
基于培養環境的混合狀態進行增光處理的新型光發酵制氫工藝
2014-07-01 01:13:34鄧明達
河南科技 2014年4期
關鍵詞:實驗
鄧明達
(3M中國有限公司,上海 200336)
基于培養環境的混合狀態進行增光處理的新型光發酵制氫工藝
鄧明達
(3M中國有限公司,上海 200336)
實驗研究了不同培養環境的混合狀態下光照強度對光合細菌生長和光發酵產氫的影響,提出靜態培養和動態培養狀態下的光飽和點分別為4000lux和6000lux。分析了Rhodobacter sphaeroides ZX-5培養環境中,光照強度隨菌體細胞濃度及光傳播距離增加而衰減的變化規律,得到了光在菌液傳遞過程中衰減的數學模型,即I=I0exp[-(0.4 814+0.2 991 OD660nm)·L]。首次提出了基于培養環境的混合狀態進行增光處理的新型光發酵制氫策略,使得最大產氫速率達到161.7mlH2/lh。
生物制氫;球形紅假單胞菌;光照強度;產氫速率
隨著社會文明的不斷進步和工業化程度的不斷提高,人類對能源的需求日益增加,有限的化石能源已經不能滿足世界各國對能源的需求。能源短缺,環境污染是人類未來面臨的難題,尋找可再生的清潔能源已成為世界范圍內能源界及其相關領域關注的焦點問題。化石燃料燃燒時生成CHx、COx、NOx、SOx、灰塵、煙霧、焦油等有機污染物,造成了嚴重的環境污染并使全球氣候發生變化[1]。
生物制氫所用的原料是城市污水、生活垃圾、動物糞便等有機廢物,通過發酵細菌或光合細菌的生物降解處理可以獲得氫氣,同時凈化水質,達到保護環境的作用。因此,無論從環境保護,還是從新能源開發的角度來看,生物質制氫都具有很大的發展前途,它不僅能給人們提供清潔的能源,又能處理有機廢物,保護環境,獲得可觀的經濟效益,是一條可持續發展的道路[2-4]。生物制氫作為一種符合可持續發展戰略的方向,已在世界上引起廣泛重視,迄今為止,國際上已有的生物制氫方法有藍細菌和綠藻光分解制氫[5]、體外酶合成制氫[6]、厭氧發酵產氫[7-11]、光發酵產氫[12-14]以及厭氧-光合細菌耦合發酵產氫[15-17]等多種。
光合細菌簡稱PSB(Photosynthetic Bacteria),是一群能在厭氧或好氧條件下,以光源作為能源分解有機物進行光合作用的細菌[18]。光合細菌制氫過程中,影響光合產氫的因素很多,主要有菌株特性、溫度、光照條件、pH值、種齡、C源、N源以及與氫代謝相關的三種酶的酶活等因素,其中最大的問題就是光的轉化效率。光在光生物反應器(Photobioreactor,PBR)內隨著光程的增加光強急劇下降,光強在反應器內部的分布很不均勻;同時,有些底物在代謝過程中,往往會產生顏色變化,更進一步阻礙光的傳遞。另一方面,隨著細胞的代謝生長,PBR內透光率隨之下降,因此賴以光能的產氫效率會顯著下降。這已成為PBR光合菌制氫研究中亟待解決的技術難題[19-21]。
