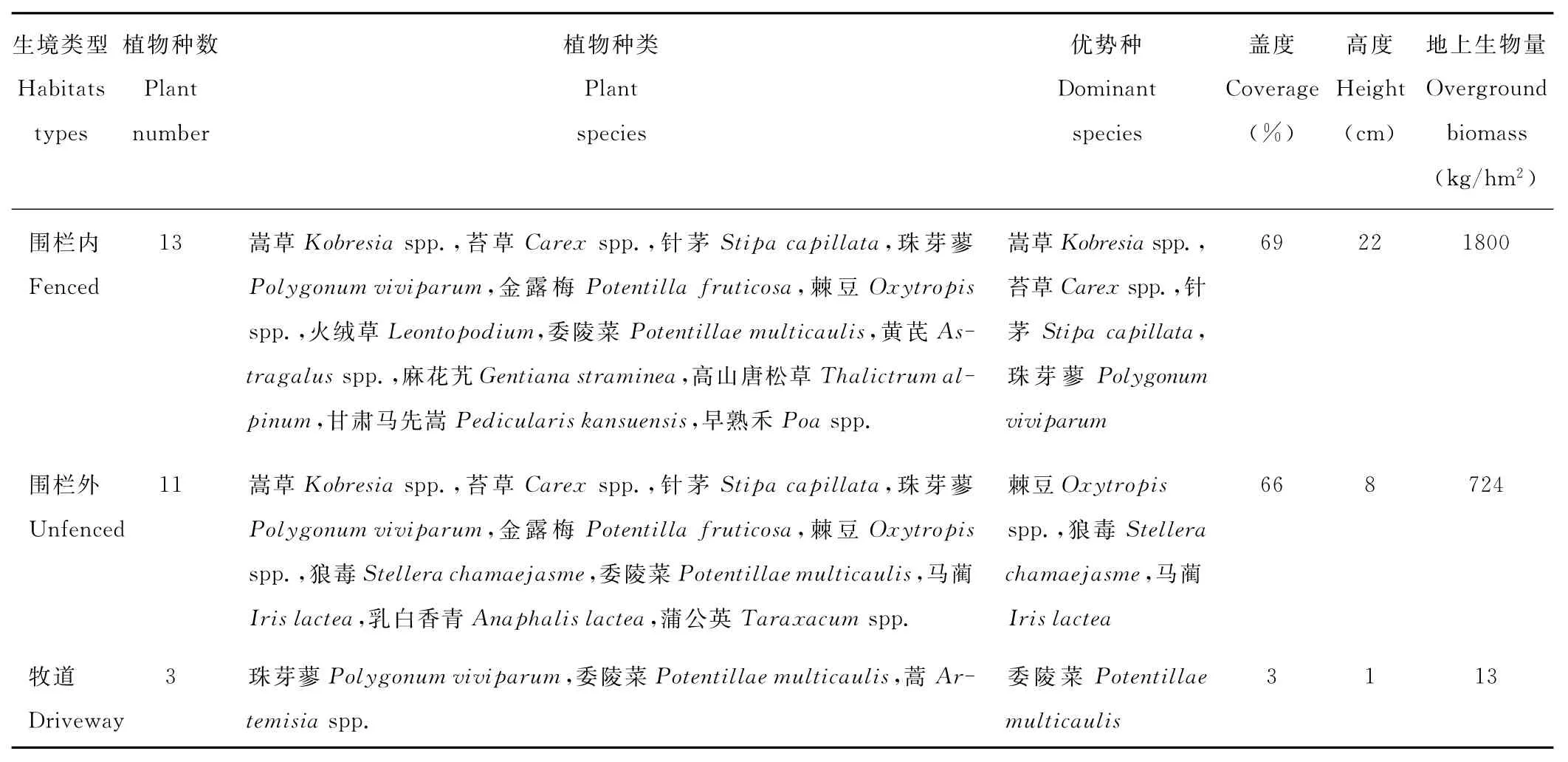
表2 不同干擾生境植被特征Table 2 Vegetation characteristics under different disturbed habitats
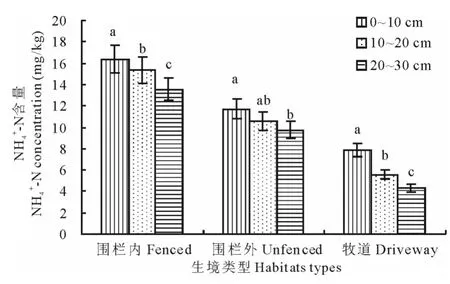
圖1 不同干擾生境下土壤銨態氮的空間變化Fig.1 Space change of ammonium-nitrogen under different disturbed habitats
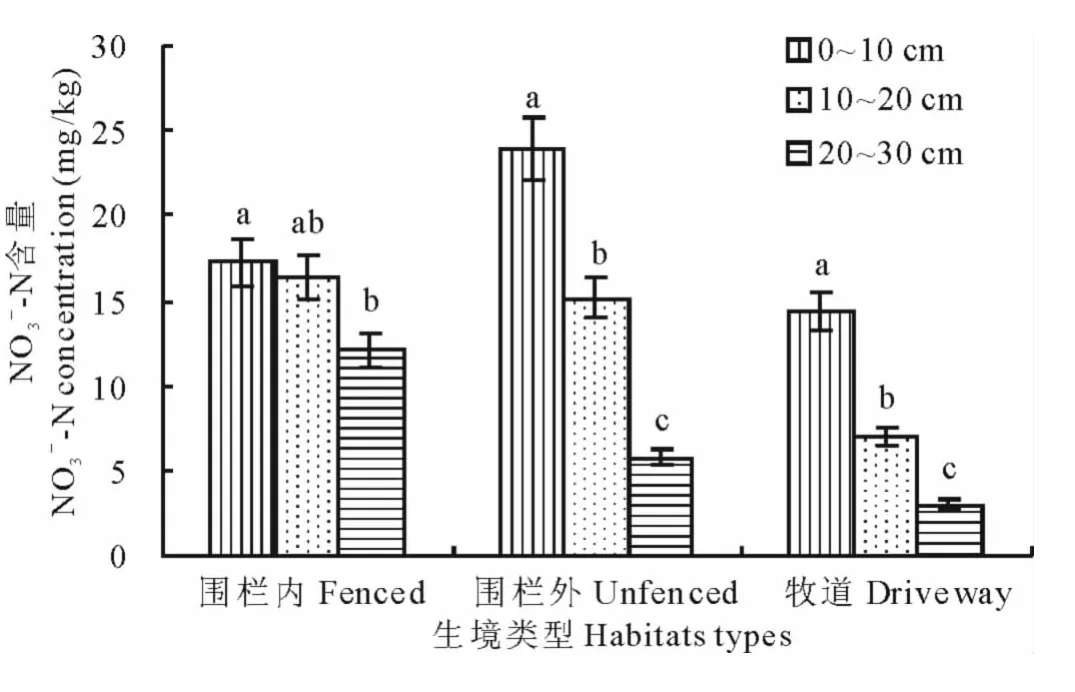
圖2 不同干擾生境下土壤硝態氮的空間變化Fig.2 Space change of nitrate-nitrogen under different disturbed habitats
2.3.1 不同干擾生境銨態氮季節動態變化 3種干擾生境下NH4+-N含量均表現出先增加后減少的趨勢(圖3),除10月圍欄內外,其他月份圍欄內、圍欄外、牧道的銨態氮含量季節變化趨勢基本一致,表現為8月>6月>10月>4月。圍欄內6月、8月、10月動態變化差異較小,10月相對于8月的硝態氮含量只下降0.4178mg/kg,4月含量最低(7.1551mg/kg)。圍欄外6、8月銨態氮含量較高,變化幅度較小,4月含量最低(3.1242mg/kg),10月相對于8月降低了11.3678mg/kg。牧道6月銨態氮含量比4月高出0.664mg/kg,8月份銨態氮含量最高(18.5058mg/kg),10月含量最低(2.3498mg/kg)。
