電針聯合貞芪扶正顆粒對肝纖維化大鼠自由基反應、NF-κB活化、TGF-β1和CTGF mRNA表達的影響
2014-09-12 08:45:04張超賢郭李柯郭曉鳳
中國老年學雜志 2014年9期
張超賢 郭李柯 郭曉鳳
(新鄉醫學院第一附屬醫院消化內科,河南 衛輝 453100)
在肝臟損傷時,肝星狀細胞(HSCs)受到各種致病因子的刺激,從靜息狀態化轉為具有增生性、纖維原性和可收縮性的肌成纖維細胞, 后者產生大量細胞外基質(ECM)在肝臟沉積, 在肝纖維化發生、發展中起關鍵作用〔1〕。研究表明,自由基反應 、轉化生長因子-β1(TGF-β1)和結締組織生長因子〔2,3〕在促使HSCs增殖與活化,誘導肝纖維化過程中發揮重要作用,核因子(NF)-κB是一種核轉錄因子,能夠被自由基反應產物、內毒素等因素激活,調節包括TGF-β1、結締組織生長因子(CTGF)等多種基因的表達,間接參與肝纖維化的形成〔4〕。電針足三里等穴位及貞芪扶正顆粒具有益氣扶正功能,從中醫理論上推測可能應具有有良好抗肝纖維化作用。本實驗通過電針聯合貞芪扶正顆粒治療二甲基亞硝胺誘導的肝纖維化大鼠。
1 材料和方法
1.1材料 雄性SD大鼠, 3 月齡300 只,體重180~220 g(新鄉醫學院實驗動物中心提供),DMN(日本化成公司), 將DMN 335 μl加0.15 mol/L 的氯化鈉溶液中稀釋至66.96 ml,制備腹腔注射液備用; 貞芪扶正顆粒(15 g/袋,山西德元堂藥業有限公司生產,批準文號:豫藥制字Z14020883);凝膠成像系統掃描儀(UVP公司,美國),Trizol試劑(美國Invitrogen公司),TGF-β1、CTGF及β-actin引物(上海博亞生物技術有限公司),RT-PCR(一步法)試劑盒(大連寶生物工程有限公司)。ROS 、SOD、GSH-Px、CAT 、T-AOC、 LPO檢測試劑盒(南京建成生物工程公司),NF-κB檢測試劑盒(美國RD公司)。
1.2實驗動物分組與模型制作方法 SD大鼠飼養籠飼養,自由飲水,標準飼料喂養。適應環境1 w后,隨機分為五組:正常對照組、模型組、電針組、貞芪扶正顆粒組(蒸餾水稀釋為0.15 g/ml)、針藥聯合組。每組60只大鼠。除正常對照組外,其余各組均給予上述DMN稀釋液隔日1次按2 ml/kg腹腔注射,共4 w。電針組取雙側足三里、關元、內關、三陰交、肝俞穴,采用華佗牌30號寸毫針,針尖圓滑,針刺時大鼠固定為仰臥位,對上述穴位作常規消毒,術者右手持針,針體與皮膚呈90°刺入皮下,進針深度為0.5 cm,連接G6805電針治療儀,將電針儀調整如下:調整輸出調節旋鈕置于1檔(即輸出電壓為1 V保持不變)頻率20 Hz,每次留針30 min,2次/d ,共4 w。貞芪扶正顆粒組予貞芪扶正顆粒稀釋液2 ml/(100 g-1·d-1)分2次灌胃,針藥聯合組則予上述電針和貞芪扶正顆粒治療。正常組和模型組予等量蒸餾水灌胃。在造模過程中模型組死亡2只,貞芪扶正顆粒組死亡1只。實驗4 w后,乙醚吸入麻醉下麻醉下,處死動物并剖腹,取出肝臟并稱重;在肝右葉位置切取小塊組織,置入10%中性甲醛緩沖液固定。其余肝組織分裝于離心管中低溫保存。
