雙功能融合蛋白RDP-BDNF對東莨菪堿所致認知功能障礙小鼠學習記憶的影響
2014-05-17 02:28:05張苗苗張恩齊高飛燕項松濤付愛玲
中國藥理學通報 2014年11期
張苗苗,張恩齊,高飛燕,吳 靜,項松濤,付愛玲
(西南大學藥學院,重慶 400716)
腦源性神經營養因子(brain-derived neurotrophin factor,BDNF)是體內含量最高的神經營養因子,具有維持多種神經元存活并直接促進軸突生長的作用,能夠保護膽堿能神經元并與學習記憶密切相關[1]。然而由于血腦屏障(blood-brain barrier,BBB)的存在,限制了外源性BDNF等具有治療潛力的大分子化合物入腦并發揮作用。中樞神經系統給藥方法包括侵入性給藥方法和非侵入性給藥方法,前者藥物注射入腦后很快被組織清除,且不易擴散、對技術要求高、風險大,所以不能在臨床使用[2-3];后者則主要是通過載體的協助作用,將大分子物質轉運入腦。狂犬病毒糖蛋白(rabies virus glycoprotein,RVG)具有嗜神經性,能夠攜帶其他物質向中樞神經系統轉運[4]。序列分析顯示,RVG上的330-357氨基酸序列是病毒與神經元結合的關鍵部位[5],我們將這段序列與含9個精氨酸的膜轉導肽(cell penetrating peptides,CPPs)連接后形成 RVG衍生肽(RVG-derived peptide,RDP)。有文獻報道,RDP能夠攜帶多種生物大分子通過血腦屏障并發揮其生物學功能[6-8]。本實驗試圖通過將 RDP肽段與BDNF蛋白融合,檢測其攜帶BDNF蛋白的入腦情況,探究RDP-BDNF對認知障礙模型小鼠的作用。
1 材料與方法
生物工程研究所;Ach ELISA檢測試劑盒購自上海源葉生物科技有限公司;石杉堿甲、氫溴酸東莨菪堿,購自Aladdin公司;其它均為國產分析純試劑。
1.2 pET28a-RDP-BDNF,p ET28a-BDNF載體構建及表達 為了盡量RDP與BDNF兩個肽段的空間結構,在BDNF序列的上游引入Linker,將RDP基因與BDNF基因序列連接,并構建pET28a-BDNF載體作為對照。設計兩對引物,導入HindⅢ和XhoⅠ酶切位點。pET28a-RDP-BDNF上游引物:5′-CAAAGCTTGGTGGAGGCGGTTCAGGCGGAGGTGGC TCAATGCACTCTGACCCTGCCCG-3′,下游引物:5′-GACTCGAG CTATCTTCCCCTTTTAATGG-3′;pET28a-BDNF上游引物:5′-CAAAGCTTCAATGCACTCTGACCCTGCCCG-3′,下游引物:5′-GACTCGAGCTATCTTCCCCTTTTAATGG-3′。
以pGEM-T-rhBDNF載體為模板,PCR擴增:反應條件:熱啟動5 min后加入pfu酶;94℃變性40 s,50℃退火40 s,72℃延伸3 min,40個循環;72℃保溫10 min,使反應物擴增充分;4℃保存。所得產物膠回收后酶切,連接,轉化至BL21中,挑取陽性菌落,提取質粒酶切鑒定并測序。
陽性克隆接種至LB培養基(含50 mg·L-1卡那霉素),37℃,160 r·min-1培養至 OD600約為0.6~0.8,加入終濃度 1.0 mmol·L-1的 IPTG,20℃,180 r·min-1,6 h誘導表達。誘導結束后高速離心收集菌體,用MCAC-0緩沖液(20 mmol·L-1Tris-HCl,500 mmol·L-1NaCl,質量分數 10%甘油,pH 8.0)洗滌后重懸浮。-20℃凍融,超聲破碎后高速離心,取上清,經Ni+親和層析柱純化,冷凍干燥后得蛋白純品,SDS-PAGE電泳。
1.3 RDP-BDNF在腦和血清中的含量測定 將小鼠分為3組,分別經尾靜脈注射RDP-BDNF、BDNF蛋白(2 mg·kg-1),陰性對照注射等量的生理鹽水,于 0.25、0.5、1、3、5 h取血清、大腦,-80℃保存備用。BDNF在腦和血清中的含量用BDNF ELISA試劑盒檢測。
1.4 AD模型鼠的制備及RDP-BDNF治療 將小鼠隨機分為:正常對照組、東莨菪堿模型組、石杉堿甲治療組(0.1 mg·kg-1)、RDP-BDNF治療組(2 mg·kg-1)、BDNF治療組(2 mg·kg-1),共 5組。實驗前連續腹腔給藥10 d,正常對照及模型組分別給予等量的生理鹽水。于11 d開始造模并進行Morris水迷宮(成都泰盟科技有限公司)測試。
于每天實驗前,除正常對照組腹腔注射等量的生理鹽水外,其余各組小鼠均予腹腔注射東莨菪堿2 mg·kg-1,直到實驗全部結束。
參照Morris[9]方法進行實驗。各組于造模后20 min進行水迷宮測試。定位航行實驗時每只小鼠每天訓練1次,每次按東、西、南和北4個入水點分別放入水池。每只小鼠有120 s找到平臺,120 s未找到平臺者,將其引至平臺,放置30 s,使其記憶平臺的位置;數據采集和處理均由圖像自動監視和處理系統完成。整個實驗需要6 d完成,每只小鼠平均游泳時間和平均游泳速度作為衡量其學習記憶能力的指標。定位航行結束后,移去平臺,將小鼠從某一點放入,每只小鼠游泳時間為90 s,記錄其游泳軌跡及在目的象限游泳的時間。
1.5 生化指標測定 水迷宮實驗結束后,將小鼠處死,快速取腦,-80℃保存備用。分別按照各試劑盒說明書上的方法測定各組小鼠腦中BDNF、乙酰膽堿(acetylcholine,ACh)、谷胱甘肽(glutathione,GSH)的含量及膽堿乙酰化轉移酶(choline acetyltransferase,ChAT)和超氧化物歧化酶(superoxide dismutase,SOD)的酶活性。
1.6 統計學處理 使用SPSS 13.0軟件進行數據分析,實驗數據以ˉx±s表示,數據采用單因素方差分析,組間比較采用q檢驗。
2 結果
2.1 融合蛋白的表達 以pGEM-T-rhBDNF載體作為模板,擴增出360 bp左右條帶,經測序證實為BDNF基因,采用基因工程技術構建pET28a-RDPBDNF、pET28a-BDNF載體,測序驗證成功后,IPTG誘導表達。樣品處理后SDS-PAGE電泳分析,得到一條17 ku左右的特異性條帶,與融合蛋白的理論大小一致,說明我們成功的得到了融合蛋白(見Fig 1)。