2.3.2 不同干擾生境硝態氮季節動態變化 由圖4可以看出,4-8月3種干擾生境硝態氮含量逐漸升高,除10月圍欄外的持續上升外,其他2種干擾生境硝態氮含量8月以后降低,季節變化趨勢為8月>6月>4月>10月。圍欄內4,6,8月硝態氮含量變化幅度較小,10月含量最低(5.9450mg/kg),相比于8月降低了18.4748 mg/kg;圍欄外硝態氮含量持續增加,從4月的9.2869mg/kg上升到10月的40.3676mg/kg,但6,8月含量相近。牧道的硝態氮含量變化趨勢與圍欄內的相似,4月最低(7.4299mg/kg),8月最高(26.4973mg/kg),6,10月硝態氮含量只相差0.2739mg/kg。
2.4 土壤銨態氮(NH4+-N)和硝態氮(NO3--N)與草地植被相關性分析
以8月銨態氮、硝態氮含量分別與地上生物量做相關性分析(表3)。3種干擾生境下銨態氮與硝態氮同地上生物量之間均呈現正相關關系(P<0.05),但相關程度因草地類型而異。圍欄外、牧道銨態氮含量與地上生物量之間為中度相關,相關系數為0.6~0.8,圍欄內銨態氮含量與地上生物量之間為高度相關,相關系數大于0.8;圍欄內硝態氮含量與地上生物量間為中度相關,圍欄外、牧道硝態氮含量與地上生物量間為低度相關,相關系數為0.3~0.5。
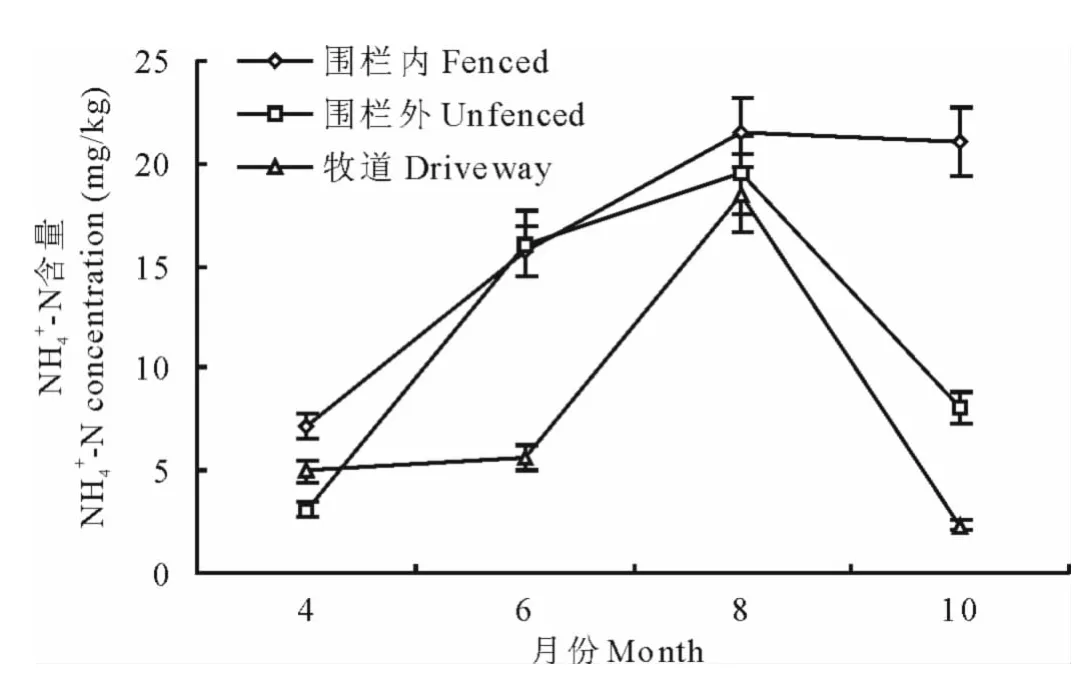
圖3 不同干擾生境下土壤銨態氮季節動態變化Fig.3 Seasonal dynamic of ammonium-nitrogen under different disturbed habitats
