手是怎么形成的(四):決定肢體進化的基因
2014-05-03 08:14:24
中國三峽 2014年11期
手是怎么形成的(四):決定肢體進化的基因
文/卡爾·齊默 譯/靳 萌 編輯/吳冠宇
同源異型基因是整個動物王國進化遺產的一部分,它們能告訴我們生物進化是如何發生的。

中國演員在普希金莫斯科劇院表演芭蕾舞。舞蹈也是肢體語言的表達方式。 攝影/rna/CFP
一位名叫威廉·貝特森(William Bateson)的英國動物學家于19世紀90年代首先觀察到一組尤其引人注目的變異。一只葉蜂的腳長在了觸角上,一只螯蝦在眼睛部位長出了一根觸須,一只飛蛾原先的腿長成了翅膀。
圖靈模式決定肢體生長的理論遭到一些人的惡意批評。根據奧斯特建立的模型,在肢芽仍是相互一致的一批細胞時,作用于肢芽的各種力量集中創造了后來成為骨骼的聚合的模式,而不是創造了任何形式的決定將來肢體形狀的早期化學信號。1990年,英國的生物學家們設計了一項實驗來檢驗這兩種可能性。他們將一只小雞的翅膀的一半去除掉,將另一只翅膀的一半移植在該部位。因為這項移植實驗是在肢芽發育的早期進行的,所以通過顯微鏡肉眼可以觀察到,這只翅膀看上去仍是相互一致的間質細胞群。在奧斯特模型中,這樣的肢芽仍能生成正常的臂,因為這時的形態發生場還沒有對軟骨的形成產生作用。然而在那次實驗中,肢芽卻形成了兩根肱骨。該實驗結果使生物學家們得出這樣的結論,即在試驗移植之前,并早在奧斯特所說的作用于肢芽的力量產生影響之前,肢芽中必定已經存在了一個看不見的標識。
當時科學家們不可能考慮到還有其他的解釋,因為那時候的基因學家幾乎不明白基因是如何創建胚胎的。幾十年來,基因學家們在突變研究領域相互較勁。其中,一位名叫威廉·貝特森(William Bateson)的英國動物學家于19世紀90年代首先觀察到一組尤其引人注目的變異。一只葉蜂的腳長在了觸角上,一只螯蝦在眼睛部位長出了一根觸須,一只飛蛾原先的腿長成了翅膀。哺乳動物也有一些盡管更細微但與上述相似的突變,例如頸椎骨中長出了小肋骨。貝特森將它們稱為同種突變,因為突變的器官本身形態沒有發生改變,只是生長的位置發生了變化。
直到20世紀80年代,基因學家們才在一只發育中的果蠅幼蟲中將骨骼的同種隨機序列追溯至發生作用的同源異型基因。當他們將所有的基因一一鑒別后,再重新審視這一成果時,才驚訝地發現這個理論是如此的簡單。同源異型基因在果蠅的DNA上像一串燈籠順序串起,序列中的第一個基因開始活動,在幼蟲頭上的一些細胞里生成蛋白質,其余的基因按順序進行相同的活動,直至幼蟲尾部細胞內的蛋白質生成為止。幼蟲由體節構成,每節體節內重疊的同源異型基因組合方式不同,這些基因在細胞內生成蛋白質。在每個細胞內,這些蛋白質游到DNA的其他地方,開啟其他基因,而這些基因再生成每節體節所需的骨骼,如1條腿、1只翅膀或1只觸角之類。貝特森發現的同種突變是同源異型基因在錯誤的體節內的活動使該體節有了新的特征。
