新疆莎車縣5個扁桃品種抗寒性比較研究
2014-06-21 12:42:48王夢雪李志軍
塔里木大學學報 2014年1期
關鍵詞:植物
王 琳 王夢雪 李志軍
扁桃(Amygdalus communis L.)屬薔薇科李亞科桃屬扁桃亞屬植物,為世界著名的干果樹種,原產于中亞細亞和非洲北部山區,新疆也是扁桃的原產地之一[1]。其適應性強,耐旱、耐貧瘠、耐熱、耐鹽堿,特別適合在新疆發展,是南疆干旱荒漠及綠洲地區生態建設的優良樹種。扁桃品種極其豐富,據報道,全世界有扁桃品種4 000余個,我國自行培育和引進的品種有200多個,其中新疆約有90多個[2]。主要集中在南疆的喀什、疏勒、疏附、英吉沙、葉城、澤普、和田等地,栽培面積約為10 000 hm2[3]。雖然品種資源豐富,但是由于冬季不能順利越冬等原因影響其經濟效益。因此,選育抗寒扁桃品種是促進扁桃產業化發展的必由之路,而研究扁桃品種的抗寒能力對抗寒品種的選育以及在我國各地的推廣具有重要的意義。目前對新疆莎車縣主栽扁桃品種抗寒性的研究尚未見報道。本研究以新疆莎車縣5個主栽扁桃品種為研究對象,分析與抗寒性相關生理生化指標(相對電導率、MDA含量、Pro含量、SOD活性、可溶性蛋白及可溶性糖)的變化規律,初步評價5個扁桃品種在栽培地抗寒性的強弱,為抗寒性品種選育和栽培推廣提供理論依據。
1 材料與方法
1.1 材料
供試材料為紙皮1號、阿曼尼莎11號、葉爾羌14號、晚豐18號、選育品種5號,5個扁桃品種一年生枝條于2013年1月取自新疆莎車縣扁桃生產基地。
1.2 儀器
DDS-11C電導儀、高速臺式冷凍離心機(HERMLE Z323K)、電子天平(梅特勒 -特利多AL104)、WD4005低溫試驗箱、755B單光束紫外可見分光光度計等。
1.3 試驗方法
1.3.1 材料處理 將采集的供試扁桃1年生枝條剪成30~40 cm長的枝段,石蠟封閉兩端剪口,先后用自來水和去離子水洗滌晾干后,用塑料袋分裝好待處理。處理溫度為-15℃、-20℃、-25℃、-30℃、-35℃、-40℃。每處理設3個重復,每個處理12個枝條。冷凍處理時,將分裝好的枝條放人低溫冰箱,以3℃/h-1速度降溫,降到所設溫度后維持24 h,備測。
1.3.2 萌芽率統計 將不同低溫處理后的枝條于室溫條件下進行水培,4周后統計萌芽率,每個處理重復3次,每個處理12個枝條。萌芽率=(萌發芽個數/水培枝條芽的總數)×100%。
1.3.3 生理指標的測定
1.3.3.1 電解質相對外滲率的測定[4]
將低溫冷凍處理后的枝條切片,然后稱取1g試樣投入三角瓶中,加入50 ml的蒸餾水,浸泡24 h,即可以測定出浸出液的電導率值。測試材料需設重復三次,然后將低溫處理后測定的各樣品,放在水浴鍋中煮沸1 h,殺死組織,再加蒸餾水補充到原來溶液的定量,靜止冷卻1 h后測定其電導率,用百分比法計算供試樣品的電解質的相對外滲率。
電解質的相對外滲率(傷害度)=處理電導率值/處理煮沸后電導率值×100%
1.3.3.2 可溶性蛋白質含量的測定[5]:采用考馬斯亮藍G-250染色法。
1.3.3.3 丙二醛(MDA)含量的測定[4]:硫代巴比妥酸(TBA)法測定。
正常情況下由于植物體內活性氧的產生與清除處于平衡狀態,不會導致植物傷害。但在低溫脅迫環境下,植物器官往往發生膜脂過氧化作用。
1.3.3.4 脯氨酸含量的測定[6]:茚三酮法測定。
1.3.3.5 保護酶活性變化[6]:超氧化物歧化酶 S0D用愈創木酚法測定。
1.3.3.6 可溶性糖含量的測定[5]:蒽酮比色法。
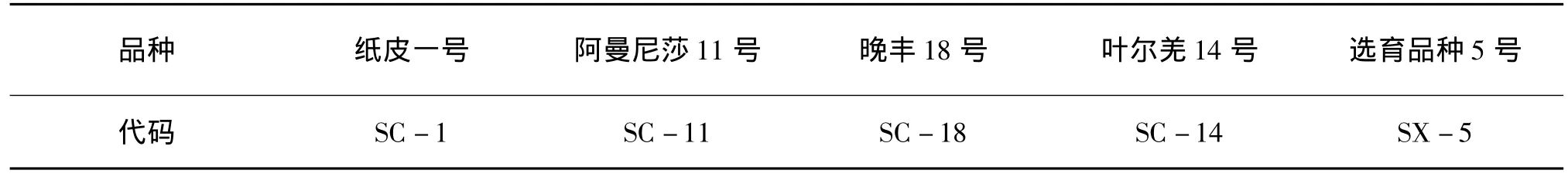
