AMPK/PPARα/SCAD信號途徑對心肌肥大的調控研究*
2014-08-08 09:02:22黃金賢羅佳妮劉培慶陳少銳黃秋菊潘雪刁臧林泉周四桂
中國病理生理雜志 2014年5期
黃金賢, 羅佳妮, 劉培慶, 陳少銳, 黃秋菊, 潘雪刁, 臧林泉, 周四桂△
(1廣東藥學院臨床藥學系,廣東 廣州 510006; 2中山大學藥學院藥理與毒理學實驗室,廣東 廣州 510006)
我們采用定量蛋白質組學技術比較了16周齡自發性高血壓大鼠和血壓正常大鼠的心肌蛋白質組,首次發現短鏈酰基輔酶A脫氫酶(short-chain acyl-coenzyme A dehydrogenase,SCAD)在自發性高血壓大鼠肥大心肌中的表達顯著降低[1]。SCAD是脂肪酸β氧化的限速酶,對脂肪酸β氧化起著關鍵作用。我們的研究進一步證實,SCAD的蛋白表達及酶活性的顯著下調,可能是導致自發性高血壓大鼠肥厚心肌能量代謝“胚胎型再演”的分子基礎[2];體外苯腎上腺素(phenylephrine,PE)刺激誘導的心肌細胞肥大模型中,SCAD在蛋白及mRNA水平均顯著下調;采用siRNA干擾SCAD后,心肌細胞肥大指標明顯上升,表明SCAD參與了心肌肥大的調控[3]。然而,SCAD調控心肌肥大的分子機制尚不清楚。
過氧化物酶體增殖物激活受體α(peroxisome proliferator-activated receptor α,PPARα)是核受體超家族成員,也是脂肪酸氧化酶基因的主要轉錄調控子。研究證實,與野生型小鼠相比,PPARα-/-小鼠心肌中SCAD蛋白表達顯著下調,且心肌對脂肪酸攝取、氧化能力下降,表明SCAD基因的表達受PPARα調控。已有大量的研究證明,心肌肥大過程伴隨著PPARα的表達下調[4-6]。我們的研究發現,PPARα受體激動劑非諾貝特可能通過上調SCAD的表達抑制大鼠心肌肥大,改善心肌能量代謝障礙[7-8]。
腺苷酸活化蛋白激酶(adenosine monophosphate-activated protein kinase, AMPK)是一種絲/蘇氨酸蛋白激酶,參與不同的生理及病理過程中細胞能量代謝的調控。研究表明,AMPK可通過激活PPARα信號通路,從而抑制心肌肥大[9]。因此,探討AMPK/PPARα/SCAD信號途徑是否對心肌肥大具有一定的調控作用,對于進一步闡明心肌肥大的分子機制具有重要的意義。
材 料 和 方 法
1 材料
單克隆兔抗p-AMPKα和AMPKα購于Cell Signaling Technology;單克隆鼠抗PPARα、單克隆兔抗SCAD、單克隆鼠抗α-tubulin、非諾貝特、5-bromo-2’-deoxyuridine(BrdU)、PE和5-氨基咪唑-4-甲酰胺核糖核苷酸(5-aminoimidazole-4-carboxamide ribonu-cleotide,AICAR)購于Sigma;BCA蛋白定量試劑盒和Western blotting發光液購于Thermo;游離脂肪酸含量測定試劑盒購于南京建成;RT-PCR測定試劑盒和TRIzol購于TaKaRa。
2 方法
2.1乳鼠心肌細胞培養 采用乳鼠心肌細胞改良法分離并培養心肌細胞,取2~3 d SD乳鼠(廣州中醫藥大學實驗動物中心)心臟用0.8%胰蛋白酶(Sigma)冰上消化20 min后,37 ℃水浴4 min,1次消化將乳鼠心臟消化成為單細胞懸液,在差速貼壁分離1 h后,調節細胞密度接種于培養皿中,置于37 ℃、5% CO2培養箱中培養,并加入BrdU (0.1 mmol/L)抑制成纖維細胞生長48 h。無血清培養24 h后,心肌細胞預先給予非諾貝特(10 μmol/L)或AICAR (0.5 mmol/L)30 min,再加入PE(20 μmol/L)刺激心肌細胞24 h,建立心肌細胞肥大模型。