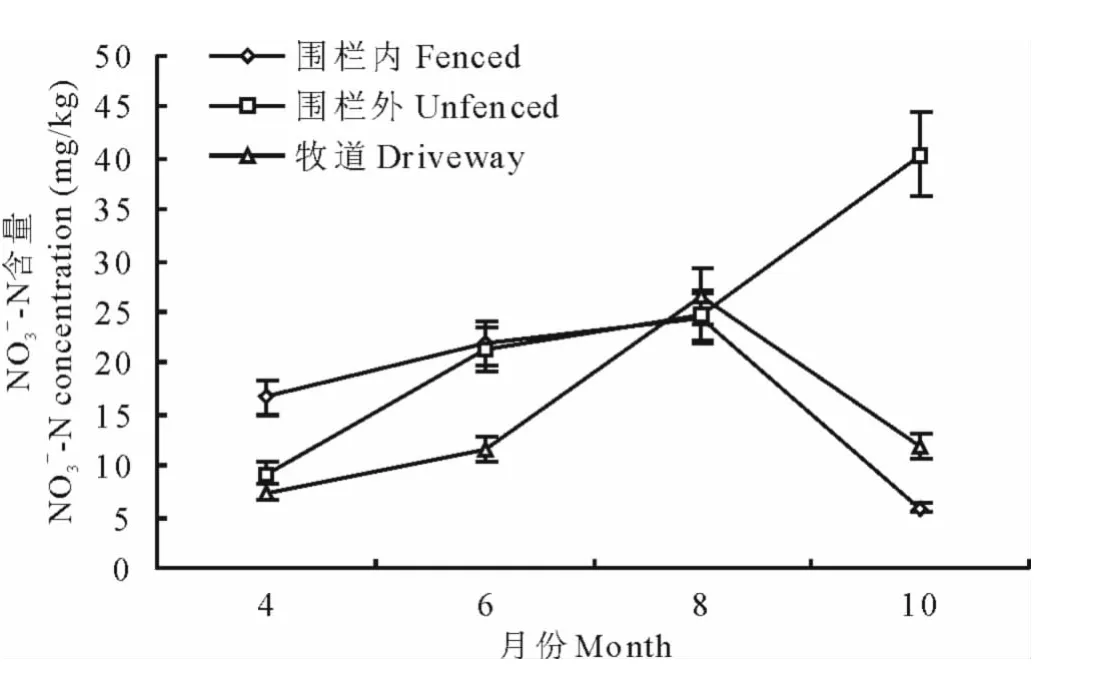
圖4 不同干擾生境下土壤硝態氮季節動態變化Fig.4 Seasonal dynamic of nitrate-nitrogen under different disturbed habitats
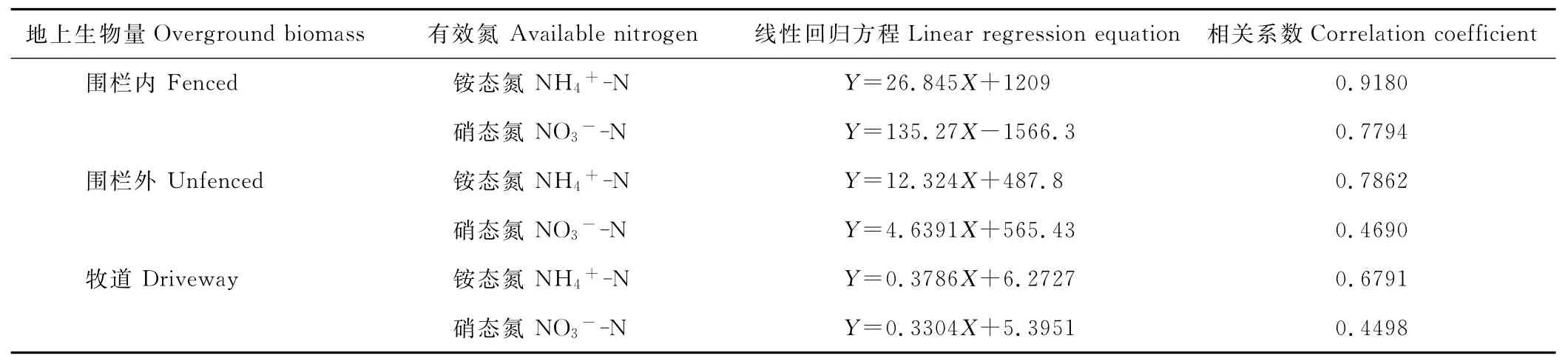
表3 土壤有效氮與地上生物量的相關分析Table 3 Correlation analysis between overground biomass and available nitrogen
3 討論
3.1 天祝高寒草地植被特征
禾本科、莎草科等可食牧草逐漸減少和雜類草蓋度增加的趨勢可以反映高寒草甸退化演替過程植被變化的基本特征。退化程度較輕的草地多處于禾草-矮嵩草群落時期,植被分為兩層結構,上層為禾本科牧草,下層為莎草屬牧草,隨退化程度的加重,草地植被逐漸演替為小嵩草、雜類草[3]。研究地天祝金強河流域平均海拔2700~3300m,氣候陰冷潮濕,年均溫-0.1℃,屬于典型的高寒生態脆弱區。植被以嵩草、苔草、針茅、珠芽蓼、棘豆、狼毒、黃芪、甘肅馬先嵩等為主,總蓋度較低,易受外界因子干擾,圍欄內天然草地除放牧外,無其他人類生產活動,劃區輪牧,放牧強度小,因此植被高度、地上生物量等都明顯高于其他干擾生境,且優勢種多為家畜的可食牧草。圍欄外天然草地由于長期過度放牧,退化嚴重,植物群落優勢種嵩草、苔草、珠芽蓼等被棘豆、狼毒、黃芪等毒雜草代替。牧道長年被家畜踐踏和排泄,土壤緊實、對植被種類、種數、優勢種、蓋度、高度及地上生物量都產生了很大影響。