脊椎動物的發育模式與果蠅的發育模式相差很大,這一點并不難理解,因為脊椎動物與果蠅在10億年前才有共同的祖先。脊椎動物的胚胎在其背部也有體節,這些體節除生成皮膚、肌肉及神經外,還生成內部骨骼——脊椎及肋骨。然而,20世紀80年代后期,基因學家發現小雞胚胎及老鼠胚胎中的活動基因與昆蟲的同源異型基因幾乎相同。人的同源異型基因能夠植入果蠅的DNA中并能發揮與果蠅基因相同的作用。脊椎動物的同源異型基因也規定了胚胎從頭至尾的樞椎。而且如果基因學家在這些基因中創造突變,他們將得到同種突變。在胸腔后部,頸椎骨有了椎骨的形狀。哺乳動物胚胎中第二個鰓狀的袋狀結構通常生成一種細胞,這種細胞最終將形成鐙骨,即中耳內3個小骨中的一塊。但是如果同源異型基因不被認真對待,這種袋狀物將不會生成鐙骨,而是生成砧骨和錘骨——中耳的兩塊小骨,而這兩塊小骨通常是由第一個鰓狀的袋結構生成的。

幾乎完全相同的同源異型基因(用方格表示)既決定了昆蟲幼蟲體節的未來形態,也決定了四足動物胚胎的未來形態。圖中豎條顯示的是某種基因在細胞內對蛋白質生成的決定性的大小。
每一次變化都要求有更為復雜的協調發展,一整套復制出的同源基因,它們大量制造出超出身體發展所需的蛋白質,從而為新結構的形成提供條件。
幾年內,科學家們在蠕蟲、水蛭、海星及許多其他動物中發現了相同的基因。他們驚奇地發現有些動物的同源異型基因不止一組,而是有幾組,每組與其他組稍微有所不同。果蠅只有一組同源異型基因,但諸如七鰓鰻之類的無頜脊椎動物有3組,而多數有頜的魚及四足動物有4組或5組。絕大多數動物體內存在著同源異型基因的事實使同源異型基因在研究果蠅的實驗室內發揮的作用不再只是一種工具:同源異型基因是整個動物王國進化遺產的一部分,它們能告訴我們生物進化是如何發生的。所有帶有同源異型基因的動物必定是從共同的祖先進化而來的,并運用一組同源異型基因在從頭至尾的樞椎上形成體節。這有著10億年歷史的生物繁衍出子孫,進化成昆蟲及其他無脊椎動物,它們用遺傳密碼來控制幼蟲的生長。與之不同的是,海星利用同源異型基因來伸展出每只足的樞椎。脊椎動物在進化過程中對其自身的同源異型基因進行多次復制,一次是在第一批脊椎動物出現時,后來一次是在有頜脊椎動物出現時。
對整組基因的一次復制就是一次激進的突變,而在幾百萬年中,這種突變經常發生。起先,復制出的基因僅僅像原先的基因一樣產生蛋白質,因為對復制基因的改變不會影響原基因起的作用。然而持續不斷地誘發DNA的突變更容易改變復制基因的作用,多數情況下,這些副本只是讓DNA多出一些無用的基因,但有時復制出的基因能突變成一種新的形式,發揮新的作用。
例如,許多動物體內的免疫細胞用一種叫做溶菌酶的蛋白質來抵抗病菌。免疫細胞吞噬下微生物,釋放出溶菌酶,溶菌酶附在細菌表面鉆孔,讓分子出入細菌膜,直至細菌死去。像多數哺乳動物一樣,人體內的溶菌酶在血液外惟一能存在的地方就是眼淚,通過眼淚防止眼睛受到感染。但是反芻動物——如牛、山羊及綿羊之類進行反芻的有蹄類哺乳動物在胃中存在著大量的變異溶菌酶。反芻動物的祖先顯然是復制了生成溶菌酶的基因,而復制出的那套基因適應了胃中的強酸。反芻動物與存活在它們胃里的細菌有著共生關系,反芻動物會周期性地吞食一些微生物進入胃內,新形式的溶菌酶在胃里能鉆開微生物,釋放出反芻動物能消化的維生素及營養物。