Fig 1 SDS-PAGE for RDP-BDNF fusion proteinM:Protein molecular weight markers;1:Null plasmid BL21 bacteria control;2:Uninduced pRDP-BDNF BL21;3:Induced pRDP-BDNF BL21;4:Purified RDP-BDNF fusion protein;5:RDP-BDNF in supernatant after ultrasonic disruption;6:Fusion protein in precipitation after ultrasonic disruption of bacteria.
2.2 RDP-BDNF在小鼠大腦和血清中的時效曲線由Fig 2A可知,尾靜脈注射RDP-BDNF蛋白15 min后,即在小鼠腦中檢測到外源性BDNF蛋白,蛋白的含量逐漸降低,在5 h時基本恢復到正常值;然而注射BDNF蛋白的小鼠腦中BDNF含量沒有明顯變化,說明RDP-BDNF可以穿過血腦屏障。Fig 2B則顯示,注射RDP-BDNF和BDNF蛋白后,兩組小鼠血清中BDNF含量都明顯提高,BDNF組上升幅度更大。

Fig 2 Time-course relationship of RDP-BDNF delivery in cerebrum(A)and serum(B)

Fig 3 Results of probe testA:The time spent in target quadrant in probe test.**P<0.01 vs control;#P<0.05,##P<0.01 vs model;B:The representative swimming route of animals in probe test.
2.3 RDP-BDNF融合蛋白對東莨菪堿所致認知障礙小鼠學習記憶的影響 由Tab 1可知,在定位航行實驗中,模型小鼠找到平臺的潛伏期比對照組長,差異有統計學意義,說明認知障礙模型制備成功;石杉堿甲治療組和RDP-BDNF治療組的潛伏期比模型組短,且差異有統計學意義,說明融合蛋白對模型鼠的認知功能有明顯改善作用;Tab 2中,各組小鼠之間的游泳速度沒有差異,說明RDP-BDNF改善了小鼠的學習記憶能力,而不是其運動功能。由Fig 3可得,在空間探索實驗中,移去平臺后,石杉堿甲治療組和RDP-BDNF治療組在目的象限的停留時間比模型組明顯延長,進一步說明融合蛋白治療能增強模型小鼠的記憶能力。
2.4 RDP-BDNF的作用機制 為了探究RDP-BDNF的作用機制,我們測定了相關物質的指標。由Fig 4可知,與對照組相比,模型組小鼠腦中BDNF含量下降,RDP-BDNF治療組模型小鼠腦中的BDNF蛋白含量則明顯增高,說明RDP-BDNF可以穿過血腦屏障,改善模型組小鼠腦內BDNF的不足;Fig 5顯示,與對照組相比,模型小鼠腦內ChAT酶活性和ACh水平都明顯降低,而RDP-BDNF和陽性對照藥石杉堿甲治療都可以提高模型小鼠腦中二者的水平,說明補充模型小鼠腦內BDNF含量,可以增強膽堿能神經功能;Fig 6則表明,RDP-BDNF治療明顯提高模型小鼠腦中SOD酶活性和GSH水平,由此可知,提高腦內BDNF含量可以維持氧化應激平衡,保護神經元免受自由基損害。