2.2SCADRNA干擾SCADsiRNA購于上海吉瑪生物制藥技術有限公司,序列見表1。采用35 mm皿,siRNA∶Lipofectamine 2000 為 20 pmol∶1 μL。實驗分組如下: (1) 空白對照組(control,Con): 不加任何轉染試劑,不加siRNA;(2) 陰性對照組(negative control,NC): 加轉染試劑和陰性對照siRNA;(3) siRNA-1186轉染組:加轉染試劑和siRNA-1186;(4) siRNA-744轉染組:加轉染試劑和siRNA-744;(5) siRNA-207轉染組:加轉染試劑和siRNA-207。轉染方法按照公司提供的轉染試劑說明書進行。

表1 siRNA 序列
2.3SCAD活性測定 按上海杰美基因醫藥科技有限公司提供的說明書進行實驗,心肌細胞以(1~5)×106細胞密度接種于60 mm皿中,細胞生長密度達到70%~80%時以無血清培養基培養16~24 h 后,更換新的無血清培養基并且加入處理因素。
古樹名木是一種自然資源,也是一種文化遺產,具有重要的經濟、生態、科研、歷史、紀念等價值。古樹名木作為森林資源,可涵養水源、保持水土、除塵抑菌,具有穩固的生態功能[4];是珍貴的活文物,與城鎮和宗教文化發展息息相關,具有深刻的歷史文化價值[5];是自然科學研究的活標本,具有重要的科學價值[6];具有獨特的觀賞價值,是自然景觀和文化景觀的結合,是絕佳的旅游資源[7]。對古樹名木的現狀進行調查分析,并探討相應的保護策略具有重大理論和現實意義。
2.4心肌細胞表面積測定 PE刺激24 h后,用Zeiss(AX10)倒置熒光顯微鏡(×200)進行拍照,各處理組隨機選取10個視野,測量100個細胞的表面積,重復3次以上實驗,用ImageJ圖像分析系統進行數據分析。
2.5心肌細胞游離脂肪酸的含量測定 PE刺激24 h后,提取心肌細胞的總蛋白,采用BCA試劑盒進行蛋白定量,按照試劑盒說明書進行游離脂肪酸濃度的測定,根據吸光率計算心肌細胞中游離脂肪酸的含量。
2.6Real-time PCR檢測PPARα、SCAD、心房利鈉因子(atrial natriuretic factor,ANF)和腦利鈉肽(brain natriuretic peptide,BNP) mRNA的表達 按照TRIzol說明書步驟提取細胞總RNA,采用紫外分光光度計檢測RNA樣品的A260/A280比值在1.8~2.0之間,以確定RNA純度,并計算出RNA的濃度。參照RT-PCR試劑盒說明書進行逆轉錄反應,將反應管放置于PCR儀中進行逆轉錄反應,逆轉錄反應參數:95 ℃ 30 s (1個循環), 95 ℃ 5 s,60 ℃ 30 s(40個循環)。
RT-PCR擴增的引物序列見表2。兩步法進行PCR擴增反應,按照說明書加入SYBR熒光染料、引物序列(上海生工生物工程股份有限公司)和RT產物后在real-time PCR儀中進行反應。反應參數如下:95 ℃ 10 s,90 ℃ 5 s,循環50次;95 ℃ 15 s,60 ℃ 1 h,65 ℃ 30 s,循環61次。每組樣品設置3個復孔,以保證實驗結果的有效性和重復性。

表2 RT-PCR擴增的引物序列
