外源稀土Ce3+對鳳眼蓮鈣相關光合作用的影響
2014-08-07 14:10:50宋雪梅趙仕林四川師范大學化學與材料科學學院四川成都60066農田生態服務能力建設四川省高校工程中心四川成都60066四川大學建筑與環境學院四川成都60065
中國環境科學 2014年9期
關鍵詞:植物
宋雪梅,夏 超,廖 洋,2*,趙仕林,余 江(.四川師范大學化學與材料科學學院,四川 成都 60066;2.農田生態服務能力建設四川省高校工程中心,四川 成都 60066;.四川大學建筑與環境學院,四川 成都 60065)
外源稀土Ce3+對鳳眼蓮鈣相關光合作用的影響
宋雪梅1,夏 超1,廖 洋1,2*,趙仕林1,余 江3(1.四川師范大學化學與材料科學學院,四川 成都 610066;2.農田生態服務能力建設四川省高校工程中心,四川 成都 610066;3.四川大學建筑與環境學院,四川 成都 610065)
以造成水體污染的主要水生生物之一的鳳眼蓮為研究對象,探究外源稀土 Ce3+對鳳眼蓮鈣相關光合作用的影響.相比于對照組,營養液中缺鈣但添加外源稀土Ce3+的條件下,鳳眼蓮葉綠素含量減少8.31%、葉綠體熒光產額分別減少6.14%(440nm)和3.82%(480nm)、類囊體膜上Mg2+-ATP酶和Ca2+-ATP酶分別減少1.93%和5.05%,蛋白質含量減少3.94%;但較缺鈣的營養液處理組相應指標分別增加了9.89%、15.21%和3.05%、8.08%和10.26%、10.16%.添加Ce3+的營養液處理組的鳳眼蓮鈣相關光合作用指標同樣顯著提高.結果表明,Ce3+有類似于 Ca2+的特性,適量的 Ce3+可以補償替代鈣,對植物光合作用有促進效果,增加植物生物量,促進鳳眼蓮型水生植物的生長,對水體富營養化進程的發展起正催化作用.
Ce3+;Ca2+;鳳眼蓮;光合作用
稀土元素及其化合物因具有獨特的光、磁、電子性能,被譽為“工業味精”,不僅廣泛應用于輕工、冶金、化工、農業等傳統行業,而且在電子、信息、新能源等眾多高新技術領域也在逐步開發應用[1].然而,隨著稀土應用強度的增加,稀土元素及其化合物大量進入環境,我國淡水域中的稀土元素含量比世界平均水平(0.64μg/L)高出6倍之多[2].據報道,長江上游主要支流及部分干流的豐水期在2007~2009年稀土含量為9.61~63.37μg/L,枯水期稀土含量為 12.11~90.20μg/L,其中以輕稀土離子為主,且稀土元素含量比20世紀80年代豐水期已高出100多倍[3-6].
稀土元素能夠在一定濃度范圍內能增強植物的光合作用,有效促進植物生物量的增加,影響植物的外部形態和生長發育[7].宋凌云等[8]研究表明稀土元素對水生植物的光合作用有促進效應,可以促進水生植物對N、P元素的吸收,導致水生植物大量繁殖,引起水體富營養化;張貝克等[9]研究表明外源稀土鈰對紫背浮萍生長的影響中也具有類似作用.同時,稀土 Ce3+與 Ca2+具有許多共同的性質,如 Ce3+半徑與 Ca2+的半徑相近[10],因而在生物體內常作為Ca的拮抗劑或發生取代作用,研究Ca對植物的生理功能[11-12]. Hong等[13]研究發現,部分稀土元素可以促進酶的活性.
鳳眼蓮(Eichhornia crassipes),俗稱水葫蘆,浮水植物,廣泛生長于我國南方水域,具有較強的凈化污水能力[14],但大量繁殖的水葫蘆覆蓋水域表面,造成水體富營養化,破壞水體使用功能.大量研究表明水體中氮磷等營養元素可以促進鳳眼蓮的生長,進而促進水體富營養化的發生發展
[15],然而稀土元素對鳳眼蓮生長的影響及其機理的研究甚少.因此,本文以鳳眼蓮為實驗材料,研究稀土元素是否具有類似鈣的生理作用,進而闡明稀土元素對長江上游水域水體富營養化的影響機理,為防治水體富營養化提供理論依據.