Tab 1 Time of escape latency in Morris water maze

Tab 2 Swimming speed(cm/s)of mice in Morris water maze

Fig 4 Effect of exogenous RDP-BDNF on BDNF level in cerebra**P<0.01 vs control;##P<0.01 vs model.
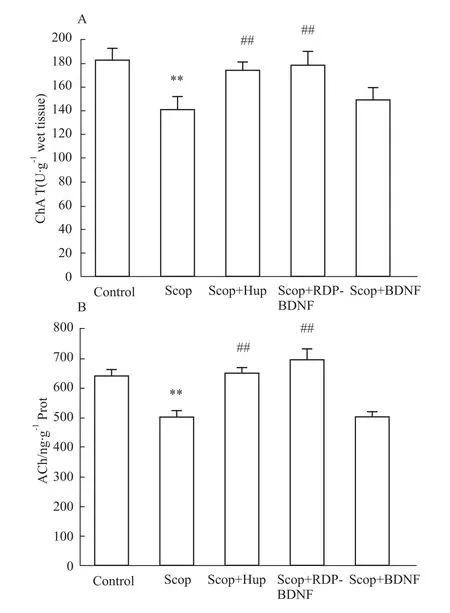
Fig 5 Influence of RDP-BDNF on activity of ChAT and level of ACh of model mice**P<0.01 vs control;##P<0.01 vs model.
3 討論
BDNF幾乎在神經回路發育的各個環節都發揮至關重要的作用,其可與酪氨酸激酶受體(trkB)結合,激活細胞分裂素活化蛋白激酶(MAPK)和磷脂酰肌醇(-3)激酶(PI3K)通路[10],同時影響下游的轉錄因子的活性[11]。研究表明,在癡呆病人的海馬、顳葉及其他皮質區BDNF的水平都有降低,而給予外源性BDNF蛋白可以減緩膽堿能神經元的退化,提高其功能[12-14]。然而,BBB卻限制了外源性BDNF蛋白活性的發揮。因此,本研究提出將新型穿膜肽RDP與BDNF連接制備融合蛋白,并通過ELISA方法檢測融合蛋白的活性及其在腦和血清中分布的時效曲線;在行為學測試中,RDP-BDNF融合蛋白治療明顯改善認知障礙小鼠的學習記憶能力。以上結果表明,RDP可以攜帶BDNF進行入腦轉運,并且不影響其生物活性。
關于RDP-BDNF治療認知障礙小鼠的機制,我們基于兩個假說進行研究。首先,根據膽堿能假說可知,膽堿能神經元的損傷對小鼠的認知功能有影響,因此,我們檢測了小鼠大腦中ChAT的酶活性及ACh的水平。結果顯示,RDP-BDNF治療組小鼠腦中ChAT酶活性和ACh水平與模型組相比都有明顯提高。其次,根據氧化應激假說,癡呆病人腦中自由基氧化會導致蛋白質氧化、脂質超氧化及線粒體氧化損傷。為了研究RDP-BDNF治療與氧化應激系統的關系,我們檢測了各組小鼠腦中SOD的酶活性及GSH的水平。結果顯示,RDP-BDNF治療組的SOD酶活性和GSH水平與模型組相比都有明顯提高。所以,RDP-BDNF對東莨菪堿誘導的認知障礙小鼠的治療可能與增強膽堿能神經功能及抑制自由基氧化,維持氧化應激狀態平衡有關。
目前,RDP-BDNF穿過BBB進入腦組織的機制尚不清楚,我們實驗室之前的研究表明,γ-氨基丁酸(GABA)和氯丙嗪可以阻斷細胞對RDP的特異性攝取,這說明RDP可能通過與GABA受體結合,并經網格蛋白調節的細胞內吞途徑進入細胞[7]。由此,我們推測,RDP-BDNF穿過血腦屏障的機制可能與受體介導有關,有關這方面的研究仍在繼續進行。
盡管近年來科學家們一直在尋找能有效將有活性的生物大分子轉運入腦的方法,BBB仍然是限制這些活性因子發揮作用的主要障礙。本研究結果表明,RDP可以攜帶BDNF進入腦內,并且不影響其生物活性,因此,我們推測由RDP攜帶有治療活性的蛋白質入腦并發揮作用將為腦部疾病的治療提供一種新方法。

