丙型肝炎病毒HLA-A*1101和A*2402限制性CD8+T細胞表位的研究
2014-08-08 09:41:12鐘曉芝段志良郭江龍李強王思娜李靜林榮文金生
溫州醫(yī)科大學(xué)學(xué)報 2014年8期
關(guān)鍵詞:研究
鐘曉芝,段志良,郭江龍,李強,王思娜,李靜,林榮,文金生
丙型肝炎病毒(hepatitis C virus,HCV)為有 包膜的單股正鏈RNA病毒,其基因組編碼三個結(jié)構(gòu)蛋 白(C、E1和E2)和七個非結(jié)構(gòu)蛋白(P1、NS2、NS3、NS4a、NS4b、NS5a和NS5b)[1]。HCV感染肝臟后引起急性丙型肝炎,可發(fā)展成慢性肝炎、肝硬化,甚至肝細胞癌。目前全球已有約1.7億丙型肝炎患者[2]。由于臨床上丙型肝炎治療效果不太理想,因而,研發(fā)有效的疫苗仍是目前HCV研究的重點和熱點。
有研究表明:HCV特異性抗體無保護作用,而活化的CD8+T細胞是控制HCV感染和清除細胞內(nèi)感染病毒的決定因素[3-4]。因此,能誘導(dǎo)特異性CD8+T細胞反應(yīng)的CD8+T細胞表位肽疫苗是一種很有希望的候選疫苗。HLA-A的三種等位基因HLA-A*0201、A*1101和A*2402在不同種族人群中總百分率均大于90%[5],因此由三種等位基因限制性表位組成的CD8+T細胞表位肽疫苗具有廣泛的人群覆蓋率。在前期研究中,我們課題組以及國外研究人員已鑒定出了大量HCV特異性HLA-A*0201限制性CD8+T細胞表位[6-9],但HLA-A*1101及A*2402限制性細胞表位的報道則寥寥無幾。本研究擬鑒定HCV特異性HLA-A*1101和A*2402限制性CD8+T細胞表位,為研發(fā)具有廣泛人群覆蓋性的CD8+T細胞表位肽疫苗奠定基礎(chǔ)。
1 材料和方法
1.1 材料 未感染HCV的健康志愿者招募自溫州醫(yī)科大學(xué)在校研究生,HCV基因型1b感染者血液由瑞安市人民醫(yī)院檢驗科及溫州市中心血站提供,人淋巴細胞分離液購自天津市灝洋生物制品科技有限責(zé)任公司,人γ-干擾素(IFN-γ)酶聯(lián)免疫斑點(ELISPOT)試劑盒購自荷蘭U-CyTech公司,APC標記抗人CD3、FITC標記抗人CD8a和PE標記抗人IFN-γ的流式單抗購自美國Biolegend公司,ELISPOT分析儀來自北京賽智創(chuàng)業(yè)科技有限公司,F(xiàn)ACS Calibur流式細胞儀來自美國BD bioscience公司。
1.2 方法