本實驗研究了光強對光合生物產氫的影響,模擬光在生物反應器中的傳遞,并通過靜態培養和動態培養的比較,指出光合細菌光飽和點并非固定不變的,而是隨著培養環境的混合狀態和菌體生理條件的變化而改變的,同時確立了基于培養環境的混合狀態進行增光處理的新型光發酵制氫策略,大大提高光合細菌的產氫速率。
1 材料與方法
1.1 菌種和培養條件
菌種:Rhodobacter sphaeroidesZX-5。
種子培養:將1ml種子甘油管倒入裝有100mlRCVBN種子培養基[22]的250ml三角瓶中,pH=7±0.1,180rpm,30±1℃,遮光好氧培養,約20h接種于產氫培養基。
產氫培養:將1ml種子液加入裝有34mlRCVBN產氫培養基的38ml光合反應器中,pH=7±0.1,30±1℃,光照厭氧培養,并采用60ml無菌注射器計量氫氣產量。
1.2 光衰減動力學的測定
取一系列菌體濃度不同的培養液,注入比色皿中,并通過改變比色皿的個數模擬反應器中光程的增加,用光照度計測量光通過不同濃度的菌液時光強的衰減程度;入射光的初始光強為7 000lux。
1.3 實驗器材與檢測方法
光源為60W白熾燈;pH值測量采用FE20型實驗室pH計(Mettler-Toledo儀器有限公司);生物量的測量以OD660nm表示,采用Spectrumlab22pc型分光光度計(上海棱光技術有限公司);光照度的測量采用ZDS-10自動量程照度計(上海市嘉定學聯儀表廠);氣體成分及氫氣含量的測定采用尾氣質譜分析儀(Extrel CMS Max 300-LG,USA);動態培養采用TS-1型脫色搖床(江蘇海門市其林貝爾儀器制造有限公司)。
2 實驗結果與討論
2.1 光照強度對光合細菌生長和產氫性能的影響
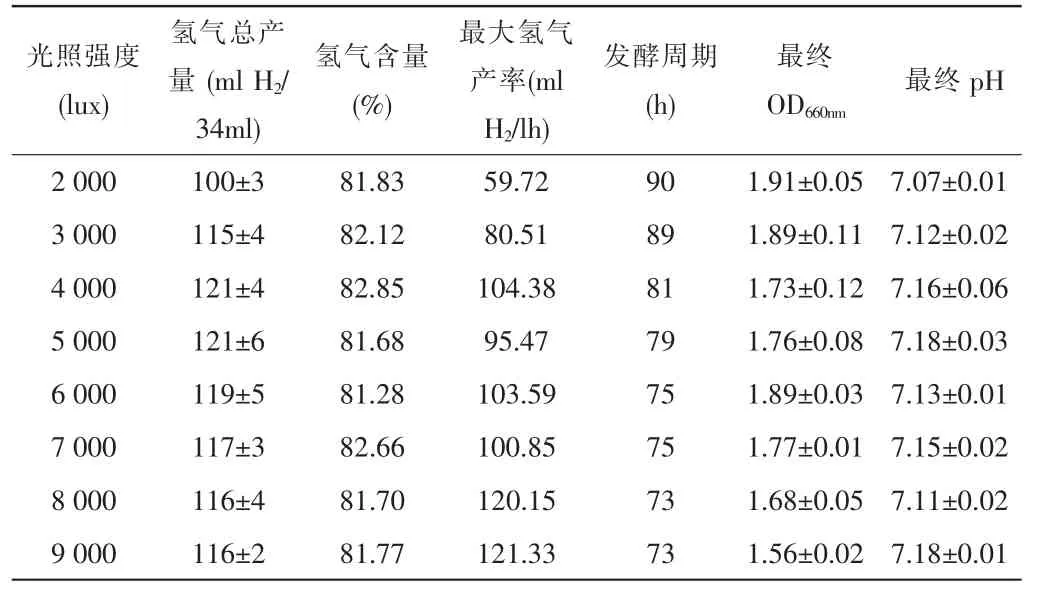
表1 -光照強度對光合細菌生長和產氫性能的影響
實驗研究了不同光照強度(分別為2 000、3 000、4 000、5 000、6 000、7 000、8 000、9 000lux)對光合細菌產氫能力的影響。從表1可知,隨著光照強度的增加,最大產氫速率逐漸增加,并且發酵周期由90h減少至73h,但pH幾乎不隨光強的變化而變化。另外,光強從2 000lux提高到4 000lux時,氫氣的產量隨之增加,當光強為4 000lux時產量最大,達到121mlH2/34ml culture medium;繼續提高光強至9 000lux,氫氣產量反而略有下降。