1 材料與方法
1.1 材料培養
鳳眼蓮取自成都東郊三環外一水塘.將鳳眼蓮用清水清洗干凈,放入實驗室外水池中培養,待用.將鳳眼蓮橫生的匍匐莖割成幾段或帶根切離幾個腋芽,選擇帶有3~4片嫩葉的大小形態基本相同的鳳眼蓮,放入1000mL燒杯中進行實驗,并對杯身用黑色塑料袋進行遮光處理.
1.2 營養液配置
營養液配制是以改良后Hoagland營養液為基礎,并參考其他文獻[16-17]確定配方.
1.3 培養方法
Ce3+最適濃度確定的培養,將稀土 Ce3+的濃度梯度設置為以下五個濃度梯度:0、0.6、1.2、1.8、2.4mg/L,每個濃度梯度設置3個平行樣.
將燒杯置于 25℃培養箱中,模擬自然光(光照強度3500~4500lx,光照比13:11),培養7d后測定各試樣中鳳眼蓮的葉綠素含量.
培養液分為4個組:(1)改良 Hoagland 營養液對照組;(2)缺 Ca2+的 Hoagland 營養液處理組;(3)添加1.20mg/L Ce3+的Hoagland營養液處理組;(4)添加1.20mg/LCe3+的缺Ca2+的Hoagland營養液處理組.每組設置3個平行樣.
鳳眼蓮培養7d后測定試樣中葉綠素、蛋白質、葉綠體熒光光譜、Mg2+-ATP和 Ca2+-ATP酶活性,根據《植物生理學》中的方法測定[18-19].鳳眼蓮葉片清洗、切片后,采用電子顯微鏡觀察其細胞形態結構.
1.4 數據處理
數據均由均值±標準差表示,統計處理采用軟件SPSS 16.0,顯著性水平設置為P<0.05.
2 結果分析與討論
2.1 不同Ce3+濃度的鳳眼蓮葉綠素含量
如圖1所示,葉綠素含量隨稀土Ce3+濃度的增加先上升再下降.稀土濃度在 0.6,1.2,1.8mg/L時鳳眼蓮葉綠素含量均比對照組高,且 1.2mg/L時葉綠素含量達到最大值.稀土濃度為 1.8mg/L時葉綠素含量下降,與對照組相差無幾,稀土濃度為2.4mg/L時鳳眼蓮葉綠素含量降到最低.
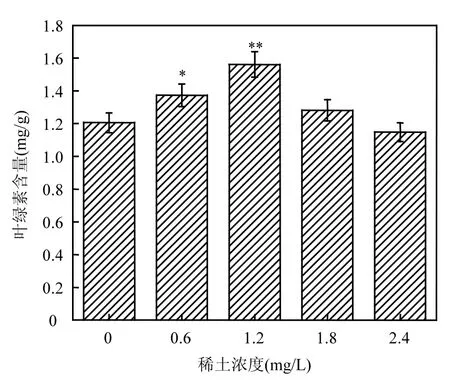
圖1 不同稀土Ce3+濃度對鳳眼蓮葉綠素含量影響Fig.1 Amount of chlorophyll in Eichhornia crassipes with different concentration of Ce3+
葉綠素含量的高低可在一定程度上反映出植物光合作用的強弱以及植物的生長情況.實驗結果表明適宜濃度(0~1.2mg/L)的稀土元素,可以促進鳳眼蓮葉綠素含量增加,進而促進鳳眼蓮生長發育,其中最適稀土濃度為 1.2mg/L;高濃度(2.4mg/L)稀土會抑制鳳眼蓮的生長,甚至會產生類似重金屬的毒性作用.
2.2 葉片電子顯微鏡圖
由圖 2可知,葉片中細胞呈半月狀且處于張開狀態,兩兩相對而生的保衛細胞圍成了氣孔,保衛細胞中所含的葉綠體數量比對照組明顯增多.