2.7Western blotting檢測p-AMPKα、PPARα和SCAD蛋白的表達 提取各組心肌細胞總蛋白,BCA試劑盒檢測細胞蛋白含量后調整上樣量,分裝、變性,配置10% SDS分離膠和5%濃縮膠進行電泳,電泳結束后轉移至PVDF膜,室溫封閉2 h后加入Ⅰ抗(SCAD為1∶1 000; PPARα為1∶500; p-AMPKα為1∶1 000; AMPKα為1∶1 000;α-tubulin為1∶5 000),過夜。漂洗后加入Ⅱ抗,室溫孵育1 h,化學發光試劑增強反應,X光壓片曝光、顯影、定影,結果采用ImageJ圖像分析系統對條帶進行分析。
3 統計學處理
數據均以數±標準差(mean±SD)表示,采用SPSS 13.0 統計軟件處理,組間比較應用單因素方差分析,以P<0.05為差異有統計學意義。
結 果
1 siRNA的篩選結果
采用Western blotting、RT-PCR和SCAD活性測定比較3條干擾序列對SCAD的干擾效果,從圖1可見,與空白對照和陰性對照組相比較,siRNA-1186、siRNA-744和siRNA-207均可以不同的程度降低SCAD蛋白、mRNA表達和酶活性,其中以siRNA-1186在蛋白、mRNA和酶活性水平上的降低程度最明顯。因此選擇siRNA-1186進行后續實驗。

Figure 1. Selection of the siRNA sequences in cardiomyocytes.A: Western blotting results;B: RT-PCR results;C: SCAD activity.Mean±SD.n=5.*P<0.05,**P<0.01 vs Con.
2 siRNA-1186敲低SCAD基因對心肌細胞表面積及脂質代謝的影響
由圖2可見,siRNA-1186通過瞬時轉染對SCAD基因表達進行干擾后,心肌細胞表面積明顯增加,心肌細胞出現了明顯肥大,其程度與刺激因素PE誘導的心肌肥大趨勢一致。此外,心肌細胞對脂肪酸的氧化能力明顯降低,心肌細胞內游離脂肪酸含量顯著增多,SCAD下調對心肌細胞脂質代謝的影響與PE誘導心肌細胞肥大導致的脂質變化一致。這表明SCAD的表達下調可能導致了心肌細胞脂肪酸β氧化能力下降,從而引起心肌細胞的游離脂肪酸含量增加,導致心肌細胞肥大發生。
3 siRNA-1186敲低SCAD基因對心肌細胞目的基因表達與SCAD酶活性的影響
由圖3可見,siRNA-1186敲低SCAD基因和PE刺激均引起SCAD mRNA、蛋白表達及酶活性水平顯著下降。此外,心肌肥厚標志因子ANF和BNP mRNA表達均有明顯的上升,表明二者均引起心肌細胞出現了明顯肥大,SCAD在心肌肥大過程中具有重要作用,其表達量的降低可能是造成心肌肥大的一個重要因素。
4 非諾貝特對1186敲低SCAD基因的心肌細胞保護作用
從圖4可以看出,siRNA-1186通過瞬時轉染對SCAD基因表達進行干擾后,SCAD mRNA、蛋白及酶活性均明顯降低,心肌細胞表面積明顯增加,心肌肥厚標志因子ANF和BNP mRNA表達明顯上升,心肌細胞內游離脂肪酸含量明顯增加。與siRNA-1186干擾SCAD后的心肌細胞相比,非諾貝特預處理作用于敲低SCAD的心肌細胞后,心肌細胞表面積明顯變小,細胞內游離脂肪酸含量顯著減少,肥厚標志因子ANF和BNP mRNA表達有明顯的下降,SCAD mRNA水平明顯升高,相對應的蛋白表達量及SCAD酶活性也顯著增高,見圖4、5。這表明心肌肥大過程中PPARα對SCAD有直接的調控作用。

Figure 2. The changes of surface area (A,B) and content of free fatty acids (C) in the cardiomyocytes treated with phenylephrine (PE, 20 μmol/L) or siRNA-1186.Mean±SD.n=5. **P<0.01 vs Con.