通過本實驗說明Rhodobacter sphaeroidesZX-5的最適光強為4 000lux,此時已達到了“光飽和”現象。
2.2 光衰減動力學模型的建立
在光合細菌Rhodobacter sphaeroidesZX-5的光發酵體系中,以6 800lux為初始入射光光強,測量菌體濃度(以OD660nm表示)和光程距離的變化對光強在反應器內部分布的影響,結果見圖1。從圖1可以看出,在Rhodobacter sphaeroidesZX-5的光發酵體系中,隨著菌體濃度和光程距離的增加,反應器內部的光照強度迅速衰減。這說明:在反應器內部的同一位置,光強是隨著菌體濃度的增加而變化的。在光發酵初期,菌體細胞濃度較小,光衰減程度低,而到了光發酵后期,隨著菌體細胞濃度的增加,菌體細胞的遮光作用增強,導致光衰減嚴重。

圖1 光程距離和菌體濃度對光強衰減的影響
通過回歸分析,可得到光照強度在Rhodobacter sphaeroides ZX-5光發酵體系中衰減的數學模型:

其中,I為某一光程距離上的實際光強(lux),I0為初始入射光強度(lux),L為光程距離(cm),OD660nm代表菌體細胞濃度。
光發酵體系中光衰減數學模型的建立,有助于我們模擬光強在反應器內部的分布,并據此對培養過程中對光照不足的位置進行增加光照處理。
2.3 培養環境的不同混合狀態對光發酵產氫的影響
實驗2.1采用動態培養的方式,在實驗過程中,我們發現反應器底部有大量菌體沉淀的現象。因此,我們擬采用動態培養的方式將菌體細胞懸浮于培養體系中,從而使菌液混合更加均勻,同時通過動態培養使氫氣析出迅速,盡量減少底物抑制,觀察能否提高產氫速率。
根據實驗2.1的結果,動態培養選在4000lux光強下進行,實驗結果見表2。
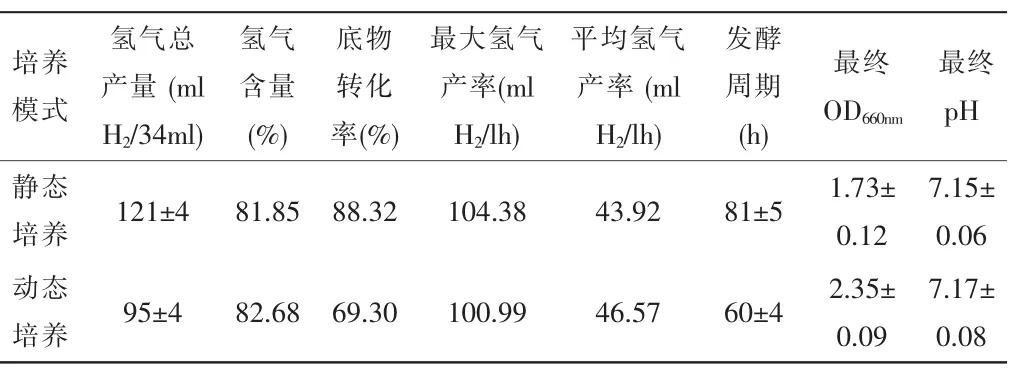
表2 -不同培養模式對光發酵產氫的影響
從表2可知,動態培養與靜態培養相比,縮短了光發酵周期,但是底物轉化率較低,僅為靜態培養時的78.46%。因此,能否提高動態培養的底物轉化率,則直接影響是否可以大幅度提高光發酵的產氫速率。
由圖2可知:在光發酵過程中,動態培養從第8小時開始,菌體濃度(OD660nm)始終大于靜態培養的菌體濃度(OD660nm),并且從第18小時開始,兩個體系的菌體濃度(OD660nm)均趨于穩定,進入菌體生長的穩定期;另外,通過與圖3比較,不管是動態培養還是靜態培養,光發酵產氫過程均發生在菌體細胞生長的穩定期內。