1.3肝組織病理 取經10%中性甲醛緩沖液固定的大鼠肝組織,石蠟包埋,切片為5 μl厚度,多級酒精脫水,二甲苯透明,作網狀纖維染色、Masson染色。肝臟纖維化程度分期標準參照文獻〔2,3〕:0級:無纖維化;Ⅰ級:匯管區纖維化擴大,局限竇周及小葉內纖維化; Ⅱ級:匯管區周圍纖維化,纖維間隔形成,小葉結構保留; Ⅲ級:纖維間隔伴小葉結構紊亂,無肝硬化; Ⅳ級:早期肝硬化。利用病理圖像分析儀,對Masson染色膠原定量分析,首先設定窗口面積:每組至少選8個視野,測定窗口范圍內的膠原面積。
1.4肝組織指標的測定 2%的肝組織勻漿10 000 r/min離心,1 min后取上清,4℃保存,分別按試劑盒說明書進行。活性氧族ROS含量測定采用Fenton反應顯色法,抗氧化能力(T-AOC)比色法,超氧化物歧化酶(SOD)的測定采用黃嘌呤氧化法 , 谷胱甘肽過氧化物酶(GSH-Px)活力測定用DTNB直接法,過氧化氫酶(CAT)紫外吸收法, 脂質過氧化物(LPO)含量的測定采用TBA比色法, ELISA法測定NF-κB。
1.5TGF-β1 mRNA的檢測 在4 w末同組織學檢查法取大鼠肝組織,剪成1 cm×1 cm×1 cm大小,儲存于-80 ℃冰箱中,Trizol試劑盒(美國Invitrogen公司)提取大鼠肝組織總RNA, 采用RT-PCR法檢測TGF-β1 mRNA,TGF-β1 mRNA引物序列:上游5'-GGACTACTACGCCAAAGAAG-3',下游5'- TCAAAAGACAGCCACTCAGG -3',擴增產物長度294 bp,內參β-actin序列:上游5'- GTGCCCATCTACGAGGGTTA -3',下游5'-TCTCAGCTGTGGTGGTGAAG -3',擴增產物長度730 bp。PCR條件:94 ℃預變性5 min, 94 ℃變性30 s,退火TGF-β1為54 ℃,β-actin為55 ℃,72 ℃延伸60 s, 30個循環,最后 72 ℃延伸10 min,取PCR反應液8 μl進行瓊脂糖凝膠電泳,分析PCR產物,測定擴增帶(目的條帶和β-actin)的吸收度值,計算TGF-β1 mRNA與β-actin的吸光度比值。
1.6肝組織CTGF mRNA檢測 Trizol試劑盒提取大鼠肝組織總RNA, CTGF上游引物: 5'-CGGGAAATGCTGTGAGGAGT -3',下游引物5'-CAGGCTTGGCAATTTTAGGC -3',擴增產物長度326 bp;β-actin引物序列為上游引物: 5'-GAG CGT GGC TAC AGC TTC AC-3',下游引物5'-GGATGC CAC AGG ATT CCA TA-3',擴增長度120 bp。PCR反應體系為20 μl,PCR循環參數為:94℃預變性3 min;94 ℃變性30 s,54 ℃退火30 s,72 ℃復性30 s,共35個循環,最后72 ℃延伸7 min。每次PCR反應均以無菌雙蒸水代替cDNA模板作為陰性對照。取7 μl PCR產物在15 g/L瓊脂糖凝膠中進行電泳,紫外線下觀察結果,并在凝膠電泳分析系統下拍照。求出每條帶的密度值,以β-actin PCR產物作為內參照。求出每條目的條帶與β-actin條帶的比值,進行統計學處理。