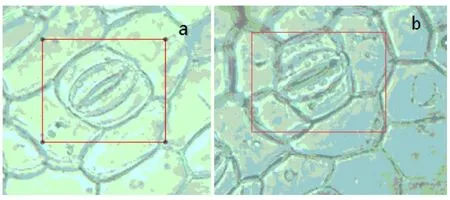
圖2 鳳眼蓮葉片(a)對照組和(b)1.2mg/L Ce3+處理組細胞電子顯微鏡圖(×400)Fig.2 The electron microscope images of Eichhornia crassipes leaf cells (magnification: ×400)
植物葉片內含葉綠體的保衛細胞的氣孔作為植物與外界環境進行氣體交換的重要通道,在調節植物光合作用中扮演著至關重要的角色
[20-21],其會隨著光合作用強度的增大而增大.受控實驗表明,添加 Ce3+營養液處理組鳳眼蓮葉片中葉綠體較對照組明顯增多,表明 Ce3+可以促進鳳眼蓮的光合作用.因此適宜濃度的稀土可以促進鳳眼蓮的生長,其中 1.2mg/LCe3+是促進鳳眼蓮生長的最適濃度.
2.3 葉綠素含量
如圖3所示,相比于對照組,缺Ca2+處理組鳳眼蓮葉綠素含量降低了0.19mg/g,添加Ce3+處理組葉綠素含量增加了0.17mg/g.缺Ca2+添加Ce3+處理組的葉綠素含量少于對照組,但比缺 Ca2+處理組增加了 0.1mg/g.統計表明,添加 Ce3+處理組鳳眼蓮葉綠素含量明顯高于對照組(P<0.05),比對照組增加了 18.2%;缺 Ca2+處理組的鳳眼蓮葉綠素含量明顯低于對照組,減少了 16.31%;缺 Ca2+添加 Ce3+處理組的鳳眼蓮葉綠素含量較對照組變化不明顯.
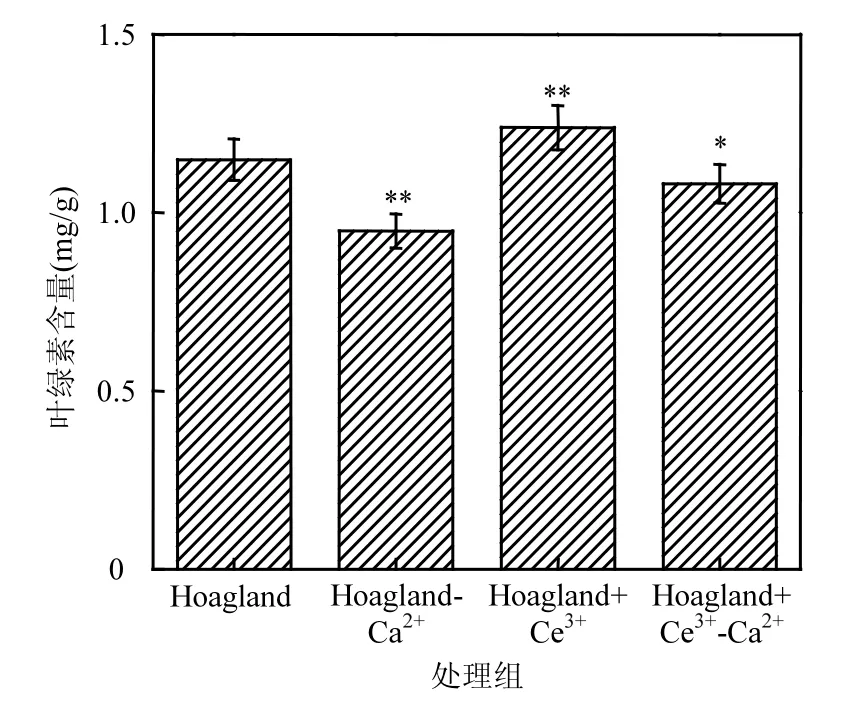
圖3 不同培養液中鳳眼蓮葉綠素含量Fig.3 The amount of chlorophyll of Eichhornia crassipes in different culture solutions
鈣元素是植物體生長發育所必需的礦質營養元素,作為葉綠體基本單元的重要組分,Ca2+在光合作用中扮演重要角色[22-25].若植物體內缺少Ca2+則會直接影響植物的光合作用進而影響生長速率[26-27].而植物生長過程中最重要的是光合作用,植物葉片吸收光和熱,通過光合作用產生淀粉、脂肪、蛋白質等有機物,實現光能轉化為化學能.綠色植物通過葉綠體中的光合作用將光能轉換為化學能并將其儲存在有機體中.