3.2 土壤有效氮空間變化特征
土壤中的有效氮以銨態氮和硝態氮形式存在。一般來說,銨態氮在土壤中大多被吸附在土壤膠體表面,而硝態氮不被土壤膠體吸附,移動性較強。由于土壤中的NO3--N含量易受NH4+-N供給、硝化細菌數量、降水淋溶以及氣態釋放的影響,NO3--N含量比NH4+-N含量具有更大的不穩定性,其空間變異性很大。即使在相同干擾類型下,由于微域條件的變化,土壤NO3--N含量也明顯表現出不同[10]。同時,植物對于銨態氮和硝態氮的吸收具有明顯的偏向性[18],即土壤銨、硝態氮含量變化也受植被種類的影響。3種干擾生境下土壤銨態氮與硝態氮含量均隨著土層深度的增加而逐漸減小,此結果與史作民等[11]、孫景玲等[19]的研究結果一致。在牧道中,濕度是氮礦化的主要決定因素[20],即銨、硝態氮含量變化的主要原因。牧道家畜的活動量大,尤其是牧道的家畜糞便量增大,經家畜踩踏的牧道土壤緊實,容重增加,土壤空隙變小,好氣微生物數量減少,氮素少被分解,因此牧道0~10cm層的銨態氮含量較高。圍欄外各土層間硝態氮含量差異較大,出現這種狀況的原因可能是采樣時采到牛羊糞便的地盤或者植被蓋度較大的區域,通過硝化細菌的轉化,硝態氮含量增加。
3.3 土壤有效氮季節變化特征
NH4+-N和NO3--N的季節變化規律特征在以往的研究中多有出現[21-22],一般為夏秋季節高,春冬季節低[23-24]。本研究中NH4+-N及NO3--N的動態變化均符合此規律,原因在于溫度對氮礦化作用影響顯著,隨著溫度的季節性變化,土壤有效氮也隨著發生相應的變化。