2 結 果
2.1病理學表現 正常組肝細胞排列整齊,胞質均勻,肝小葉及小葉中央靜脈完整,輪廓清楚,肝細胞索網狀纖維結構完整。模型組肝臟廣泛肝細胞脂肪變性、壞死,多數匯管區有淋巴細胞、單核細胞至匯管區向鄰近肝小葉內浸潤,同時小葉內大范圍壞死塌陷;各治療組可見肝組織內炎癥細胞浸潤、肝細胞壞死和纖維組織增生較模型組減輕,纖維間隔較薄(圖1,圖2)。各組肝組織纖維化分期、Masson染色膠原纖維面積比較(表1)。
2.2電針和貞芪扶正顆粒對大鼠肝組織ROS和LPO含量、T-AOC、SOD、GSH-Px、CAT及NF-κB活性、TGF-β1 mRNA及CTGF mRNA表達的影響 模型組肝組織ROS、LPO含量、NF-κB活性、TGF-β1 mRNA及CTGF mRNA較正常對照組顯著增高,T-AOC、SOD、GSH-Px、CAT活性顯著降低(P<0.01),治療組ROS、LPO含量、NF-κB活性、TGF-β1 mRNA及CTGF mRNA明顯低于模型組,T-AOC、SOD、GSH-Px、CAT活性顯著升高(P<0.01),聯合治療組治療效果優于單治療組(P<0.05),NF-κB活性與TGF-β1 mRNA及CTGF mRNA呈正相關關系(r=0.627,P< 0.05;r=0.581,P< 0.05)(表2、圖3)。

正常對照組
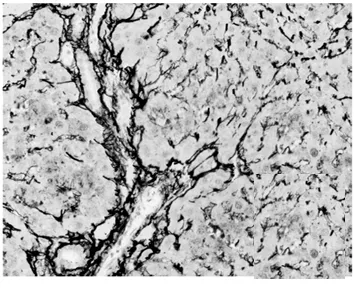
模型組
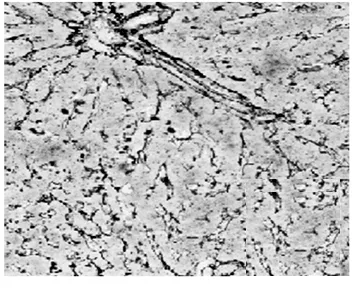
電針組
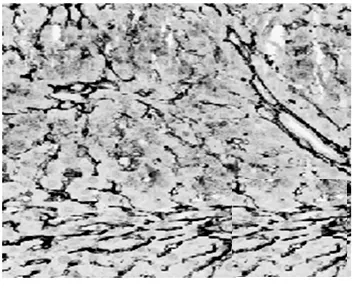
貞芪扶正顆粒組

針藥聯合組
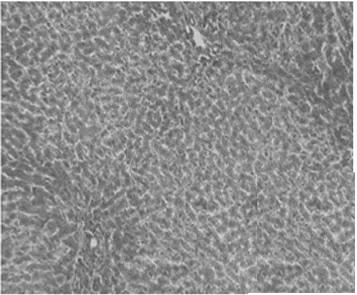
正常對照組
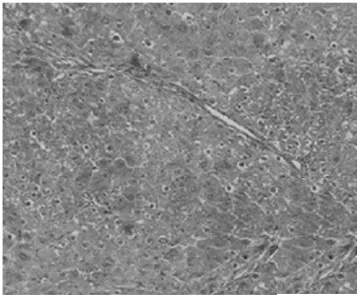
模型組

電針組
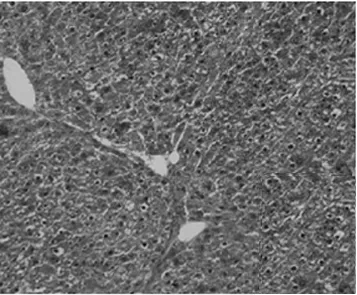
貞芪扶正顆粒組
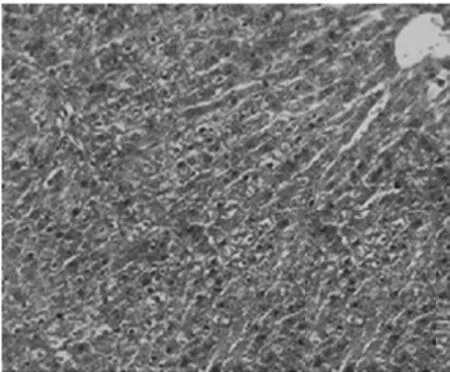
針藥聯合組

表1 各組大鼠肝組織纖維化分期、染色膠原纖維面積比較
表2 各組大鼠肝組織ROS和LPO含量、T-AOC、SOD、GSH-Px、CAT及NF-κB活性、TGF-β1 mRNA及CTGF mRNA表達的比較
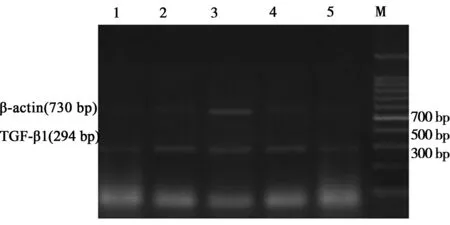
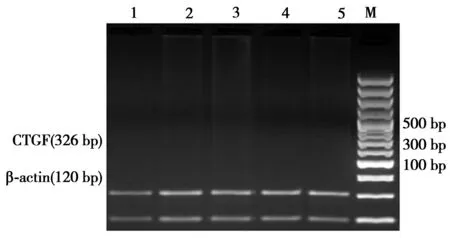
M:Marker;1:正常對照組; 2:模型組;3:電針組; 4: 貞芪扶正顆粒組;5:針藥聯合組
圖3各組肝組織TGF-β1、CTGFmRNA的表達
3 討 論