葉綠素是一類與光合作用有關的最重要色素.添加 Ce3+處理組鳳眼蓮葉綠素含量的增加表明稀土可以促進植物的光合作用,而在缺 Ca2+條件下的鳳眼蓮葉綠素含量下降,說明鈣對植物葉綠素的合成起主要作用,鈣的缺少抑制了葉綠素對光能的吸收.但在缺Ca2+情況下添加了Ce3+,葉綠素含量的部分增加表明 Ce3+可以減弱缺 Ca2+導致的損傷,補償性代替Ca2+,保持植物進行光合作用的能力.
2.4 葉綠體熒光光譜
由圖4可知,在440nm的光激發下,各處理組中鳳眼蓮葉綠體熒光光譜在680nm附近有一較強發射峰.加Ce3+處理組的熒光峰值增加,比對照組上升了 5.4%,缺 Ca2+處理組峰值最低,比對照組下降了21.35%,缺Ca2+添加Ce3+處理組熒光峰值比缺Ca2+處理組上升了15.21%.

圖4 不同培養液中鳳眼蓮葉綠體熒光(440nm)Fig.4 Chlorophyll fluorescence spectrum (λ=440nm) of Eichhornia crassipes in different culture solutions
由圖5可知,在480nm的光激發下,各處理組中鳳眼蓮葉綠體熒光光譜在660nm附近有一較強發射峰,在680nm附近有一較弱發射峰.加Ce3+處理組的熒光峰值增加,比對照組上升了 9.16%,缺Ca2+處理組峰值最低,比對照組下降了6.87%,缺Ca2+添加Ce3+處理組熒光峰值比缺Ca2+處理組上升了3.05%.
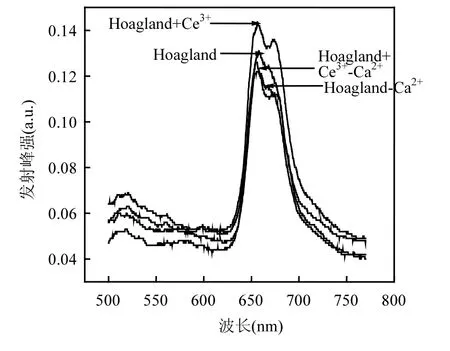
圖5 不同培養液中鳳眼蓮葉綠體熒光(480nm)Fig.5 Chlorophyll fluorescence spectrum (λ=480nm) of Eichhornia crassipes in different culture solutions
葉綠素吸收光能后通過類胡蘿卜素和葉綠素a、b傳遞給中心色素(PSⅠ和PSⅡ),再經過光合磷酸化將電能轉化為活躍的化學能.在光系統II(PS II)中含有析氧中心復合體,在此發生原初反應,最初電子供體H2O被氧化并釋放氧.該中心結合了特有的Ca2+,在水氧化過程中有重要作用.已知680nm左右的熒光發射峰來自PSII, 440nm和480nm的光激發下,缺Ca2+處理組中鳳眼蓮葉綠體的PSII熒光發射峰峰強最低,兩個光系統的電子流被顯著抑制,這對從 PSII 中獲得電子有嚴重影響,因此降低了光能到電能的轉化效率,降低了 PSII中心色素葉綠素 a(P680)的熒光產額;而缺Ca2+處理組中添加了Ce3+后,鳳眼蓮葉綠體的PSII熒光發射峰峰強相比于單獨缺Ca2+組要高,表明經過 Ce3+處理后被胡蘿卜素和葉綠素 b吸收的光能有效傳遞到PSII作用中心色素葉綠素a后使其熒光產額提高.這表明Ce3+在析氧中心具有和Ca2+相似的功能,補償了Ca2+缺失導致的光能轉化率,所以Ce3+可以減弱缺Ca2+導致的光能轉化率下降的損傷.