研究地3-5月干旱無雨,牧道長年被牲畜踩踏,土壤緊實,硝態氮不易被淋失[23],因此4月銨、硝態氮含量均較低,但銨態氮含量小于硝態氮。6月,8月硝態氮含量較高,這是由于研究地6月份天氣回暖,土壤溫度上升,濕度增大,7-9月降水多,導致土壤含水量增大,有效地促進氮礦化作用,尤其是NH4+-N的硝化作用。6月圍欄內和圍欄外銨態氮含量相近,可能是由于相似草地類型具有相近的植被組成、類似的生物群落,造成了它們相似的NH4+-N變化趨勢。8月牧道銨態氮含量與圍欄內、圍欄外的含量相似,是因為8月家畜的活動量大,尤其是牧道的家畜糞便量增大,促使牧道0~10cm的銨態氮含量的增加,此結果與孫志高等[25]的結果不一致。10月溫度降低以后植被生長緩慢或者停止生長,對土壤中的銨態氮利用較低,同時土壤中的微生物活動受到溫度的限制,轉化為硝態氮較少;而深秋季節降雨較多,圍欄內土壤相對疏松,增加了硝態氮的淋失[25]。
3.4 土壤銨態氮和硝態氮與草地植被相關性分析
作為直接氮源的銨、硝態氮與地上植物量的相關性強弱因草地類型不同而改變,3種干擾生境下銨態氮與硝態氮同地上生物量之間均呈現正相關關系,但相關程度因草地類型而異。隨著土壤銨、硝態氮含量減少,地上植物量也存在不同程度的減少,但其減少的幅度不同。地上生物量與圍欄外、牧道銨態氮含量之間為中度相關,與圍欄內銨態氮含量間為高度相關;地上生物量與圍欄內硝態氮含量間為中度相關,與圍欄外、牧道硝態氮含量間為低度相關。造成上述結果的原因很可能與干擾強度有關[26]。3種干擾生境間的區別是圍欄內草地能夠維持較為穩定的群落結構,具有較高的植物生產能力,屬于健康狀況良好的植被類型。