1.2.1 候選CD8+T細胞表位的預(yù)測及其合成:基于HCV基因型1b(AF165064)的氨基酸序列,采用T細胞表位預(yù)測軟件SYFPEITHI(http://www.syfpeithi.de/Scripts/MHCServer.dll/EpitopePrediction.htm)預(yù)測HCV特異性HLA-A*1101及A*2402限制性候選CD8+T細胞表位[10-11]。選擇候選表位的標準如下:①候選表位具有較高的預(yù)測得分;②通過Blast檢索,候選表位在1b基因型中高度保守;③候選表位C-末端殘基是蛋白酶體裂解預(yù)測結(jié)果。FITC標記的已報道HLA-A*1101限制性表位肽NS4b_147(GVAGALVAFK)[12]和FITC標記的已報道HLA-A*2402限制性表位肽NS4b_49(FWAKHMWNF)[13]用作陽性表位肽。委托上海強耀生物科技有限公司合成候選表位肽及陽性表位肽(純度>95%)。肽溶于PBS(1 mg/mL),儲存于-80 ℃。
1.2.2 永生化的HLA-A*1101陽性、A*2402陽性B細胞株的建立:設(shè)計擴增HLA-A*1101和A*2402的引物。在健康志愿者簽署了知情同意書后,抽取每人1 mL EDTA抗凝的外周靜脈血,采用DNA提取試劑盒提取DNA。基于上述引物,采用PCR儀擴增HLA-A*1101和 A*2402,PCR產(chǎn)物采用2%瓊脂糖凝膠電泳。采集HLAA*1101陽性、A*2402陽性者的10 mL EDTA抗凝外周靜脈血,采用人淋巴細胞分離液分離外周血單個核細胞(PBMCs),將PBMCs懸浮于RPMI-1640培養(yǎng)基中并置于細胞培養(yǎng)板孔中。傳代培養(yǎng)B95-8細胞(感染EB病毒的絨猴白細胞,其培養(yǎng)上清中含有大量具有轉(zhuǎn)化B細胞作用的EB病毒基因),將其培養(yǎng)上清加入PBMCs中誘導(dǎo)B細胞永生化[14]。
1.2.3 競爭性肽結(jié)合實驗:大量培養(yǎng)上述永生化的 HLA-A*1101陽性、A*2402陽性B細胞,收集細胞。用 枸櫞酸(0.131 mol/L枸櫞酸和0.061 mol/L磷酸氫 二鈉,pH 3.2)處理細胞90 s以洗脫表面HLA-A*1101、 A*2402分子肽結(jié)合凹槽中結(jié)合的肽[15]。