大量研究發現肝細胞受損時ROS含量明顯增高,ROS能攻擊生物膜磷脂中的多聚不飽和脂肪酸發脂質過氧化反應,形成 LPO,所以LPO在體內的含量反映了脂質過氧化的程度和速率, LPO可通過誘導炎性細胞浸潤, 激活Kuppffers細胞和HSCs, 引發肝纖維化。SOD 是體內存在的一種抗氧化酶 , 能將氧自由基歧化為 H2O2, H2O2進一步在 CAT 、GST-Px 的催化下被清除 ,從而保護細胞免受損傷〔5〕。T-AOC的作用主要是維持內環境活性氧的動態平衡,使機體處于氧化還原相對穩定的狀態,其大小可代表和反映機體抗氧化酶系統和非酶促系統對外來刺激的代償能力以及機體自由基代謝的狀態〔6〕。包括酶促與非酶促兩個體系:酶促體系有SOD、GSH-PX、CAT、谷胱甘肽S-轉移酶(GST)等。聯合測定ROS、T-AOC、SOD、GST-Px 、CAT、 LPO能了解自由基反應與脂質過氧化與疾病的關系,可作為疾病的診斷、預后和抗氧化治療的一個有用指標。
TGF-β1 是目前已知致纖維化最強的因子 ,可以促使活化肝星狀細胞(HSC)和纖維母細胞生長 ,同時 HSC能 自分泌TGF-β1,是肝纖維化后期TGFβ1的主要來源,導致細胞外基質循環不斷產生,從而導致肝纖維化的發生〔7〕。CTGF是一種與肝纖維化密切相關的促纖維化細胞因子,在慢性肝病患者及實驗性動物肝組織中,CTGF表達水平增高,并與肝纖維化記分呈正相關。CTGF可介導TGFβ1、血管內皮生長因子(VEGF)、脂質過氧化物等多種信號刺激的促纖維化作用,可促進包括HSCs在內的多種纖維活性細胞合成并分泌ECM〔8〕。 NF-κB是一種具有多向轉錄調節作用的蛋白, 在靜止期細胞的胞質中,NF-κB處于P50、P65與抑制蛋白IκB結合的多聚體非活化狀態。當細胞受到細菌、病毒、活性氧、過氧化脂質、化療藥物等外源性刺激時,多聚體上的IκB磷酸化降解,NF-κB活化,進入細胞核內與特定基因啟動因子區域位點結合,啟動免疫相關受體、細胞因子、炎癥因子、黏附分子等靶基因的轉錄,產生大量的包括TGFβ1、VECG、CTGF、TNF-α等細胞因子〔9,10〕。
綜上,氧自由基和脂質過氧化物可能是NF-κB重要刺激物,可活化NF-κB,促進CTGF等相應基因的轉錄,LPO不僅可活化Kupffer細胞,分泌TGF-β1等細胞毒性因子,繼而激活HSCs,還能直接激活HSCs,而活化的HSCs是CTGF主要來源,TGF-β1也可促進CTGF的轉錄,CTGF反過來又可介導TGFβ1、脂質過氧化物等多種信號刺激的促纖維化作用,可促進包括HSCs在內的多種纖維活性細胞合成并分泌ECM ,所以在肝纖維化發生、發展過程中,自由基反應、NF-κB、 TGF-β1和CTGF是相互協同、相互促進的,形成一種正反饋網絡, 因此若能抑制脂質過氧化、抑制NF-κB活化,下調TGF-β1及CTGF表達,則有希望減少HSCs活化、增殖和膠原的沉積達到發揮抗纖維化的作用。