圍欄外天然草地由于長期過度放牧干擾造成草地嚴重退化,地上植物量和草地植被總蓋度也明顯降低。牧道主要是家畜行走的通道,長期踐踏造成土壤緊實,微生物數量相對較少,植被蓋度很低,土壤中養分轉換、循環較慢。
4 結論
圍欄內和圍欄外植物種類相似度較高,蓋度相近,但兩種干擾生境下的優勢種、植被高度與地上生物量明顯發生了變化,牧道植物種數、優勢種、蓋度、高度及地上生物量相比于圍欄內外草地的均明顯減少。
相同干擾生境下的銨態氮和硝態氮含量均隨著土層深度增加而減少,且變化幅度較大,0~10cm層的含量與20~30cm層的存在顯著差異。相同土層3種干擾生境下銨態氮和硝態氮均表現出圍欄內>圍欄外>牧道的趨勢。
3種干擾生境下土壤NH4+-N的季節變化趨勢一致為8月>10月>6月>4月,10月份圍欄內的銨態氮含量與8月的差異不顯著;NO3--N的季節變化趨勢為8月>6月>4月>10月,10月圍欄外的硝態氮含量顯著高于其他月份相同生境含量。
3種干擾生境下銨態氮與硝態氮同地上生物量之間均呈現正相關關系,圍欄內銨態氮含量與地上生物量之間為高度相關,圍欄外、牧道硝態氮含量與地上生物量間為低度相關。
[1] 閻欣,甄峰,席廣亮.高寒生態脆弱地區城市綠色工業選擇與布局研究[J].經濟地理,2011,31(7):1139-1145.
[2] 劉金榮,謝曉蓉.祁連山高寒草地特點及可持續發展利用對策[J].草原與草坪,2002,2:15-18.
[3] 王建兵,張德罡,曹廣民,等.青藏高原高寒草甸退化演替的分區特征[J].草業學報,2013,22(2):1-10.