Figure 3. The expression and activity of SCAD in cardiomyocytes treated with phenylephrine (PE, 20 μmol/L) or siRNA-1186. A: Western blotting results; B: RT-PCR results; C: SCAD activity.Mean±SD.n=5.##P<0.01 vs NC;**P<0.01 vs Con.

Figure 4. The effect of fenofibrate (Feno) on the surface area (A) and content of free fatty acids (B) in the cardiomyocytes treated with siRNA-1186.Mean±SD.n=5. **P<0.01 vs Con; #P<0.05, ##P<0.01 vs siRNA-1186.

Figure 5. The effect of fenofibrate (Feno) on the expression and the activity of SCAD in cardiomyocytes treated with siRNA-1186. A: Western blotting results; B: RT-PCR results; C: SCAD activity.Mean±SD.n=5.**P<0.01 vs Con; #P<0.05, ##P<0.01 vs riRNA-1186.
5 非諾貝特對PE處理的心肌細胞表面積及游離脂肪酸含量的影響
圖6顯示,PE刺激24 h后,心肌細胞表面積明顯增大,出現了心肌細胞肥大,而用非諾貝特能明顯預防心肌細胞肥大,心肌細胞表面積明顯減少。心肌細胞內的游離脂肪酸含量明顯增多,而非諾貝特能顯著降低心肌細胞的游離脂肪酸含量。

Figure 6. The effect of fenofibrate (Feno) on the surface area (A) and content of free fatty acids (B) in cardiomyocyte treated with phenylephrine (PE). Mean±SD.n=5.**P<0.01 vs Con; #P<0.05 vs PE.
6 非諾貝特對PE處理的心肌細胞PPARα和SCAD蛋白及mRNA表達的影響
由圖7可見,PE刺激24 h后,心肌細胞內肥大標志物ANF和BNP mRNA表達量均顯著上升,說明心肌細胞肥大模型建立成功。而SCAD和PPARα蛋白及mRNA表達水平均顯著下調,表明心肌細胞肥大模型中,SCAD的表達下調,可能導致了心肌細胞的脂肪酸β氧化能力下降,從而引起心肌細胞的游離脂肪酸含量增加。非諾貝特能使PPARα和SCAD蛋白及mRNA水平顯著上調,表明非諾貝特可能通過增加SCAD的表達,從而導致心肌細胞脂肪酸β氧化能力增強,引起心肌細胞的游離脂肪酸含量下降,預防心肌細胞肥大。
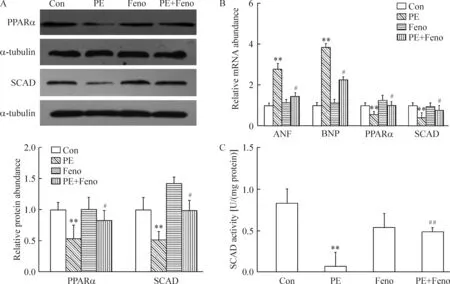
Figure 7. The effect of fenofibrate (Feno) on the expression of PPARα and SCAD at mRNA and protein levels in the cardiomyocytes treated with phenylephrine (PE).A: Western blotting results; B: RT-PCR results; C: SCAD activity.Mean±SD.n=5. **P<0.01 vs Con; #P<0.05,##P<0.01 vs PE.
7 AICAR對PE處理的心肌細胞表面積及游離脂肪酸含量的影響
由圖8可見,PE刺激24 h后,心肌細胞表面積明顯增大,而AICAR能顯著逆轉心肌細胞肥大;心肌細胞內的游離脂肪酸含量明顯增多,而AICAR能顯著降低心肌細胞的游離脂肪酸含量,這一變化趨勢與非諾貝特預處理相一致。
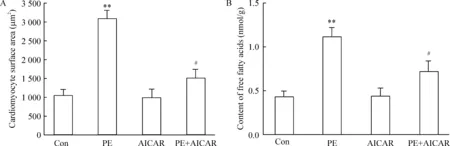
Figure 8. The effect of 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) on the surface area (A) and content of free fatty acids (B) in the cardiomyocytes treated with phenylephrine (PE).Mean±SD.n=5. **P<0.01 vs Con; #P<0.05 vs PE.