溶菌酶基因的復制使反芻動物的進化有了巨大的不同,但相較之下同源異型基因的復制——這種復制對建立胚胎的身體發育起著關鍵性作用——是一個非常重要的宏觀進化事件。同源基因(即脊椎動物的同源異型基因)的復制似乎與頭——環繞內有兩個光敏感器官的神經細胞叢的骨質外殼——的出現,是在相同時間里發生的。第二次基因的復制發生于脊椎動物從鰓弓進化成頜的時期,這個時候脊椎動物從鰷魚大小的以垃圾為食的動物轉化成海洋食肉動物,并進而在一億年內轉化成能鑿穿一條船的巨大獵食動物。每一次變化都要求有更為復雜的協調發展,一整套復制出的同源基因,它們大量制造出超出身體發展所需的蛋白質,從而為新結構的形成提供條件。
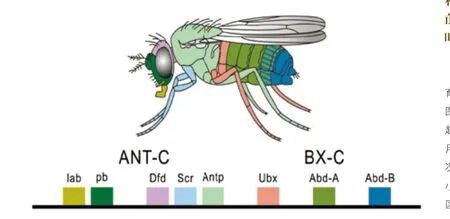
黑腹果蠅的八個同源異型基因,第一個基因影響嘴巴,第二個基因影響面部,第三個基因影響頭頂,如此類推,第八個基因影響下腹。基因與果蠅身體的顏色,表示基因所影響的身體部位。

基因的相互作用促使肢體的形成。本圖僅顯示了這些基因中的一部分。偏振行為區中的特殊的蛋白質促進頂外胚層脊產生生長因子,生長因子再刺激間質細胞進行復制。復制促使肢芽生長,間質細胞與肢芽又共同規定了同源基因在生長區的圖案。同源基因對決定肢體結構起著主要作用。
這些瑞士科學家們發現一些與形成人類胳膊的主要基因相同的基因,它們形成斑馬魚的前鰭,并且其生成模式在開始時也是完全相同的。
1989年,當生物學家們在繪制發育中的肢芽所生成蛋白質的相互作用圖時,他們開始意識到同源基因也在起作用。為了能理解這種相互之間的作用,你得先理解臂膀開始形成時的情況,即拇指指向頭而小指向下指向腳。小指側一帶的肢芽區叫做偏振行為區,這個區域分泌一種特殊的蛋白質。這種蛋白質從該區域分游至肢芽的脊部,在那兒它們使脊部細胞分泌出一種刺激生長的蛋白質,而這種蛋白質又游離至相鄰的間質細胞中使間質細胞急劇分裂。與此同時,刺激生長的蛋白質促進生成更多的這種特殊的蛋白質,這種蛋白質再游離回至肢芽的脊部。這樣急劇的分裂繁殖不斷地進行著,同時這兩種蛋白質也大量充溢在間質細胞間,并作為向同源基因傳遞信號的載體。
當科學家們第一次發現同源基因在肢體中發生作用時,他們推測同源基因在肢體內起的作用和它們在脊椎中的作用一樣:肢體將像一條切片面包般被分割成片,每片中的同源基因組合方式各異,將骨骼塑造成適當的形狀。按他們的設想,貫穿頭尾的樞椎變為貫穿肩膀與手。科學家們改變了老鼠及小雞體內的同源代碼,看這種變異能否證實他們的設想。早期的實驗似乎驗證了這種設想。例如,當基因學家們破壞一組同源基因中的最后一個基因后,新出生的小老鼠的尾巴縮短了,趾尖也萎縮了。