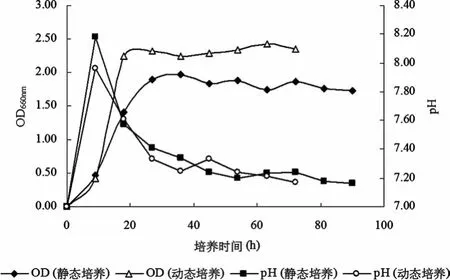
圖2 靜態培養與動態培養過程中OD660nm和pH的變化曲線
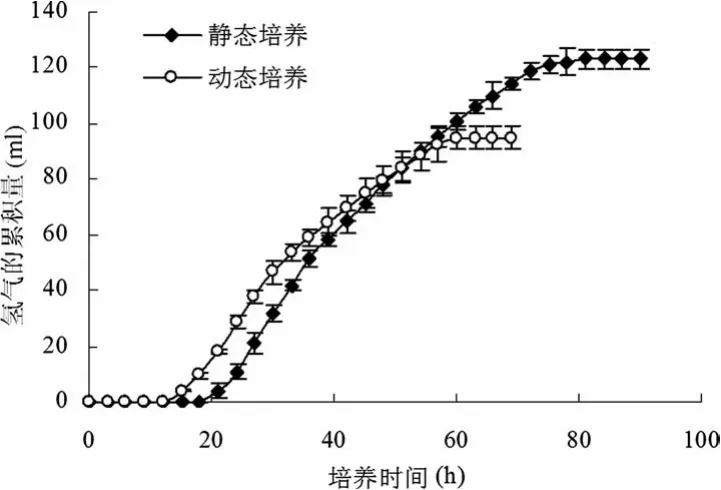
圖3 靜態培養與動態培養的光發酵產氫過程曲線
根據2.2得到的光衰減數學模型可知,OD660nm值大,意味著光在菌液傳遞的過程中衰減現象加劇。從而推斷:可能是因為菌液中光強的不足,導致動態光發酵的底物轉化率偏低。
擬設計增光實驗,通過提高初始入射光光強,觀察是否可以提高動態光發酵的底物轉化率低,從而達到提高光發酵產氫速率的目的。
2.4 基于混合狀態進行增光光發酵制氫工藝策略的確定
2.4.1 實驗原理
根據2.2得到的光衰減數學模型進行估算:
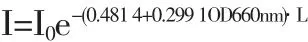
假設初始入射光為4 000lux時,靜態培養結束時OD660nm為1.73,動態培養結束時OD660nm為2.35,見表2。又因為從18小時開始,兩個體系的ODv值均趨于穩定,見圖2,所以假定發酵結束時的OD660nm等于產氫過程中的OD660nm。那么,相同光強在動態培養和靜態培養時菌液中的分布,見表3。通過提高初始入射光的強度,使得光強在動態培養時菌液中的分布與靜態培養時的相當或者優于靜態培養的光照條件,觀察能否提高振蕩光發酵的底物轉化率。

表3 不同混合狀態及初始入射光強度對發酵液中光強分布的影響
與靜態培養相比,初始入射光光強同樣為4 000lux時,在動態培養條件下的衰減要嚴重的多;當把動態培養條件下的初始入射光光強提高至6 000lux時,在光程2cm處,光強度幾乎與靜置發酵時一致,而在反應器內部則優于靜態培養內部的光強分布;繼續將光強提高到7 000lux~8 000lux時,其在反應器內部的光強分布則遠遠優于入射光為4 000lux時的靜態培養內部的光強分布(表3)。
2.4.2 實驗結果
由實驗結果表4可知,通過增加入射光光強,動態培養的氫氣產量上升,OD660nm下降,說明反應器內部光照強度不足是導致動態培養發酵底物轉化率低的原因之一。
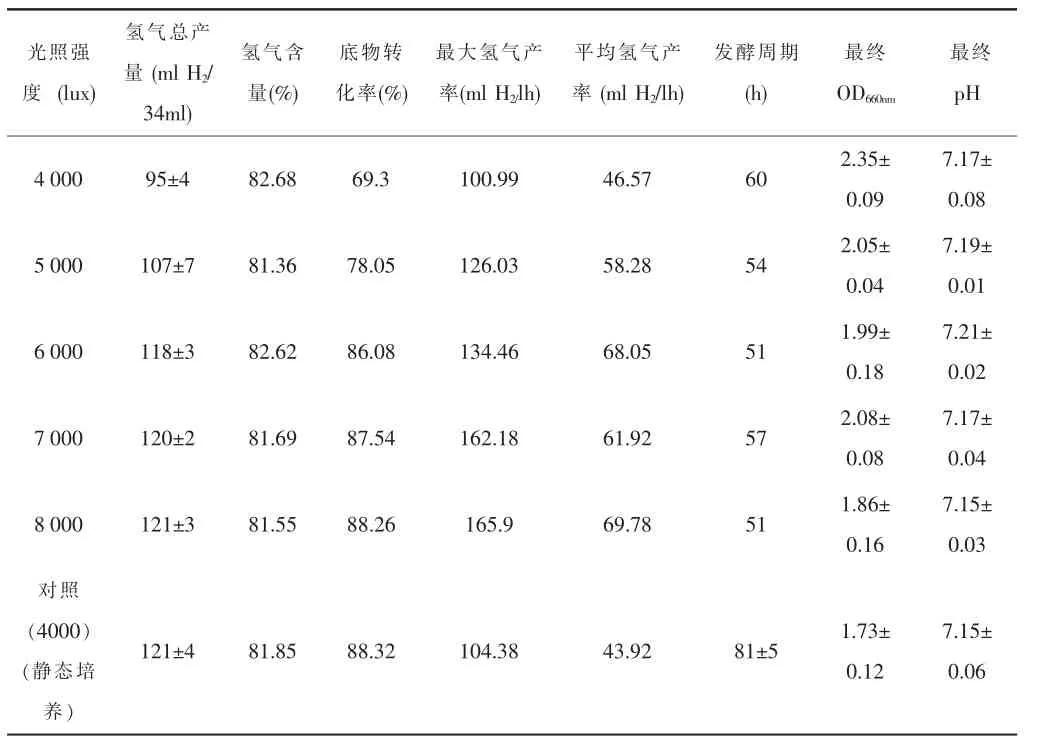
表4 不同培養模式及不同初始入射光強度對光發酵產氫的影響

圖4 動態培養模式下,不同初始入射光強度對底物轉化率和菌體生長(以OD660nm表示)的影響