2.5 葉綠體Mg2+-ATP酶和Ca2+-ATP酶活性
由圖6可知,相比于對照組,缺Ca2+處理組鳳眼蓮葉綠體的Mg2+-ATP酶和Ca2+-ATP酶活性分別降低了 25.355,54.73μmol/(mg?min),加 Ce3+處理組Mg2+-ATP酶和Ca2+-ATP酶活性分別增加了37.89,35.50μmol/(mg?min),缺Ca2+添加Ce3+處理組的葉綠體的Mg2+-ATP酶和Ca2+-ATP酶活性比缺 Ca2+處理組分別增加了 20.45, 24.22μmol/(mg?min),但低于對照組.統計分析發現,添加Ce3+處理組鳳眼蓮葉綠體Mg2+-ATP酶和Ca2+-ATP酶活性明顯高于對照組(P<0.05),缺Ca2+處理組的葉綠體 Mg2+-ATP酶和 Ca2+-ATP酶活性明顯低于對照組;缺Ca2+添加Ce3+處理組的葉綠體Mg2+-ATP酶和Ca2+-ATP酶活性較對照組變化不明顯.
植物以三磷酸腺苷(ATP)和NADPH的形式通過非循環和循環光合磷酸化過程,將電能轉換為活躍的化學能,產生的化學能被暫時儲存在ATP中,這耦合了類囊體膜上的電子傳輸和光合磷酸化,并最終將二氧化碳和水轉化為碳水化合物和氧氣.ATP合成酶包括 Mg2+-ATP酶和Ca2+-ATP酶,是整個光合磷酸化的終端.在缺Ca2+處理組中鳳眼蓮的該兩種酶相比于對照組下降較多,表明葉綠體中兩個光合磷酸化的效能被抑制,由缺 Ca2+導致的兩種光合磷酸化的抑制和葉綠體的耦合水平有密切聯系,缺 Ca2+還導致PSII反應中心復合體構造和功能的改變,這對PSII、PSI和Cytb6f化合物間電子傳遞產生影響并降低了整個光合磷酸化的效率;在加 Ce3+缺Ca2+處理組中鳳眼蓮兩種酶活性得到了一定修復,減輕了因缺Ca2+而引起的損傷.ATP合成酶的減少可能歸因于 Ca2+的缺失,特別是類囊體膜上Ca2+-ATP酶含量的減少,而這種情況通過添加Ce3+得到一定程度改善.總之,Ce3+減少了由缺Ca2+導致的光合磷酸化效率的降低,在一定程度上,源于其與Ca2+相似的結構.

圖6 不同培養液中鳳眼蓮葉綠體Mg2+-ATP酶活性和Ca2+-ATP酶活性Fig.6 the Mg2+-ATP and the Ca2+-ATP activity of Chloroplast of Eichhornia crassipes in different culture solutions
2.6 蛋白質含量
由圖7可知,相比于對照組,缺Ca2+處理組鳳眼蓮的蛋白質含量降低了0.38mg/g,添加Ce3+處理組的蛋白質含量增加了0.51mg/g,缺Ca2+添加Ce3+處理組的蛋白質含量比缺 Ca2+處理組增加了 0.27mg/g,但低于對照組.統計發現,添加 Ce3+處理組鳳眼蓮蛋白質含量顯著高于對照組(P<0.05);缺 Ca2+處理組的蛋白質含量顯著低于對照組;缺Ca2+添加Ce3+處理組的蛋白質含量較對照組變化不明顯.

圖7 不同培養液中鳳眼蓮蛋白質含量Fig.7 the protein content of Eichhornia crassipes indifferent culture solutions
植物細胞代謝的主要能源物質是淀粉,而主要的結構物質是蛋白質,光合作用中不管是光反應還是暗反應都需要多種酶的催化作用, 其大部分酶都是蛋白質.實驗表明缺 Ca2+處理組的鳳眼蓮蛋白質含量相比于對照組降低較明顯,但加Ce3+到缺 Ca2+的處理組中,明顯看出 Ce3+減輕了缺Ca2+的損傷,蛋白質含量有多增加.在添加Ce3+的情況下,降低了因缺Ca2+而引起的損害程度,相比于單獨缺 Ca2+處理組,Ce3+與 Ca2+有著相似的功能,Ce3+發揮著Ca2+的作用,促進酶的產生,保證光合作用的正常運行.