細胞經(jīng)PBS洗滌后分成以下管:陰性對照管(無肽),細胞在無候選表位肽和陽性表位肽的情況下于37 ℃孵育 6 h;實驗管,細胞中加入不同濃度(1μg/mL、10 μg/mL、100μg/mL)的候選表位肽和FITC標記的HLA-A*1101限制性或A*2402限制性陽性表位肽(NS4b_147或NS4b_49,均為30μg/mL),于37 ℃孵 育6 h;陽性對照管,細胞同F(xiàn)ITC標記陽性表位肽 (NS4b_147或NS4b_49,均為30μg/mL)于37 ℃孵育 6 h。采用流式細胞儀測定細胞平均熒光強度(MFI)。下述公式用于計算候選表位肽對陽性表位肽的百分抑制率(percentage inhibition,PI)。PI>50%表示候選表位肽與HLA-A*1101或A*2402有高結(jié)合力。
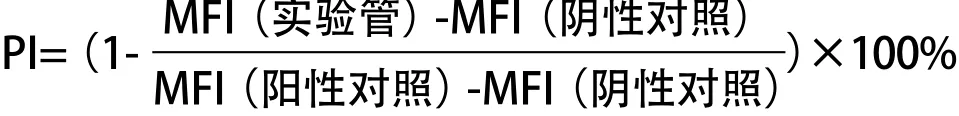
1.2.4 ELISPOT檢測HCV感染者PBMCs中肽特異性分泌IFN-γ的細胞:6例HLA-A*1101陽性HCV-1b感染者和6例HLA-A*2402陽性HCV-1b感染者(ELISA-IgM試劑盒證實為HCV感染,HCV基因型特異性RT-PCR證實為HCV基因型1b感染[16])來自瑞安市人民醫(yī)院檢驗科及溫州市中心血站,在研究對象簽署了知情同意書后,抽取每人10 mL EDTA抗凝的外周靜脈血。采用人淋巴細胞分離液分離PBMCs,用RPMI-1640培養(yǎng)基(含2 mmol/L L-谷氨酰胺,100 U/mL青霉素,100μg/mL鏈霉素和10%胎牛血清)懸浮PBMCs。ELISPOT實驗簡單過程如下:將1×105PBMCs(懸浮 于100 μL RPMI-1640培養(yǎng)液)加入人IFN-γ ELISPOT試劑盒(預(yù)包被的PVDF膜96孔板)每孔中,設(shè)置陰性對照孔(加PBMCs而不加多肽),多肽刺激孔(加PBMCs同時加入多肽,其終濃度為10μg/mL),每個處理因素均設(shè)置2孔。將ELISPOT板放于37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)24 h。棄去每孔細胞及液體,洗板4次后,每孔加入100μL生物素化抗IFN-γ的檢測抗體,于37 ℃培養(yǎng)1 h。棄去孔中液體,洗板4次后,每孔加入100μL辣根過氧化物酶(HRP)標記的鏈酶親和素,于37 ℃培養(yǎng)1 h(避光)。棄去孔中液體,洗滌4次后,每孔加入100μL現(xiàn)配的ACE顯色液,于37 ℃反應(yīng)15~20 min(避光)。移除每孔中液體,用去離子水沖洗板。采用ELISPOT斑點分析儀對每孔斑點計數(shù),一個斑點代表一個斑點形成細胞(SFC),斑點數(shù)表示為SFCs/1×105PBMCs。減去陰性對照孔斑點數(shù)后,肽刺激孔斑點數(shù)≥5 SFCs/1×105PBMCs時,結(jié)果判讀為陽性[17]。