肝纖維化屬于祖國醫學“脅痛”,“癥積”,“瘀血”等范疇。其病因為:情志郁結,氣失條達;飲食不節,嗜酒過度,蟲積水毒所傷及病后繼發。以上病因致使肝脾內傷,肝失疏泄,脾失健運,肝郁氣滯,而使血行不暢,久則血瘀絡阻。故正虛血瘀是肝纖維化的基本病機,貞芪扶正顆粒是采用黃芪、女貞子等中藥為主要原料,提取有效成分,研制而成,具有扶正固本、益氣補虛、活血化瘀的功效。從中醫理論上貞芪扶正顆粒應具有良好的抗肝纖維化作用。現代藥理研究已證明:黃芪甲苷可阻止TNF-α、細菌脂多糖 誘導NF-κB的核移位與靶基因的κB位點的結合,抑制 TGF-β1、CTGF基因的轉錄〔11〕;黃芪總黃酮和黃芪總皂苷清除自由基的主要機制是結構中的酚羥基類基團在與氧自由基作用時,通過提供氫原子,發生氧化還原反應而發揮作用〔12〕。另褪黑素是強有力的內源性抗氧化劑,其抗氧化作用包括直接清除自由基,間接刺激抗氧化酶活性及基因表達,刺激谷胱甘肽合成、減少線粒體電子傳遞鏈電子滲漏、和其他抗氧化劑有協同效應,而黃芪總皂苷則可促進機體褪黑素分泌或加強其與受體結合而增強機體的抗氧化能力〔13,14〕;黃芪有效成分黃芪多糖、硒、木糖-葡萄糖-環黃芪醇可啟動DNA修補酶、刺激抗體產生、清除有害自由基 ,啟動使谷胱甘肽過氧化物酶、阻止黃嘌呤及黃嘌呤酶的氧化損傷發揮抗氧化作用〔15〕。
針刺穴位可激發經絡本身協調陰陽的功能,泄其有余,補其不足,陰陽平復,達到調整虛實防治疾病的作用。現代研究表明,神經內分泌系統與免疫系統之間有密切的雙向調節聯系。免疫細胞上有多種神經內分泌激素受體,多種激素會影響免疫功能,免疫細胞也可合成神經遞質和激素影響內分泌系統,神經-內分泌-免疫系統紊亂參與肝癌發病。針刺可影響神經內分泌激素和細胞因子的生成和表達,雙向調節神經-內分泌-免疫網絡,使異常機能向有利于機體的方向轉化。有研究表明針灸對提高免疫系統機能的調整功能是扶正的物質基礎,又是除邪的物質條件,表明針灸"補虛瀉實、扶正祛邪"的理論是科學的,其調整功能是可靠的。現代針灸機制研究表明針刺足三里、關元、內關、三陰交等穴可明顯提高大鼠SOD、GSH-Px、CAT活性,清除自由基,抑制脂質過氧化物的生成〔16~19〕。
本研究進一步印證了自由基反應、脂質過氧化、NF-κB異常活化TGF-β1及CTGF高表達在肝纖維化中的作用;研究說明電針和貞芪扶正顆粒均能提高機體抗氧化能力,降低自由基產和脂質過氧化物的生成,抑制 NF-κB的活化,下調TGF-β1 mRNA及CTGFF mRNA表達,打破了自由基反應-NF-κB的活化-TGF-β1 mRNA促纖維化的正反饋機制,從而達到抗纖維化目的,這與現代醫學所揭示的針刺和貞芪扶正顆粒的作用機制相符;另本研究發現,電針和貞芪扶正顆粒聯合應用時效果更佳,提示兩者在治療肝纖維化時可起協同作用,為臨床針藥聯合治療肝纖維化提供了理論和實驗依據。
4 參考文獻
1Lu P,Liu H,Yin H,etal.Expression of angiotensinogen during hepatic fibrogenesis and its effect on hepatic satellate cells〔J〕.Med Sci Monit,2011;17(9):BR248-56.