由統計分析可知,葉綠素含量和蛋白質含量之間的Pearson相關系數是0.935,表明兩者具有相關性;葉綠素含量和 Ca2+-ATP酶活性之間的Pearson相關系數是0.968,并且在0.05水平上顯著相關;蛋白質含量和 Mg2+-ATP酶活性之間的Pearson相關系數是0.998,并且在0.01 水平上顯著相關.表明在不同處理條件下光合作用各指標間存在一定的相關性且相關性較顯著,最終表現為稀土離子對植物鈣相關光合作用的促進作用.
3 結論
3.1 不同 Ce3+濃度可以影響鳳眼蓮的生長,濃度為 0~1.2mg/L對鳳眼蓮的生長有促進作用,其中 1.2mg/L Ce3+促進作用最明顯,但高濃度
(2.4mg/L)Ce3+可抑制鳳眼蓮的生長,甚至產生毒害作用.
3.2 適宜濃度的Ce3+可能替代Ca2+影響與鈣相關的光合作用,在一定濃度范圍內促進植物的光合作用,增加有機物質的合成,進而增加其生物量.在長江上游地區的四川支流中,由于稀土元素的協同作用,可能導致植物生長,最終表現為促進水生植物鳳眼蓮的生長,推進水體富營養化進程.
[1]徐光憲.稀土 [M]. 北京:冶金工業出版社, 2005.
[2]趙仕林,廖 洋,李瑞楨,等.四川地區稀土污染源調查及防控對策研究 [M]. 決策參考, 2009,184-203.
[3]Yu J, Chen W Q, Li J Q, et al. Study of the promoting mechanism of rare earth on water eutrophication in the upper reaches of the Yangtze River [J]. Disaster Advances, 2012,5(4):1595-1602.
[4]Matthew I L, Karen H J. Rare earth elements (REE) and yttrium in stream waters, stream sediments, and Fe-Mn oxyhydroxides: fractionation, speciation, and controls over REE+Y patterns in the surface environment [J]. Geochimicaet Comsmochimica Acta, 2008,72(24):5962-5983.
[5]胡勤海,金明亮.稀土元素在水體中的環境化學行為及其生物效應 [J]. 農業環境保護, 2006,25(5):274-277.
[6]張 輝,馮 嘉,朱為方.稀土高背景區生物鏈中稀土元素的分布特征 [J]. 中國稀土學報, 1999,17:365-368.
[7]錢 蕓,戴樹桂,劉廣良,等.硝酸鑭對銅綠微囊藻生長特性的影響 [J]. 中國環境科學, 2003,23(1):7-11.
[8]宋凌云,胡文月,趙繼貞.稀土元素鑭對滿江紅魚腥藻的生理影響 [J]. 北京大學學報(自然科學版), 2000,36(6):123-128.
[9]張貝克,余 江,廖 洋,等.外源稀土 Ce3+對紫背浮萍光合作用和水體富營養化的影響 [J]. 中國稀土學報, 2012,30(6):236-242.
[10]周曉波,魏幼璋.稀土離子與Ca2+在生物體內的相互作用機制及應用 [J]. 生命科學研究, 1999,3(1):30-35.
[11]王偉列,涂楚橋,王光輝.稀土金屬離子與酶分子的相互作用研究進展 [J]. 稀土, 1998,19(3):57-65.
[12]肖鳳娟,常 虹,劉德龍,等.稀土離子與鈣調素相互作用的研究進展 [J]. 稀土, 2003,2(6):64-68.
[13]Hong F S, Liu C, Zheng L. Formation of complexes of Rubisco-Rubisco activase from La3+, Ce3+treatment spinach [J]. Sci China Ser B, 2005(b),48(1):67-74.
[14]王 智,張志勇,張君倩,等.水葫蘆修復富營養化湖泊水體區域內外底棲動物群落特征 [J]. 中國環境科學, 2012,32(1):142-149.
[15]凌 禎,楊具瑞,于國榮,等.不同植物與水力負荷對人工濕地脫氮除磷的影響 [J]. 中國環境科學, 2011,31(11):1815-1820.
[16]種云霄,胡洪營,錢 易,等.無機氮化合物及pH值對紫背浮萍生長的影響 [J]. 中國環境科學, 2003,23(4):417-421.
[17]丁士明,梁 濤,閆軍才,等.有機配體對稀土元素在小麥體內積累和分異的影響 [J]. 生態學報, 2005,25(11):2888-2894.