1.2.5 細胞內(nèi)細胞因子染色(ICS)檢測HCV感染者PBMCs中肽特異性IFN-γ+CD8+T細胞:5×105PBMCs(懸浮于500μL RPMI-1640營養(yǎng)液)加入EP管中,設(shè)置陰性對照管(無肽)和肽刺激管(多肽的終濃度為10μg/mL)。細胞管中加入布雷菲德菌素A(BFA)使其達到終濃度為10μg/mL,于37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)6 h。PBS洗滌細胞后,采用4%的多聚甲醛于4 ℃固定20 min。PBS洗滌細胞后,采用1%皂素懸浮細胞,然后加入APC標記抗人CD3、FITC標記抗人CD8a和PE標記抗人IFN-γ的單抗,避光于4 ℃孵育40 min。PBS洗滌細胞后,采用流式細胞儀檢測CD8+T細胞中IFN-γ+CD8+T細胞的百分率。
1.3 統(tǒng)計學(xué)處理方法 采用SPSS 17.0軟件統(tǒng)計。數(shù)據(jù)用±s來表示,組間比較采用t檢驗。P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié)果
2.1 候選CD8+T細胞表位與HLA-A*1101、A*2402分子的結(jié)合實驗 根據(jù)SYEPEITHI預(yù)測結(jié)果,我們選擇了5條HLA-A*1101限制性候選表位[E1_61(SIPTTTIRR)、 NS3_609(ITLTHPITK)、NS2_165(VVFSDMETK)、NS3_ 351(GKAIPLEVIK)和NS5b_251(QVIRSLTER)]和3條HLA-A*2402限制性候選表位[NS3_373(KCDELASKL)、NS5b_382(YYLTRDPTI)和NS5b_451(TYSIEPLDL)]并委托多肽合成公司加以合成。液相色譜和質(zhì)譜結(jié)果均證實合成多肽為目的肽,肽的實際純度均>95%。候選表位肽和陽性表位肽的詳細信息見表1。