2吳文娟,楊妙芳,朱人敏.肝纖維化發生過程中的信號傳導通路〔J〕.肝臟,2008;13(2):166-8.
3Gressner OA,Lahme B,Siluschek M,etal.Connective tissue growth factor is a Smad regulated amplifier of transforming growth factor beta actions in hepatocytes--but without modulating bone morphogenetic protein 7 signaling〔J〕.Hepatology,2009;49(6):2021-30.
4Inoue J,Gohda J,Akiyama T,etal.NF-kappaB activation in development and progression of cancer〔J〕.Cancer Sci,2007;98(3):268-74.
5江海艷,王春妍,楊世忠.化瘀通絡解毒法對肝纖維化大鼠血清超氧化物歧化酶、丙二醛和轉化生長因子β1 的影響〔J〕.中國實驗方劑學雜志,2007;13(6):59-60.
6杜 鵬,周麗梅,楊昌林.暈船易感大鼠腦干總抗氧化能力、超氧化物歧化酶及活性氧的活性測定〔J〕.海軍醫學雜志,2009;30(4):293-6.
7Cho IJ,Kim YW,Han CY,etal.E-cadherin antagonizes transforming growth factor β1 gene induction in hepatic stellate cells by inhibiting RhoA-dependent Smad3 phosphorylation〔J〕.Hepatology,2010;52(6):2053-64.
8Liu Y,Meyer C,Müller A,etal.IL-13 induces connective tissue growth factor in rat hepatic stellate cells via TGF-β-independent Smad signaling〔J〕.J Immunol,2011;187(5):2814-23.
9Inoue J,Gohda J,Akiyama T,etal.NF-kappaB activation in development and progression of cancer〔J〕.Cancer Sci,2007;98(3):268-74.
10Acharyya S,Villalta SA,Bakkar N,etal. Interplay of IKK/NF-kappaB signaling in macrophages and myofibers promotes muscle degeneration in Duchenne muscular dystrophy〔J〕.J Clin Invest,2007;117(4):889-901.
11Li HL,Chen HL,Li H,etal.Regulatory effects of emodin on NF-kappaB activation and inflammatory cytokine expression in RAW 264.7 macrophages〔J〕.Int J Mol Med,2005;16(1):41-7.
12段琦梅,梁宗鎖,聶小妮,等.黃芪和黨參提取物的抗氧化活性研究〔J〕.西北植物學報,2010;30(10):2123-7.
13王懷清,劉志民.褪黑素及其代謝物抗氧化作用研究進展〔J〕.國際內分泌代謝雜志,2009;29(2):129-31.
14李燕舞,宋 寧,王汝俊.黃芪總苷對應激大鼠胃黏膜氧自由基及褪黑素受體的影響〔J〕.世界華人消化雜志,2008;16(29) :3321-3.
15陳國輝,黃文鳳.黃芪化學成份、藥理活性與臨床應用概述〔J〕.海峽藥學, 2008;20(3):13-6.
16梁 宜,方劍喬,汪存信.經皮穴位電刺激對力竭運動大鼠血漿超氧化物歧化酶和二醛的影響〔J〕.針刺研究,2008;33(2):120-3.
17程井軍,彭 銳,吳其愷.電針對非酒精性脂肪性肝炎大鼠肝組織氧化及抗氧化狀態的影響〔J〕.中國康復,2008;23(3):158-60.
18李 野,洪英杰,朱慧明.電針對亞急性衰老大鼠肝與血清SOD MDA T-AOC的影響〔J〕.遼寧中醫藥大學學報,2008;10(7):132-4.
19樸慧花,于 濤,劉新橋.針刺對心肺復蘇后兔血漿SOD、MDA的影響〔J〕.江蘇中醫藥,2010;42(3):76-7.