圖5 動態培養模式下,不同初始入射光強度對最大產氫速率和平均產氫速率的影響
由圖4知,光強從4 000lux增加到8 000lux過程中,OD660nm卻從2.35下降到1.86,與靜態培養的OD非常接近,而光強的增加則對pH影響不大,基本維持穩定在pH:7.18±0.03。另外,隨著光強的增加,氫氣的產量逐漸增加。但是當光強達到6 000lux時,再增加光強,動態培養的氫氣產量不再增加,幾乎維持恒定,說明光強6 000lux為動態光發酵條件下的光飽和點,這與靜態培養時400lux為光飽和點是不一致的。通過該實驗可以得出:光合細菌的光飽和點是隨著外部環境條件的變化而發生變化的,并非固定不變的。
由圖5知:通過光強的增加,動態培養的發酵周期縮短,氫氣得率提高,進而使得平均產氫反應速率和最大產氫速率均得到大幅度的提高。
3 實驗結論和研究趨勢
3.1 實驗結論
本文研究了不同光照強度對光合細菌產氫的影響,確定了3 000lux為靜態培養條件下的最佳光照強度;同時模擬了光在菌液中的衰減情況,建立了光衰減動力學數學模型;實驗提出了基于培養環境的混合狀態進行增光處理的新型光發酵制氫工藝,提高了光發酵的最大產氫速率。
3.2 研究趨勢
為了提高產氫速率和產氫量,光合細菌制氫系統目前還存在以下3個制約因素:(1)生物培養設備的太陽能轉化效率;(2) H2合成過程;(3)生物反應器的設計和成本。
由于光合微生物制氫過程存在的厭養、光轉化率低、連續高效率產氫時間短等問題,因此,在深入進行產氫生化機制研究,高產菌株選育的同時,應著重以下幾個方面的研究:
(1)充分利用光合微生物及其它微生物的多樣性和互助性,采用共培養或分步培養技術,利用微生物菌群制氫,解決酶抑制,利用基質種類單一,有機物轉化利用率低等問題。
(2)深入進行光能吸收、轉化和利用方面的研究,用光譜耦合,光有效導入和控制等技術提高光轉化效率。
(3)開展多種原料利用研究,進行不同原料下高產菌株的選育,及其影響因素研究,最佳工藝研究,注重利用污染源為原料的產氫技術研究。
(4)光合生物反應器的研究。由于光合微生物產氫效率的高低與太陽光利用轉化率有直接的聯系,因此反應器結構研究、光照控制系統的研究也是制氫技術的重要課題。
[1]Bockris JO’M.The origin of ideas on hydrogen economy and its solution to the decay of the environment[J].Int J Hydrogen Energy, 2002,27:73l-740.