2.2 永生化的HLA-A*1101陽性、A*2402陽性B細胞株的建立 擴增HLA-A*1101和A*2402的引物見表2。擴增A*1101的條件為:94 ℃ 4 min,30循環(huán)(94 ℃ 30 s,60 ℃ 1 min,72 ℃ 1 min),72 ℃ 4 min。擴增A*2402的條件為:94 ℃ 4 min,15循環(huán)(94 ℃ 30 s,65 ℃ 1 min),30循環(huán)(94 ℃ 30 s,55 ℃ 50 s,72 ℃ 30 s),72 ℃ 5 min。A*1101和A*2402 的PCR產(chǎn)物大小分別為500 bp和557 bp,見圖1。圖2和圖3分別為HLA-A*1101和HLA-A*2402的PCR產(chǎn)物的測序結(jié)果,Blasting檢索結(jié)果證實PCR產(chǎn)物序列與實際序列完全一致。圖4為PBMCs和永生化的B細胞株的圖片。PBMCs的細胞偏小,邊緣整齊,折光度好,無分裂生長能力。隨著B95-8細胞培養(yǎng)上清刺激PBMCs的時間不斷延長,細胞量減少,存活細胞的邊緣不整齊,細胞變大并呈現(xiàn)分裂生長趨勢。最后,細胞分裂迅速,呈團塊狀生長,經(jīng)過進一步傳代培養(yǎng)后即建立了穩(wěn)定的永生化的B細胞株。最終,我們建立了HLA-A*1101陽性、A*2402陽性B細胞株。
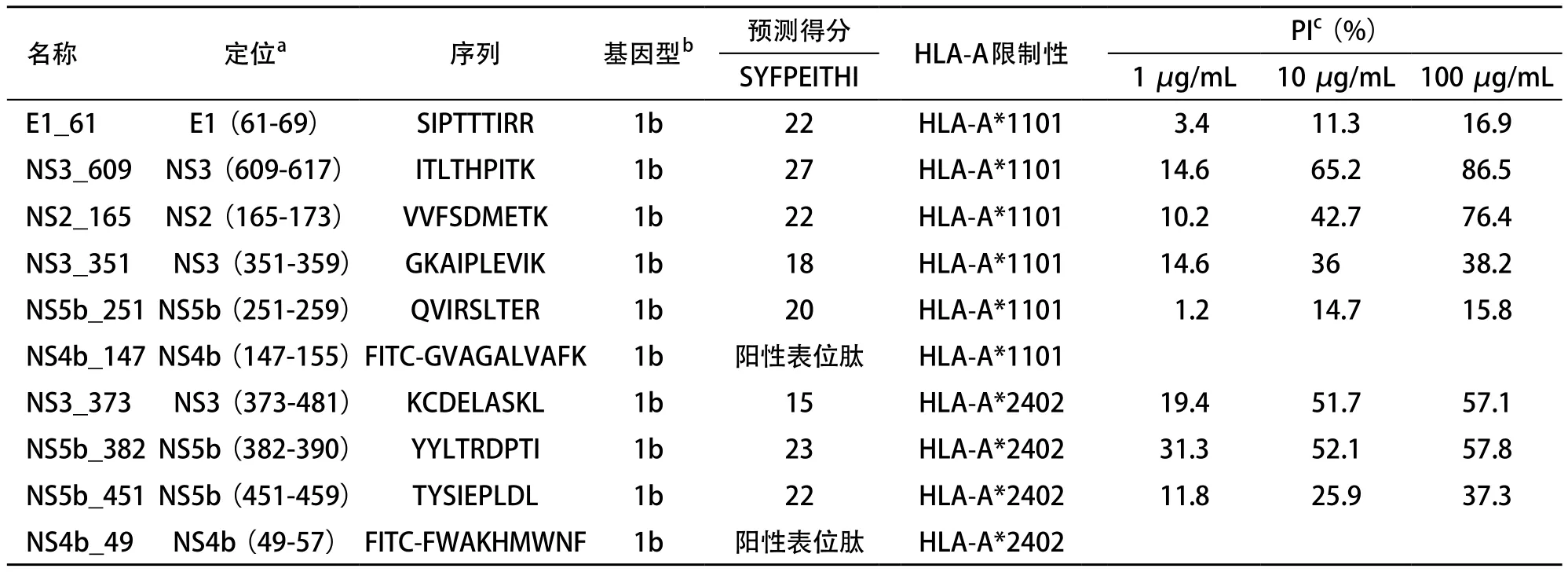
表1 HCV特異性HLA-A*1101、A*2402限制性候選CD8+T細胞表位肽及陽性表位肽
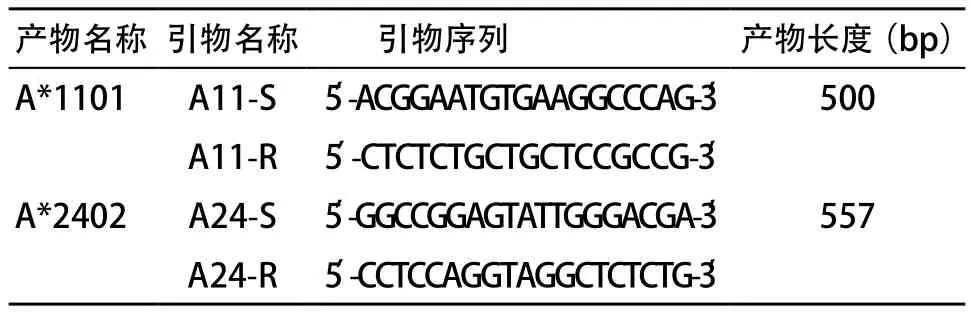
表2 擴增HLA-A*1101和A*2402的引物
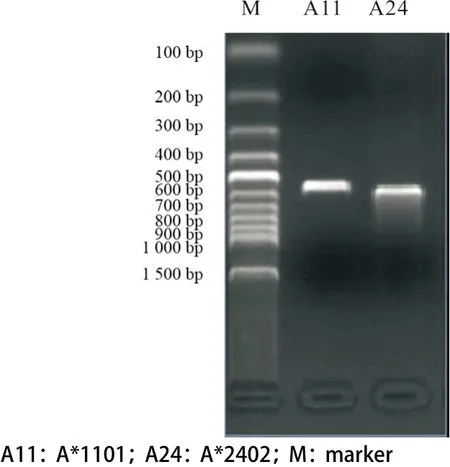
圖1 HLA-A*1101和HLA-A*2402的PCR擴增結(jié)果
2.3 競爭性肽結(jié)合實驗結(jié)果 基于上述建立的永生化的B細胞株,我們研究了候選表位肽與HLA分子的結(jié)合力。在5條HLA-A*1101限制性候選表位肽(E1_61、NS3_609、NS2_165、NS3_351和NS5b_251)中,NS3_609和NS2_165對陽性表位肽的PI與候選表位肽的濃度呈正比,當肽濃度為100μg/mL時,NS3_609和NS2_165的PI均大于50%(分別為86.5%和76.4%)(見表1和圖5)。在3條HLA-A*2402限制性候選表位肽(NS3_373、NS5b_382和NS5b_451)中,NS3_373和NS5b_382對陽性表位肽的PI與候選表位肽的濃度呈正比,當肽濃度為100μg/mL時,NS3_373和NS5b_382的PI均大于50%(分別為57.1%和57.8%)(見表1和圖6)。