Fig 6 Effects of RDP-BDNF on activity of SOD and level of GSH of model mice*P<0.05,**P<0.01 vs control;#P<0.05,##P<0.01 vs model.
參考文獻:
[1] Lu B,Nagappan G,Guan X,et al.BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases[J].Nat Rev Neurosci,2013,14(6):401-16.
[2] Pardridge W M,Kang Y S,Buciak J L.Transport of human recombinant brain-derived neurotrophic factor(BDNF)through the rat blood-brain barrier in vivo using vector-mediated peptide drug delivery[J].Pharm Res,1994,11(5):738-46.
[3] Pardridge W M.Blood-brain barrier drug targeting:the future of brain drug development[J].Mol Interv,2003,3(2):90-105.
[4] Tuffereau C,Benejean J,Alfonso A R,et al.Neuronal cell surface molecules mediate specific binding to rabies virus glycoprotein expressed by a recombinant baculovirus on the surfaces of lepidopteran cells[J].J Virol,1998,72(2):1085-91.
[5] Tang Q,Orciari L A,Rupprechti CE,et al.Sequencing and position analysis of the glycoprotein gene of four Chinese rabies viruses[J].Virol Sin,2000,15(1):22-33.
[6] Fu A L,Wang Y L,Zhan L P,et al.Targeted delivery of proteins into the central nervous system mediated by rabies virus glycoprotein-derived peptide[J].Pharm Res,2012,29(6):1562-9.
[7] Fu A L,Zhao ZZ,Gao FY,et al.Cellular uptake mechanism and therapeutic utility of a novel peptide in targeted-delivery of proteins into neuronal cells[J].Pharm Res,2013,30(8):2108-17.
[8] 高飛燕,張苗苗,徐興然,等.穿膜肽引導核酸靶向性進入神經細胞的研究[J].中國藥理學通報,2014,30(3):326-30.
[8] Gao F Y,Zhang M M,Xu X R,et al.The study of a cell-penetrating peptide mediated targeted-delivery of DNA into neuronal cell[J].Chin Pharmacol Bull,2014,30(3):326-30.
[9] Morris R.Development of a water maze procedure for studying spatial learning in the rat[J].J Neurosci Methods,1984,11(1):47-60.
[10]Barnabe-Heider F,Miller F D.Endogenously produced neurotrophins regulate survival and differentiation of cortical progenitors via distinct signaling pathways[J].J Neurosci,2003,23(12):5149-60.
[11]Ito H,Nakajima A,Nomoto H,et al.Neurotrophins facilitate neuronal differentiation of cultured neural stem cells via induction of mRNA expression of basic helix-loop-helix transcription factors Mash1 and Math1[J].J Neurosci Res,2003,71(5):648-58.
[12]Lee J,Fukumoto H,Orne J,et al.Decreased levels of BDNFprotein in Alzheimer temporal cortex are independent of BDNF polymorphisms[J].Exp Neurol,2005,194(1):91-6.
[13]Peng S,Wuu J,Mufson E J,Fahnestock M.Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer′s disease[J].J Neurochem,2005,93(6):1412-21.
[14]Morse JK,Wiegand SJ,Anderson K,et al.Brain-derived neurotrophic factor(BDNF)prevents the degeneration of medial septal cholinergic neurons following fimbria transection[J].J Neurosci,1993,13(10):4146-56.
[15]Dunbar G L,Rylett R J,Schmidt B M,et al.Hippocampal choline acetyltransferase activity correlates with spatial learning in aged rats[J].Brain Res,1993,604(1-2):266-72.
[16]Beal MF.Aging,energy,and oxidative stress in neurodegenerative diseases[J].Ann Neurol,1995,38(3):357-66.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
學苑創造·A版(2020年9期)2020-10-13 09:41:02
現代出版(2020年3期)2020-06-20 07:10:34
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