但更多的實驗顯示實際情況比設想更復雜:另一組同源基因僅沿著肢芽的小指側接通,然后突然轉向,越過指尖邊緣。困惑消失了。那些對肢體中同源基因迷惑不解的基因學家們對棘魚石螈一無所知,而科茨知道。他總是密切關注著能夠幫助他理解脊椎動物肢體進化的新科學發現,以及新發表的關于同源基因的論文。當他看到同源基因的巨大轉變時,他意識到之前已經看過這個模式:這個模式就是舒賓發現的指弓的曲線。“我看著它,”科茨說道,“并且想,這簡直太好了。”

不同區域的同源異型基因表現變化,會導致不同區域發展出不同的構造,如蝴蝶的前后翅膀。

孩子們與蝴蝶親密接觸。同源異型基因變化在蝴蝶前后翅膀上的體現。 攝影/Tolga Akmen/東方IC
科茨將這一意外發現展現給科學界,告訴他們化石、細胞及基因如何暗示著對這一宏觀進化的相同解釋。沒有人對肢體的了解能夠透徹到對同源基因在舒賓所說的“指弓中的作用”進行解釋,但無論這之中存在什么樣的機制,這種一致性都是如此清晰,不容忽略。受科茨的啟示,瑞士生物學家們在1995年論證了同源基因也會生成魚鰭。他們用來做實驗的是斑馬魚,這是一種能夠像老鼠一樣適應實驗室生存條件的長有鰭刺的小魚。斑馬魚的繁殖周期極快,能夠很快地生成魚苗,最為重要的是,在它生命的早期身體是透明的,這使得人們能夠很容易地看到它們體內對蛋白質敏感的染劑。這些瑞士科學家們發現一些與形成人類胳膊的主要基因相同的基因,它們形成斑馬魚的前鰭,并且其生成模式在開始時也是完全相同的。一簇間質細胞在斑馬魚的腹側凸起,然后脊在其外側隆起。在靠近魚尾部位的偏振行為區開始孕育特殊的蛋白質,脊與偏振行為區開始相互作用,結果,—組同源基因開始在尾部邊緣生成其蛋白質。然后,間質細胞開始聚集成簇。
如果斑馬魚是四足動物,那么離鰭較遠一側周圍的細胞將開始生成同源蛋白質,形成四足動物的弓形器官。但在斑馬魚體內,同源基因這時停止活動。間質細胞沿由同源基因標識的直直的樞椎進行聚合,然后分裂形成骨骼,這些骨骼都直接依附于肩部。但是,盡管同源基因停止活動,魚鰭卻繼續生長。在四足動物體內,一旦肢芽促使間質細胞進行增殖,肢芽的脊就開始消失;但在斑馬魚體內,這個脊自身也開始生長,伸展至一個扁平的大空間內,鰭刺在這里開始形成。

在斑馬魚體內,同源基因活動的那段鰭就是一根直樞椎,以后骨骼會沿著這根樞椎生成。在四足動物體內,相同的基因在肢芽的末端彎曲。在這個新的區域,基因圖案進行翻轉。這種基因圖案與之后的樞椎發展保持了一致。拇指側進行分叉,直至分叉延伸至腕處,在那里樞椎彎曲,分支繞到另一側,形成手指。
這些實驗結果與古生物學家的推想驚人地一致。輻鰭魚與肉鰭魚的共同祖先可能長有一個不大不小的鰭。它的同源基因形成供骨骼生長的樞椎,這條樞椎分叉成一些小的分支,脊在這些分支上形成一個小鰭。它的后代們形成兩大分支,沿著這兩個分支的發育時快時慢,在輻鰭魚這一分支內,例如斑馬魚,同源基因停止活動的時間較早,但脊內基因活動的時間也較早,從而形成較大的鰭刺。相反,肉鰭魚的同源基因起作用的時間較長,生成的樞椎相應長些,骨骼也相應大些。同時,同源基因延緩了鰭刺的生長,以致鰭刺成為環繞在內部骨骼周圍的一道須邊。而且肉鰭魚向四足動物的轉變發生的時間也要更早些:肢芽的脊很早就停止生長,所以不會長出任何鰭,但樞椎向肢芽的另一側彎曲,從而導致了手指的生長。