表1 扁桃不同品種代碼
1.4 數據統計與分析
數據統計及圖表制作用 Microsoft Excel,結果分析用DPS統計軟件處理。
2 結果與分析
2.1 低溫處理對不同品種扁桃枝條恢復生長的影響
樹木受凍害后,枝條恢復生長是評價其受凍害程度最直觀方法,能夠客觀反映品種的實際抗寒能力。該方法已在仁用杏[7]、野生扁桃[8]等植物的抗寒性研究中得到證實。不同扁桃品種經不同低溫處理后,其萌芽率有明顯的差異(見表2、圖1)。隨著處理溫度的降低,枝條萌芽率隨之降低,而抗寒性較強的品種,其枝條的恢復生長能力較強,從表2可以看出,當溫度達-35℃時,5種扁桃供試材料中,SC-11枝條萌芽最高為13%,其次是SC-14枝條萌芽為2%。

表2 不同低溫條件下不同品種扁桃枝條的萌芽率

圖1 低溫處理對扁桃品種枝條恢復生長情況
2.2 低溫處理對扁桃枝條電解質外滲率的影響
細胞膜的破壞程度可以通過細胞內電解質外滲率的的大小來反映,抗寒性強的品種,其電解質滲出率相對較低。由圖2可以看出,不同扁桃品種的枝條經低溫處理后,其電解質外滲率隨處理溫度的降低總體呈上升趨勢。說明隨著溫度的降低對細胞膜的傷害逐漸增大,細胞內可溶性物質大量外滲膜透性增加。隨著溫度的降低,5個扁桃品種休眠枝的解質外滲率變化均為從-15℃降至-30℃增加較緩慢,溫度降至-35℃時達到高峰,此時SC-1和SC-18電解質滲出率達到了61.75%和60.56%。在整個過程中,SC-11的電解質滲出率均為最小,SC-14其次,表明這兩品種抗寒性相對較強。綜上可推測5個扁桃品種抗寒性表現為 SC-11>SC-14>SX-5>SC-18>SC-1,這與枝條萌芽率結果基本相符。

圖2 低溫對不同品種扁桃枝條相對電導率的影響
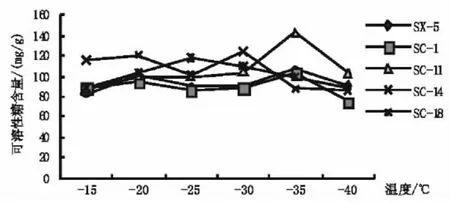
圖3 溫度處理過程中不同品種扁桃枝條可溶性糖的變化
2.3 低溫處理對扁桃可溶性糖含量的影響
研究表明,可溶性糖含量與植物抗寒性間存在一定的相關性[9,10]。隨著低溫強度的增加,植物細胞內大分子物質可通過水解轉化為糖,糖可通過提高細胞的滲透濃度。同時,糖還對原生質體、線粒體及膜上敏感偶聯因子均有保護作用,從而在一定程度上提高了植物的抗寒性[7]。從圖3可以看出,不同低溫處理對不同扁桃品種休眠枝可溶性糖含量的影響不同。隨著處理溫度的降低,5個扁桃品種枝條中可溶性糖含量的變化總體呈先升高后下降的趨勢,但不同品種達到最大值的溫度不同,-25℃時,SC-18可溶性糖含量達到最大值,為 118.03 mg/g;-30℃時,SC-14可溶性糖含量達到最大值,為124.48 mg/g;-35℃時SC-11可溶性糖含量達到最大值,為142.23 mg/g,表明抗寒性弱的品種可溶性糖含量峰值出現早于抗寒性強的品種,而整個過程中SC-1、SX-5可溶性糖含量均保持相對較低的水平,說明其抗寒性相對較差。
2.4 低溫處理對扁桃枝條脯氨酸含量的影響
脯氨酸是植物在逆境協迫下的一種重要的滲透調節物質。研究表明,脯氨酸含量高的品種抗寒性較強,反之較弱[11]。從圖4中可以看出,在休眠枝條中隨處理溫度的降低脯氨酸含量呈先上升后下降的趨勢,在整個過程中SC-14及SC-11兩個品種中脯氨酸含量相對最高,分別從最初的839.74 μg/g和671,47 μg/g至最大值 2 418.27 μg/g和2 370.19μg/g,分別比最初增加了2.88倍和3.53倍,說明其枝條對低溫的適應較好。而SC-1增幅最小,由最初的615.38μg/g增至1 280.45μg/g,比最初增加了2.08倍,并且在這個過程中脯氨酸含量相對較低。