8 AICAR對PE處理的心肌細胞p-AMPKα、PPARα及SCAD表達的影響
由圖9可見,PE刺激24 h后,心肌細胞ANF和BNP mRNA表達量均顯著上升,而SCAD和PPARα mRNA表達則有所下調。與mRNA的變化趨勢一致的是,PE刺激24 h后,SCAD、PPARα和p-AMPKα蛋白水平亦顯著下調。AICAR能顯著激活AMPK,使p-AMPKα表達下調。同時, PPARα和SCAD蛋白和mRNA水平均顯著上調,表明AICAR可能通過激活AMPK增加PPARα的表達,從而上調下游基因SCAD,導致心肌細胞的脂肪酸β氧化能力增強,引起心肌細胞的游離脂肪酸含量下降,預防心肌細胞肥大。

Figure 9. The effect of 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) on the protein and mRNA expression of p-AMPKα, PPARα and SCAD in the cardiomyocytes treated with phenylephrine (PE).A: Western blotting results; B: RT-PCR results; C: SCAD activity. Mean±SD.n=5.**P<0.01 vs Con; #P<0.05,##P<0.01 vs PE.
討 論
心肌肥大是心肌細胞對高血壓、瓣膜病、急性心肌梗死及先天性心臟病等常見臨床疾病的一種基本應答。心肌肥大的病理轉變過程可分為肥大進展、肥大代償和肥大失代償期3個階段。引起心肌肥大的發生機制極其復雜,至今仍未完全明了[10]。
心肌肥大是肥大刺激誘導核內基因異常表達的結果,細胞內信號轉導通路是肥大刺激與核內基因轉錄活化的偶聯環節。然而,不同刺激誘導的心肌肥大可能具有不同的“分子表型”,這主要取決于它們啟動的信號轉導通路。對心肌肥大信號轉導通路的深入認識,不僅有助于闡明心肌肥大的分子機制,而且可為藥物干預防治心肌肥大提供新思路[11]。
研究證實,胚胎時期哺乳動物心肌處于相對缺氧的環境,以葡萄糖及丙酮酸作為主要的能源物質,出生后則轉為依賴脂肪酸氧化[12]。大量證據表明:當病理因素導致心肌肥大時,心肌能源供給由脂肪酸β氧化轉變為以葡萄糖的酵解為主,即心肌能量代謝的“胚胎型再演”[13]。而與脂肪酸β氧化密切相關的SCAD是高度保守的酰基輔酶A脫氫酶家族成員之一,是脂肪酸β氧化第一步反應的限速酶。SCAD蛋白表達隨年齡變化逐漸增加,且血清和心肌游離脂肪酸含量逐漸減少,可能與大鼠心肌對能量代謝底物的利用轉變有關[2]。因此,SCAD作為脂肪酸β氧化的關鍵酶,其表達變化與心臟生長發育密切相關的同時,對心臟的能量代謝也起到重要的作用,進一步研究SCAD在心肌肥大能量代謝中的機制有著重要的意義。
本研究進一步采用RNA干擾技術敲低SCAD的表達,觀察SCAD與心肌細胞肥大的關系,并與PE誘導的心肌細胞肥大模型相比較。我們觀察到,敲低SCAD的表達后,心肌細胞表面積明顯增大,心肌肥大標志因子ANF和BNP mRNA水平均顯著上升,與PE誘導的心肌肥大趨勢一致,SCAD的表達下調直接導致了心肌細胞的肥大發生,表明SCAD在心肌肥大的發生中具有重要作用。
敲低SCAD的表達后,心肌細胞內SCAD的蛋白表達和酶活性明顯降低,心肌細胞內游離脂肪酸含量顯著升高,提示心肌細胞的能量代謝出現了明顯改變。SCAD下調對心肌細胞脂質代謝的影響與PE誘導心肌細胞肥大導致的脂質變化一致。此外,PE誘導的心肌肥大模型中,SCAD mRNA、蛋白表達及酶活性均顯著下降。因此,我們推測,SCAD的脂肪酸氧化作用顯著下降,可能導致了心肌細胞對脂肪酸的攝取和氧化能力明顯降低,從而引起心肌細胞游離脂肪酸含量不斷增多。