圖2 HLA-A*1101的PCR產(chǎn)物測序結(jié)果
圖3 HLA-A*2402的PCR產(chǎn)物測序結(jié)果

圖4 正常的PBMCs和B95-8細胞上清刺激HLA-A*1101陽性PBMCs產(chǎn)生的永生化B細胞株
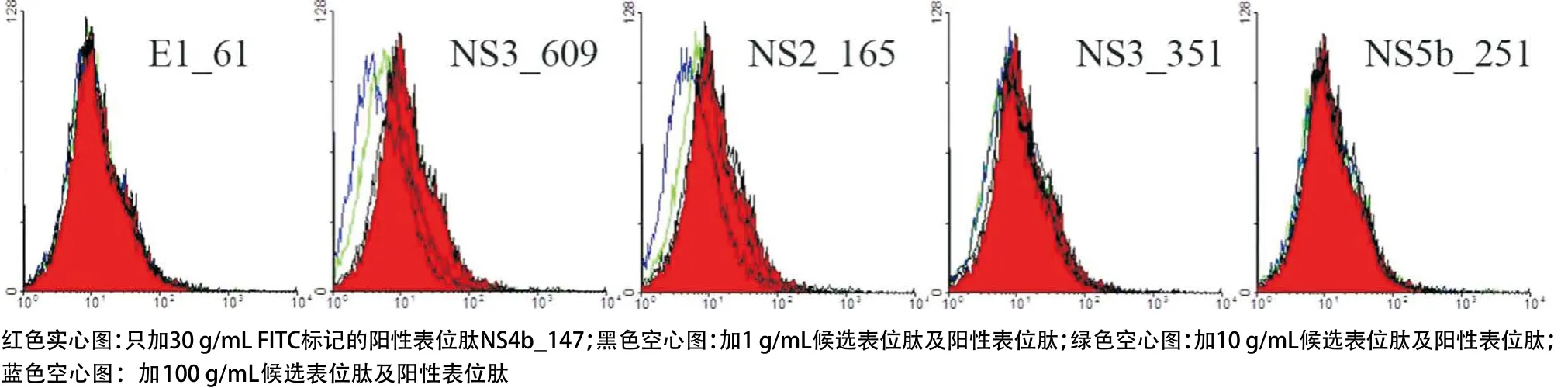
圖5 候選表位與HLA-A*1101分子的結(jié)合力
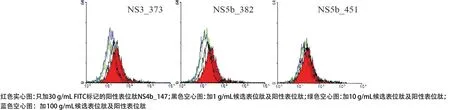
圖6 候選表位與HLA-A*2402分子的結(jié)合力
2.4 IFN-γELISPOT實驗 在6名HLA-A*1101陽性HCV感染者PBMCs中,NS3_609和NS2_165特異性分泌IFN-γ的細胞頻率分別為(14±4)SFCs/1×105PBMCs和(8±2)SFCs/1×105PBMCs(已減去陰性對照),均大于5 SFCs/1×105PBMCs;而在6名HLA-A*2402陽性HCV感染者PBMCs中,NS3_373和NS5b_382特異性分泌IFN-γ的細胞頻率分別為(12±5)SFCs/1×105PBMCs和(15±4)SFCs/1×105PBMCs(已減去陰性對照),均大于5 SFCs/1×105PBMCs;其他肽特異性IFN-γ的斑點數(shù)在減去陰性對照后均小于5 SFCs/1× 105PBMCs(見圖7)。
2.5 PBMCs中肽特異性IFN-γ+CD8+T細胞的水平 在6名HLA-A*1101陽性HCV感染者PBMCs中,NS3_609和NS2_165特異性IFN-γ+CD8+T細胞占總CD8+T細胞的百分率分別為0.12%±0.04%和0.06%±0.02%,與陰性對照比差異有統(tǒng)計學(xué)意義(P<0.05);而在6名HLA-A*2402陽性HCV感染者PBMCs中,NS3_373和NS5b_382特異性IFN-γ+CD8+T細胞占總CD8+T細胞的百分率分別為0.11%±0.03%和0.15%±0.05%,與陰性對照比較差異有統(tǒng)計學(xué)意義(P<0.05);其他肽特異性IFN-γ+CD8+T細胞的百分率與陰性對照比較差異無統(tǒng)計學(xué)意義(P>0.05)(見圖8)。
3 討論