圖4 不同溫度對不同品種扁桃枝條脯胺酸含量的影響
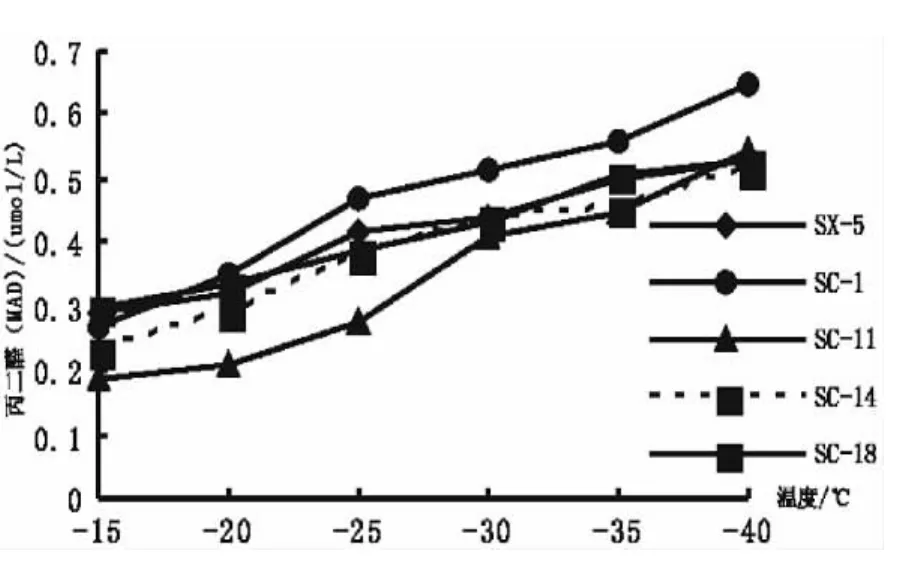
圖5 不同溫度處理對不同品種扁桃枝條兩二醛含量的影響
2.5 低溫處理對扁桃枝條丙二醛(MDA)含量的影響
從圖5可知,在降溫過程中供試的5個扁桃品種休眠枝的丙二醛(MDA)含量的變化趨勢與枝條的相對電導率的變化趨勢基本相似,隨處理溫度的降低總體呈上升趨勢。在整個低溫處理過程中SC-11、SC-14的MDA含量相對最小,當溫度為-35℃時 MDA含量分別為0.443 3 mmol/L和0.455 9mmol/L,而SC-1在整個低溫處理過程中都保持相對較高的MDA含量,當溫度為-35℃時MDA含量為0.561 2 mmol/L。當植物受到凍害后引發和加劇了膜脂過氧化作用,造成膜脂過氧化產物MDA增多,而其在組織中含量的多少反映了植物細胞受傷害的程度。MDA含量高說明植物的抗寒性弱,反之,則抗寒性強。因此,由圖5可推知整個過程中供試扁桃的抗寒表現為SC-11>SC-14>SC-18> SX-5>SC-1。
2.6 低溫處理對扁桃枝條SOD含量的的影響
SOD酶是清除超氧陰離子自由基的重要酶類,可有效清除因環境脅迫而累積的生物自由基,在保護生物免遭逆境傷害方面具有重要作用,SOD酶活性的大小和超氧陰離子自由基產生速率都可以表示植物抗寒過程中的生理狀況[12,13]。SOD 酶活性強的品種抗寒性較強[14,15]。從圖6可以看出,從總體變化趨勢來看,5個扁桃品種的SOD酶活性隨處理溫度的降低明顯呈先升后降的趨勢,在-30℃時達到最大。而SC-11及SC-14在整個低溫過程中SOD酶都保持相對較高的活性,SX-5品種的SOD酶活性最低。

圖6 低溫處理過程中扁桃枝條SOD酶活性的變化
圖7 低溫處理過程中不同品種扁桃枝條可溶性蛋白含量的變化
2.7 低溫處理對扁桃可溶性蛋白含量的影響
可溶性蛋白作為滲透調節物質、能量物質和信息傳遞物質在植物的抵御逆境的過程中起到較大的作用[13]。有研究表明,遭受低溫脅迫的植物,體內可溶性蛋白含量會增加,且細胞內可溶性蛋白和抗寒凍性之間表現出明顯的相關性[16,17]。從圖7可以看出,隨著處理溫度的降低,5個扁桃品種可溶性蛋白含量都有一個先上升后下降的變化趨勢,其中,SC-1、SC-14、SX-5在-30℃時可溶性蛋白含量達到最大,分別為 3.04 mg/g、3.01 mg/g、2.81 mg/g,而SC-11、SC-18在-35℃時可溶性蛋白含量達到最大,分別為3.69 mg/g、3.29 mg/g,表明抗寒性弱的品種可溶性蛋白含量峰值的出現早于抗寒性強的品種。
3 結論
本研究通過對低溫處理后休眠枝條的萌芽率及相對電導率、丙二醛(MDA)含量、脯氨酸(Pro)含量、可溶性蛋白含量、可溶性糖含量、超氧化物歧化酶(SOD)的活性7個生理生化指標的綜合分析,得出5個供試扁桃品種的抗寒性順序為SC-11>SC-14>SC-18>SX-5>SC-1。
在關于果樹抗寒性的研究過程中,細胞膜膜透性、滲透調節物質、抗氧化酶都被用作抗寒性的鑒定指標[18,19,20],然而植物的抗寒性受多種因素影響,孤立地用單一的生理指標很難反映植物的抗寒的實質,也不利于揭示植物抗寒的本質。而在本研究中,5個扁桃品種依據不同測定指標所得抗寒性排序結果也存在著一定的差異,因此,為了服單一指標對扁桃抗寒性鑒定的不足,必須通過綜合評價方法對本試驗測定的生理生化指標進行綜合分析,進一步確定了各測定指標對抗寒性的貢獻,并探討抗寒性鑒定的方法和模型,以期獲得新疆莎車縣扁桃品種抗寒性鑒定的有效方法,為該地區主栽扁桃種質資源評價和扁桃品種的引種提供依據。