[2]DunnS.Hydrogenfutures:towardasustainableenergy system[J].Int J Hydrogen Energy,2002,27:235-264.
[3]傅秀梅,王亞楠,王長云,鹿守本,管華詩.生物制氫——能源、資源、環境與經濟可持續發展策略[J].中國生物工程雜志, 2007,2007(2):119-125.
[4]肖建民.論氫能源與氫能源系統[J].世界科技研究與進展,1997,19(1):82-86.
[5]MelisA.Greenalgahydrogenproduction:progress, challenges and prospects[J].Int J Hydrogen Energy,2002,27:1217-1228.
[6]李永峰,任南琪,胡立杰,史英.生物制氫系統及其產氫機制[J].能源環境保護,2005,19(2):5-7.
[7]Lin CY,Lay CH.A nutrient formulation for fermentative hydrogen production using anaerobic sewage sludge microflora[J].Int J Hydrogen Energy,2005,30:285-292.
[8]Zhang YF,Shen JQ.Effect of temperature and iron concentration on the growth and hydrogen production of mixed bacteria[J].Int J Hydrogen Energy,2006,31:441-446.
[9]Hawkes FR,Dinsdale R,Hawkes DL,Huss I.Sustainable fermentativehydrogenproduction:challengesforprocess optimization[J].Int J Hydrogen Energy,2002,27:1339-1347.
[10]Li XZ,Wu JS,Sun DL.Hydrogen sulphide and volatile fatty acid removal from foul air in a fibrous bed bioreactor[J].Water Sci Technol,1998,38:323-329.
[11]N.Q.Ren,H.Chua,S.Y.Chan,etc.Assessingoptimal fermentation type for bio-hydrogen production in continuous-flow acidogenic reactors[J].Bioresource Technol,2007,98,1774-1780.
[12]Toshihiko Kondo,Tatsuki Wakayamab,Jun Miyakeb. Efficient hydrogen production using a multi-layered photobioreactor and a photosynthetic bacterium mutant with reduced pigment[J].Int J Hydrogen Energy,2006,31:1522-1526.
[13]E.Nakada,S.Nishikata,Y.Asada,J.Miyake. Photosynthetic bacterial hydrogen production combined with a fuel cell[J].Int J Hydrogen Energy,1999,24:1053-1057.
[14]YongzhenTao,YanlingHe,YongqiangWu,etal. Characteristics of a new photosynthetic bacterial strain for hydrogen production and its application in wastewater treatment[J].Int J Hydrogen Energy,2008,33:963-973.
[15]李冬敏,陳洪章,李佐虎.生物制氫技術的研究進展[J].生物技術通報,2003,4:1-5.
[16]Yongzhen Tao,Yang Chen,Yongqiang Wu,etc.High hydrogen yield from a two-step process of dark-and photofermentation of sucrose[J].Int J Hydrogen Energy,2007,32:200-206.
[17]Mark D.Redwood,Lynne E.Macaskie.A two-stage,twoorganism process for biohydrogen from glucose[J].Int J Hydrogen Energy,2006,31:1514-1521.
[18]Asada Y,Ikenaka K.Photobiological hydrogen production [J].J Biosci Bioeng,1999,88:1-6.
[19]王昶,賈士儒,賈慶竹,馬少娜.光合生物制氫技術[J].生物加工過程,2005,3(4):9-13.
[20]Eiju Nakada,Yasuo Asada,Takaaki Arai,Jun Miyake. Light Penetration into Cell Suspensions of Photosynthetic Bacteria and Relation to Hydrogen Production[J].J Ferment Bioeng,1995,80 (1):53-57.
X505
A
1003-5168(2014)04-0167-04
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