龍鰭魚顯然能用結實的胸肌與鰭處類似三頭肌的肌肉來支撐身體的前部。它有著與我們人類的肱骨、橈骨與尺骨相對應的巨大骨骼,除了這些,它還至少有8節類似我們手骨但更細長一些的骨骼。
1996年,一隊美國基因學家以一份對發育中四足動物肢體的詳細描述回應瑞士科學家們的發現。在對23個同源基因進行23天跟蹤研究之后,科學家們發現他們的研究成果與舒賓和阿爾伯齊所說的弓形器官有著難以置信的吻合。在肢芽生長的早期,同源基因所活動的區域產生條紋,形成小趾的邊緣。在肢芽發育得更大一些的時候,這些條紋繞過趾形成部位的另一側——但是這時條紋顛倒了次序,當舒賓及阿爾伯齊在將肉鰭轉變成四足動物的肢體時——那時還沒有人知道同源基因與四足動物肢體有著一定的關系——他們已經意識到了這種翻轉顛倒。在真掌鰭魚這種動物的體內,所有的骨骼分支都是從樞椎分叉出來向大趾方向延伸。當四足動物進化鉤狀器官時,這些分支轉向樞椎的另一邊,并在此處由樞椎生成趾。現在看來,如若要分支翻轉,在動物的進化中必須使同源基因翻轉。
舒賓花了幾年的時間來思考并認識到他在彎曲的樞椎上所做的研究在他畢生事業中是微不足道的,但現在基因學家們卻都要拉著他合寫關于肢體發育的論文,因為他們能夠辨識基因序列和染色體細胞,而他能夠譯注化石的密碼,這種密碼信息是任何當今動物都不能夠提供的。例如,近來,他和特德·戴施勒(Ted Daeschler)一直在研究一塊戴施勒于1996年發現的可能是長有趾的鰭的化石。這塊化石是泥盆紀一種叫做龍鰭魚(Sauripterus)的肉鰭魚的化石。它是在卡茨基爾地層發現的第一塊肉鰭化石,但是在戴施勒勘探之前,那里最好的化石已經被嚴重損壞。據古生物學家所知,龍鰭魚屬于一種已經滅絕的肉鰭魚,這種肉鰭魚與真掌鰭魚及四足動物的關系比其與肺魚的關系更緊密,但絕不是它們導致了陸地生物的出現。
戴施勒發現的化石保留了從肩至鰭刺尖端的鰭。你能摸到肌肉與骨骼相連的光滑凹處,你能看到能夠彎曲的圓形關節。龍鰭魚顯然能用結實的胸肌與鰭處類似三頭肌的肌肉來支撐身體的前部。它有著與我們人類的肱骨、橈骨與尺骨相對應的巨大骨骼,除了這些,它還至少有8節類似我們手骨但更細長一些的骨骼。盡管這些手指狀的骨骼仍長在鰭內,龍鰭魚能彎曲鰭上相當于腕部的位置,也能彎曲鰭上相當于指關節的位置。
現在,科學家們在觀察這樣的化石后能夠識別其中起作用的已消失的基因。與舒賓合作的哈佛大學醫學院的同源基因學家克里佛德·泰賓(Clifford Tabin)就已經指出,像真掌鰭魚及其他肉鰭魚一樣,龍鰭魚在靠近肩的部位,骨骼從樞椎分叉出來,并向鰭上長有大趾的方向擴展。但在鰭的末端,即趾狀骨骼形成的部位,一些分支轉變方向,不再向大趾方向擴展,而是向小趾側延伸。這絕不是四足動物的肢體——龍鰭魚肢體的樞椎內絕不會有鉤狀器官。然而,這塊化石卻顯示這種肉鰭嘗試著與我們人類祖先相同的基因處方。四足動物在大規模的手狀結構進化實驗中只是一部分,但它們碰巧是泥盆紀后惟一的幸存者。