PPARα是核受體超家族成員,主要分布在脂肪酸氧化效率高的組織,是生理條件下心肌脂質和能量代謝的重要調控因子。然而,包括SCAD在內,依賴PPARα的脂肪酸代謝酶至少有7個。因此,心肌中高分布的PPARα與心肌能量代謝密切相關。本研究發現PE刺激24 h后,心肌細胞明顯肥大,心肌細胞的游離脂肪酸含量明顯增加,并伴有PPARα和SCAD的表達下調,呈現脂肪酸的利用減少和SCAD的表達下調相一致的趨勢,表明肥大心肌存在脂肪酸氧化酶基因表達水平的降低,從而引起脂肪酸利用受阻,最終導致心肌能量代謝障礙,可能與肥大心肌細胞中PPARα的表達水平降低,引起的SCAD表達減少有關。本研究采用PPARα受體激動劑,觀察到非諾貝特在激活PPARα和SCAD表達的同時,可明顯逆轉心肌細胞肥大,增強肥大心肌細胞對脂肪酸的代謝,降低心肌細胞中游離脂肪酸的含量。并在證實SCAD下調與心肌肥大脂質代謝密切相關的基礎上,觀察到非諾貝特預處理可以上調SCAD mRNA、蛋白表達及增高酶的活性,減少心肌細胞表面積,降低肥厚標志因子ANF和BNP mRNA表達,減低心肌細胞內游離脂肪酸的含量,直接預防敲低SCAD引起的心肌細胞肥大,表明心肌肥大中PPARα對SCAD具有直接的調控作用。綜上所述,非諾貝特可能通過激活心肌細胞PPARα,進而上調依賴PPARα的下游基因SCAD,使SCAD酶活性增強,從而增強心肌細胞脂肪酸的β氧化,改善心肌細胞的能量代謝,預防心肌細胞肥大。這表明PPARα/SCAD信號途徑對心肌肥大具有直接的調控作用。
AMPK是一個表達在心血管系統的異源三聚體蛋白,能調節體內能量代謝,在壓力超負荷的作用下,會增加細胞AMP/ATP的比率,導致ATP減少。AMPK失活與心肌功能障礙及其它代謝性疾病相關[14], AICAR為AMPK的激活劑,可通過激活AMPKα磷酸化位點Thr172,對PE誘導的心肌肥大有一定的保護作用[15]。本研究顯示,PE刺激24 h后,p-AMPK的表達顯著下調,AICAR在激活AMPK的同時,可明顯逆轉心肌細胞肥大,顯著增強肥大心肌細胞的脂肪酸氧化,并伴隨著PPARα及SCAD的表達上調,表明AICAR可能通過激活AMPK進而活化PPARα,上調脂肪酸氧化限速酶SCAD的表達,從而增強心肌細胞脂肪酸的β氧化,改善能量代謝,預防心肌細胞肥大。可見,AMPK/PPARα/SCAD信號途徑可能介導了心肌細胞肥大的發生。
研究表明,體外激活AMPK抑制心肌肥大是通過FOXO1/MuRF1信號通路介導的[16]。而AICAR可通過激活ERK1/2信號途徑,從而提高PPARα活性,抑制心肌肥大[9]。可見,激活AMPK/PPARα信號通路可能與AMPK/FOXO1/MuRF1信號通路有關,ERK1/2信號途徑是否參與了AMPK/PPARα/SCAD信號通路對心肌肥大的調控,有待進一步研究。
[參 考 文 獻]
[1] Zhou SG, Zhou SF, Huang HQ, et al. Proteomic analysis of hypertrophied myocardial protein patterns in renovascularly hypertensive and spontaneously hypertensive rats[J]. J Proteome Res, 2006, 5(11): 2901-2908.