大量研究證實:HCV感染機體產(chǎn)生的抗體對HCV沒有中和作用,而誘導(dǎo)的CD8+T細胞可清除感染者體內(nèi)HCV[3-4,18-23]。由于天然HCV抗原含有不合理的表位組分,能夠誘導(dǎo)Th1/Th2平衡改變,免疫偏差或免疫顛覆,而使HCV逃避機體的免疫防御而持續(xù)存在于肝細胞內(nèi),導(dǎo)致感染慢性化、肝硬化或肝癌。因而,僅能誘發(fā)CD8+T細胞反應(yīng)的CD8+T細胞表位疫苗已成為目前HCV疫苗研究的新方向[24-25]。HLAA的常見等位基因A*0201、A*1101和A*2402在不同種族人群中具有極高的總百分率。在既往研究中,基于T2細胞株(表達HLA-A*0201分子,其肽結(jié)合凹槽中無肽),研究者們已鑒定出多條HLA-A*0201限制性CD8+T細胞表位[6-9]。然而,由于缺乏類似的細胞株,HLA-A*1101和A*2402限制性CD8+T細胞表位的鑒定受到了極大制約。

圖7 HCV感染者PBMCs中肽特異性分泌IFN-γ的細胞的水平
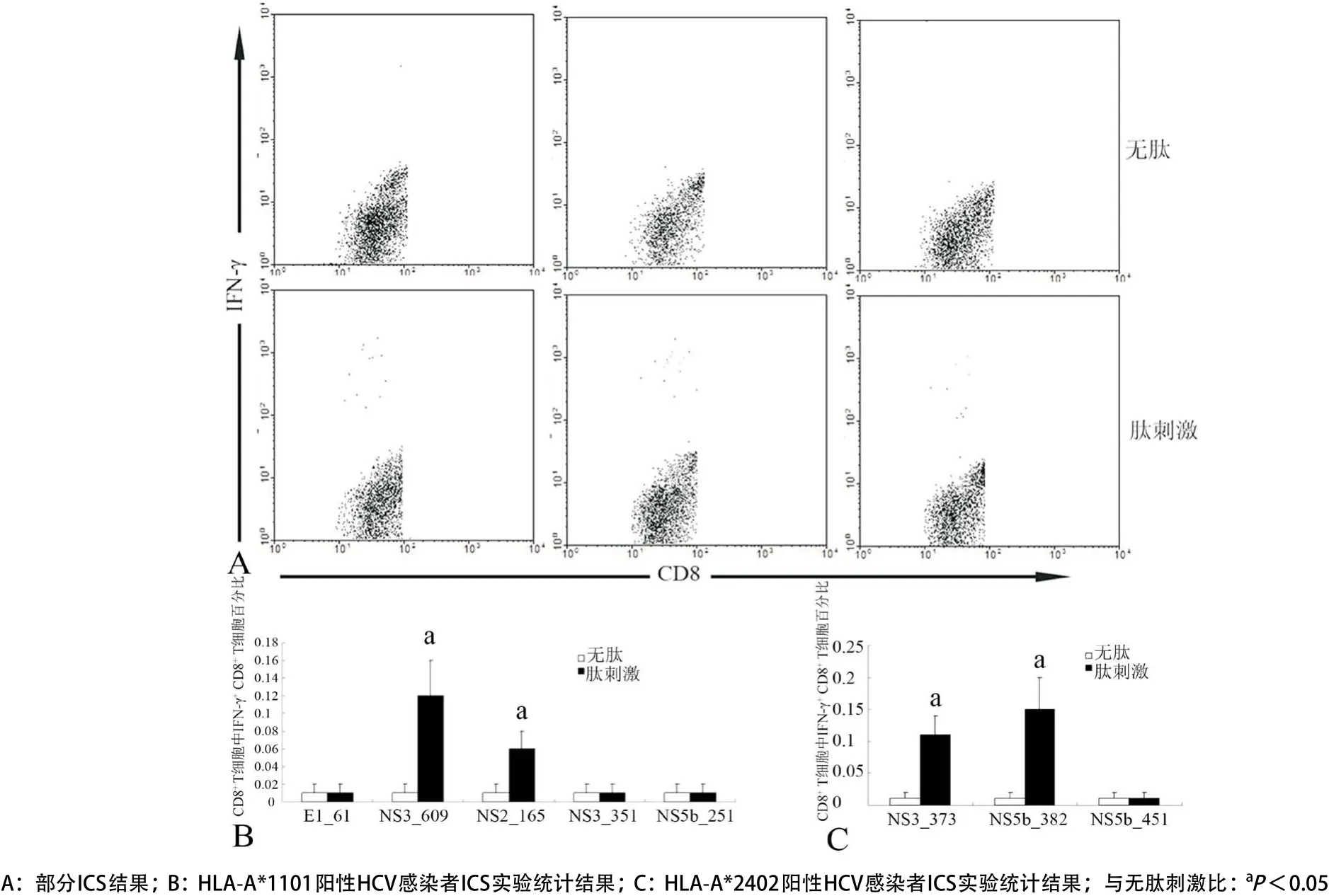
圖8 HCV感染者CD8+T細胞中肽特異性IFN-γ+CD8+T 細胞的百分比
我們前期在登革病毒和HCV中的研究表明:T細胞表位預(yù)測軟件預(yù)測結(jié)合體內(nèi)、外實驗驗證法是一種快捷、費用低廉的鑒定病毒T細胞表位的研究策略[8,26-27]。由于HCV-1b基因型感染者在HCV感染者中占極高比率[16],在本研究中,我們基于1b基因型的氨基酸序列采用SYFPEITHI預(yù)測合成了5條HLAA*1101限制性候選表位肽和3條HLA-A*2402限制性候選表位肽。研究表明:低濃度的枸櫞酸可以洗脫細胞表面HLA分子肽結(jié)合凹槽中的肽,為后續(xù)研究候選表位肽與HLA的結(jié)合奠定了物質(zhì)基礎(chǔ)[15]。基于此,競爭性肽結(jié)合實驗已成功用于檢測候選表位肽與HLA的結(jié)合力[15]。其基本原理為:首先采用枸櫞酸處理細胞(PBMCs或細胞株),然后將候選表位肽和FITC標記的已報道的能與某種HLA等位基因分子結(jié)合的陽性表位肽同枸櫞酸處理的細胞共同孵育,最后采用流式細胞儀檢測細胞MFI并計算PI。候選表位肽與HLA-A*1101或A*2402結(jié)合力越強,則占據(jù)的HLA-A分子越多,而熒光素標記的陽性表位肽結(jié)合的量就越少,因而細胞的MFI就越弱,PI則越高。因而,PI可用于反映候選表位肽與HLA分子的結(jié)合力。相對PBMCs而言,細胞表型單一并能穩(wěn)定表達HLA分子的細胞株能產(chǎn)生更穩(wěn)定和可靠的實驗結(jié)果。