[1] Kester D E,Gradziel T M.Almonds[R].Fruit Breeding,Volume Ⅲ,1996.
[2] 成建紅,侯平,李疆,等.巴旦杏的產業發展及其研究進展[J].干旱區研究,2000,17(1):32 -38.
[3] 李疆,胡芳名,李文勝,等.扁桃的栽培及研究概況[J].果樹學報,2002,19(5):346 -350.
[4] 王學奎.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[5] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[6] 高俊風.植物生理學實驗技術[M].世界圖書出版公司,2000.
[7] 鄭元,楊途熙,魏安智,等.低溫脅迫對仁用杏幾個抗寒生理指標的影響.西北農林科技大學學報(自然科學版,2008,36(1):163 -167.
[8] 魏鈺,郭春會,張國慶,等.我國幾個扁桃種抗寒性的研究[J].西北農林科技大學學報[自然科學版],2012,40(6):99-105.
[9] 王飛,陳登文,王卿,等.杏品種的需寒量與抗寒性的相關研究[J].中國農業科學,2001,34(5):465 -468.
[10] 張婷,車鳳斌,潘儼,等.低溫脅迫對核桃枝條幾個抗寒生理指標的影響[J].新疆農業科學,2011,48(8):1428-1433.
[11] Arbaoui M,Balko C,Link W.Study of faba bean(Vicia faba L.)winter 2 hardiness and development of screening methods[J].field Crops Research,2008,106:60 -67.
[12] 楊向娜,魏安智,楊途熙.仁用杏3個生理指標與抗寒性的關系研究[J].西北林學院學報,2006,21(3):30-33.
[13] 李捷,王有科.不同杏品種花的抗寒性研究[J].北方園藝,2009(8):47-50.
[14] 李捷,王有科.四個杏品種抗寒性的比較研究[J].甘肅農業大學學報,2010(1):37-40.
[15] 黃永紅,沈洪波,陳學森.杏樹抗寒生理研究初報[J].山東農業大學學報(自然科學版),2005,35(2):191-195.
[16] WallisF G,Wang H,Guerra D J.Expression ofa synthetic an-tifreeze protein in potato reduces electrolyte release at freezing temperatures[J].PlantMolecular Biology,1997,35(3):323 -330.
[17] 王飛,陳登文,王卿,等.杏品種的需寒量與抗寒性的相關研究[J].中國農業科學,2001,34(5):465-468.
[18] 潘曉云,王根軒,曹琴東.蘭州地區引種的美國扁桃的越冬傷害與臨界致死低溫[J].園藝學報,2002,29(1):63-65.
[19] 蔣寶,郭春會,梅立新,等.沙地植物長柄扁桃抗寒性的研究[J].西北農林科技大學學報:自然科學版,2008,36(5):92 -96,102.
[20] 龔鵬,李秀霞,李增萍,等.低溫脅迫對扁桃枝條細胞膜系統和滲透調節物質的影響[J].新疆農業科學,2007,44(5):561 -566.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