[2] 周四桂, 王 平, 路 遙, 等. 短鏈酰基輔酶A脫氫酶在大鼠心臟發育中的表達及其與心肌肥厚的關系[J]. 中國病理生理雜志, 2013, 29(1):9-14.
[3] 羅佳妮, 周四桂, 陳少銳, 等. 短鏈脂酰輔酶A脫氫酶與心肌肥大關系的初步探索[J]. 中國藥理學通報, 2013, 29(5):637-642.
[4] Ichihara S, Obata K, Yamada Y, et al. Attenuation of cardiac dysfunction by a PPAR-α agonist is associated with down-regulation of redox-regulated transcription factors[J]. J Mol Cell Cardiol, 2006, 41(2):318-329.
[5] Zou J, Le K, Xu S, et al. Fenofibrate ameliorates cardiac hypertrophy by activation of peroxisome proliferator-activated receptor-α partly via preventing p65-NFκB binging to NFATc4[J]. Mol Cell Endocrinol, 2013, 370(1-2):103-112.
[6] Purushothaman S, Sathik MM, Nair RR. Reactivation of peroxisome proliferator-activated receptor alpha in spontaneously hypertensive rat: age-associated paradoxical effect on the heart[J]. J Cardiovasc Phamacol, 2011, 58(3):254-262.
[7] 周四桂, 王 平, 路 遙, 等. 短鏈酰基輔酶A脫氫酶在大鼠生理性和病理性心肌肥大中的作用[J]. 中國病理生理雜志, 2012, 28(11):1921-1927。
[8] 周四桂, 徐立朋, 路 遙, 等. 非諾貝特通過上調短鏈酰基輔酶A脫氫酶抑制大鼠心肌肥厚[J]. 中國臨床藥理學與治療學, 2013, 18(8):868-873.
[9] Meng R, Pei Z, Zhang A, et al. AMPK activation enhances PPARα activity to inhibit cardiac hypertrophy via ERK1/2 MAPK signaling pathway[J]. Arch Biochem Biophy, 2011, 511(1-2):1-7.
[10] 李 潞. 心臟肥大發生機制研究進展[J]. 沈陽醫學院學報, 2009, 11(1):1-5.
[11] 符民桂, 唐朝樞. 心肌細胞肥大的信號轉導通路[J]. 生理科學進展, 2000, 31(1):19-24.
[12] Khan RS, Lin Y, Hu Y, et al. Rescue of heart lipoprotein lipase knockout mice confirms a role for triglyceride in optimal heart metabolism and function[J]. Am J Physiol Endocrinol Metab, 2013, 305(11): E1339-E1347.
[13] Breckenridge RA, Piotrowska I, Ng KE, et al. Hypoxic regulation ofHand1 controls the fetal-neonatal switch in cardiac metabolism[J]. PLoS Biol, 2013, 11(9): e1001666.
[14] Wong AK, Howie J, Petrie JR, et al. AMP-activated protein kinase pathway a potential therapeutic target in cardiometabolic disease[J]. Clin Sci, 2009, 116(8): 607-620.
[15] Pang T, Raiapurohitam V, Cook M, et al. Differential AMPK phosphorylation sites associated with phenylephrinevsantihypertrophic effects of adenosine agonists in neonatal rat ventricular myocytes[J]. Am J Physiol Heart Circ Physiol, 2010, 298(5):H1382-H1390.
[16] Chen BL, Ma YD, Meng RS, et al. Activation of AMPK inhibits cardiomyocyte hypertrophy by modulating of the FOXO1/MuRF1 signaling pathyinvitro[J]. Acta Pharmacol Sin, 2010, 31(7):798-804.