[18]陳建勛.植物生理學實驗指導 [M]. 廣州:華南理工大學出版社, 2006.
[19]趙世杰,史國安,董新純.植物生理學實驗指導 [M]. 北京:中國農業科學技術出版社, 2002.
[20]孫 麗,宋長春,黃 耀.沼澤濕地N2O通量特征及N2O與CO2排放間的關系 [J]. 中國環境科學, 2006,26(5):532-536.
[21]Chaerle L, Saibo N, Van Der Straeten D. Tuning the pores: towards engineering plants for improved water use efficiency [J]. Trends Biotechnol., 2005,23(6):308-315.
[22]李青云,葛會波,胡淑明,等.鈉鹽和鈣鹽脅迫對草莓光合作用的影響 [J]. 西北植物學報, 2006,26(8):1713-1717.
[23]肖鵬程,朱端衛,羅 媛,等.環境因素對菹草根和葉細胞質膜 Ca2+-ATPase活性的影響 [J]. 中國環境科學, 2012,32(6):1123-1128.
[24]簡令成,王 紅.Ca2+在植物細rt5443胞對逆境反應和適應中的調節作用 [J]. 植物學通報, 2008,25(3):255-267.
[25]束良佐,孫五三.Ca2+浸種對酸雨傷害玉米幼苗的影響 [J]. 中國環境科學, 2001,21(2):185-188.
[26]Liang W J, Wang M L, Ai X Z. The role of calcium in regulating photosynthesis and related physiologicalindexes of cucumber seedlings under low light intensity and suboptimal temperature stress [J]. Scientia Horticulturae, 2009,123:34-38.
[27]李 強,曹建華,余龍江,等.干旱脅迫過程中外源鈣對忍冬光合生理的影響 [J]. 生態環境學報, 2010,19(10):2291-2296.
Influences of rare earths (Ce3+) on calcium-regulated photosynthesis in Eichhornia crassipes
SONG Xue-mei1, XIA Chao1, LIAO Yang1,2*, ZHAO Shi-lin1, YU Jiang3(1.College of Chemistry and Material, Sichuan Normal University, Chengdu 610068, China;2.Engineering Research Center for the Development of Farmland Ecosystem Service Functions, Sichuan Province Institutions of Higher Education, Chengdu 610068, China;3.College of Architecture and Environment, Sichuan University, Chengdu 610065, China). China Environmental Science, 2014,34(9):2362~2367
Influences of rare earth Ce3+on calcium-regulated photosynthesis in Eichhornia crassipes were investigated. When cultured in Ce3+-containing nutrient solution without Ca2+, the chlorophyll produced in Eichhornia crassipes was found to decrease by 8.31% compared to that of counterparts cultured in Ca2+-containing nutrient solution (control experiment). The corresponding fluorescence quantum yields of chloroplast at 440and 480nm were also decreased by 6.14% and 3.82%, respectively. Additionally, the activities of Mg2+-ATP enzyme and Ca2+-ATP enzyme, and the amount of proteins were also decreased by 1.93%, 5.05% and 3.94%, respectively. However, the above parameters related to the calcium-regulated photosynthesis increased by 9.89%, 15.21%, 3.05%, 8.08%, 10.26% and 10.16%, respectively, when compared with those of Eichhornia crassipes cultured in nutrient solution without Ca2+. The photosynthesis activity of Eichhornia crassipes cultured in the absence of Ca2+had been significantly improved by the use of rare earth Ce3+. The experimental results have shown that compared with Ca2+, Ce3+exhibited similar regulation functions on the growth of Eichhornia crassipes. In the absence of Ca2+, a suitable amount of Ce3+will promote the photosynthesis of plant by increasing the plant biomass, thus resulting in positive catalytic effects on the process of eutrophication.
Ce3+;Ca2+;Eichhornia crassipes;photosynthesis
X171
A
1000-6923(2014)09-2362-06
宋雪梅(1987-),女,四川瀘州人,四川師范大學碩士研究生,主要從事水體污染預防與控制研究.
2014-01-08
國家自然科學基金資助項目(31100374,51173122);四川師范大學校團隊基金(025091)
* 責任作者, 副教授, Liaoyang_sicnu@163.com
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