B95-8細胞是一種感染EB病毒的絨猴白細胞,實驗證實其培養(yǎng)上清可誘導(dǎo)某些B細胞永生化產(chǎn)生穩(wěn)定的細胞株[14]。在本研究中,我們招募了近50例健康志愿者,最后建立了穩(wěn)定的HLA-A*1101陽性、HLA-A*2402陽性永生化B細胞株。基于建立的B細胞株,競爭性肽結(jié)合實驗結(jié)果證明了候選表位肽NS3_609和NS2_165與HLA-A*1101分子具有高結(jié)合力,NS3_373和NS5b_382則與HLA-A*2402具有高結(jié)合力。
研究表明:與HLA分子結(jié)合力高的候選表位肽不一定是真正的T細胞表位[28]。一種肽到底是不是真正的T細胞表位,不但取決于肽與HLA分子的結(jié)合力,還取決于肽是否為蛋白在人細胞內(nèi)被蛋白酶體裂解產(chǎn)物。而通過采用敏感的免疫學(xué)實驗檢測HCV感染者PBMCs中是否存在某種肽特異性分泌IFN-γ的細胞則可證實該肽是否為人蛋白酶體裂解結(jié)果。ELISPOT和ICS實驗是公認的用于研究細胞免疫的最佳方法,原因是這兩種實驗具有極高的敏感性和可重復(fù)性,前者可以反映單細胞免疫水平,后者則可以確定細胞免疫的表型。因此,這兩種方法可以結(jié)合起來從單細胞水平研究細胞免疫反應(yīng)。在本研究中,我們同時使用ELISPOT和ICS兩種方法檢測肽刺激HCV感染者PBMCs后分泌IFN-γ的細胞的頻率以及表型。結(jié)果顯示,HLA-A*1101陽性HCV感染者PBMCs中存在NS3_609和NS2_165特異性能分泌IFN-γ的CD8+T細胞,HLA-A*2402陽性HCV感染者PBMCs中存在NS3_373和NS5b_382特異性能分泌IFN-γ的CD8+T細胞。證實NS3_609、NS2_165、NS3_373和NS5b_382均是HCV蛋白經(jīng)人細胞內(nèi)蛋白酶體裂解后產(chǎn)生的,從而表明這4條肽為HCV特異性CD8+T細胞表位。
總之,本研究成功鑒定出了2條HCV特異性HLAA*1101限制性CD8+T細胞表位和2條HLA-A*2402限制性CD8+T細胞表位。本研究證實,將永生化的B細胞株和競爭性肽結(jié)合實驗結(jié)合起來是一種極好的檢測候選表位肽與HLA分子結(jié)合力的研究策略。在將來的研究中,我們將探討這些表位誘導(dǎo)的CD8+T細胞能否殺傷HCV感染的靶細胞,對機體有無免疫保護效應(yīng)。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