[4] 門中華,李生秀.水培硝態氮濃度對冬小麥幼苗氮代謝的影響[J].廣西植物,2010,4:77-79.
[5] 史正軍,樊小林,Klaus D,等.根系局部供氮對水稻根系形態的影響及其機理[J].中國水稻科學,2005,19(2):147-152.
[6] 郭亞芬,米國華,陳范駿,等.硝酸鹽供應對玉米側根生長的影響[J].植物生理與分子生物學學報,2005,31(1):90-96.
[7] 李貴才,韓興國,黃建輝.哀牢山木果柯林及其退化植被下土壤無機氮庫的干季動態特征[J].植物生態學報,2004,46(2):194-201.
[8] 胡璐,李心清,黃代寬,等.中國北方-蒙古干旱半干旱區土壤銨態氮的分布及其環境控制因素[J].地球化學,2008,37(6):572-580.
[9] Rasbid G H,Scbeafer R.Seasonal variation in the nitrogen mineralization and mineral nitrogen accumulation in the temperate forest soils[J].Pedobiologia,1988,31:335-347.
[10] 顏淑云,周志宇,鄒麗娜,等.瑪曲高寒草地不同利用方式下土壤氮素含量特征[J].草業學報,2010,19(2):153-159.
[11] 史作民,劉世榮,程瑞梅.內蒙古鄂爾多斯地區四個植物群落類型的土壤碳氮特征[J].林業科學,2004,40(2):21-27.
[12] 李榮,何興東,張寧,等.沙丘固定過程中土壤銨態氮和硝態氮的時空變化[J].土壤學報,2010,47(2):295-300.
[13] 朱毅,侯新村,武菊英,等.氮肥對兩種沙性栽培基質中有機氮類物質含量的影響[J].草業學報,2013,22(2):38-46.
[14] 梁飛,田長彥,田明明,等.追施氮肥對鹽地堿蓬生長及其改良鹽漬土效果研究[J].草業學報,2013,22(3):234-240.
[15] 牛翠娟,婁安如,孫儒泳,等.基礎生態學(第二版)[M].北京:高等教育出版社,2007:12.
[16] 杜森,高祥照.土壤分析規范[M].北京:中國農業出版社,2006:44-46.
[17] 梁蘭英.紫外分光光度法測定土壤中的硝態氮[J].甘肅環境研究與監測,2001,14(2):80-81.
[18] 田霄鴻,李生秀.幾種蔬菜對硝態氮、銨態氮的相對吸收能力[J].植物營養與肥料學報,2000,6(2):194-201.
[19] 孫景玲,王平,王德權.河西地區間作小麥土壤硝態氮含量時空動態變化分析[J].土壤通報,2010,41(4):882-885.
[20] Noy-Meir.Desert ecosystems:environment and producers[J].Annual Review of Ecology and Systematics,1973,4:25-51.
[21] Raghubanshi A S.Effect of topography on selected soil properties and nitrogen mineralization in a dry tropical forest[J].Soil Biology and Biochemistry,1992,24:145-150.
[22] 郁夢德,莫江明,孔國輝.離子交換樹脂袋法測定鼎湖山季風常綠闊葉林土壤有效氮的初步研究[J].熱帶亞熱帶植物學報,1995,34:44-48.
[23] 孟盈,甘建民,鄭征,等.熱帶雨林及在其下種植砂仁的土壤中氮素礦化和硝化的對比研究[J].農業環境科學學報,2003,22(1):21-24.
[24] 許翠清,陳立新,顏永強,等.溫帶森林土壤銨態氮、硝態氮季節動態特征[J].東北林業大學學報,2008,36(10):19-21.
[25] 孫志高,劉景雙,牟曉杰.三江平原小葉章濕地土壤中硝態氮和銨態氮含量的季節變化特征[J].農業系統科學與綜合研究,2010,26(3):277-282.
[26] 高海峰,白軍紅,王慶改,等.霍林河下游典型洪泛區濕地土壤pH值和土壤含水量分布特征[J].水土保持研究,2011,18(1):268-271.
