小干擾RNA抑制MDM2對HepG2細胞的影響及機制
2014-09-12 03:22:08趙燕穎李亞剛趙春燕楊澤成張舵舵孫遠杰
中國老年學雜志 2014年1期
關鍵詞:肝癌
趙燕穎 李亞剛 趙春燕 楊澤成 張舵舵 孫遠杰
(吉林大學第四臨床醫院內科,吉林 長春 130011)
肝癌存在多種原癌基因的激活和抑癌基因的失活、突變,最終導致其編碼蛋白質的改變,從而引起細胞惡變〔1〕。MDM2的基因產物可以和野生型p53(wtp53)結合,可以起到一種對p53的負調節作用,因為MDM2基因的過度表達,可以減少p53基因的抑癌作用,即使是低水平MDM2的表達,也將會引起細胞成瘤性的增強〔2〕。目前已有大量研究表明在消化系統腫瘤如胃癌、肝癌、胰腺癌中MDM2都是過渡表達的〔3〕。siRNA技術已經成為研究基因功能的一種公認的有效工具,并且可以用于篩選藥物靶點,為治療腫瘤、病毒感染帶來突破。目前尚未見用siRNA技術干擾MDM2表達對人肝癌細胞的報道。本文作者設計siRNA MDM2并轉染人肝癌細胞,觀察其對細胞增殖和凋亡的影響和對MDM2及其上下游基因蛋白的表達影響,探討其作用機制。
1 材料與方法
1.1材料 胎牛血清、IMDM培養基 (Gibco);Trizol Reagent,逆轉錄和PCR試劑盒(Invitrogen)、SilencerTMsiRNA construction kit(Ambion 公司);LipofectamineTM2000 轉染試劑盒(Invitrogen公司);引物由上海生工生物技術公司合成;細胞株(中國科學院上海細胞所)。兔抗人MDM2、wtp53、p21、Bcl-2和β-actin抗體及羊抗兔二抗購于UPSTATE,Hybond 硝酸纖維素膜 (NC) 購自Amersham。
1.2siRNA 的設計和合成 根據已知Gen Bank MDM2 mRNA序列和siRNA的設計原則〔4〕,利用Ambion 公司的在線設計來尋找靶序列,并做同源序列搜索,排除那些和其他編碼序列或表達序列標簽(EST)同源的序列,用SilencerTMsiRNA construction kit體外合成siMDM2-1,siMDM2-2,陰性對照(control siRNA)購于Ambion,與任何編碼序列無同源性。
1.3細胞培養及轉染 人肝癌細胞HepG2細胞株由中科院上海細胞所提供。在含10%小牛血清的IMDM培養液中培養,于5%CO2和37℃條件下培養。HepG2細胞采用脂質體LipofectamineTM2000進行轉染。取對數生長期的HepG2細胞用0.25 %胰酶消化后,制成單細胞懸液,用細胞記數板進行細胞記數。取96孔板和10 ml培養瓶。96孔板每個培養皿接種細胞數為1×104,培養瓶接種1×106,37℃,5%CO2條件下培養24 h后進行轉染。做好分組標志(Ⅰ組:PGCsilencerTM-MDM2 siRNA-1;Ⅱ組:PGCsilencerTM-MDM2 siRNA-2;Ⅲ組:PGCsilencerTM-control siRNA;Ⅳ組:只加轉染試劑lipofectin)。轉染步驟依據LipofectamineTM2000轉染說明書。
1.4RT-PCR檢測siMDM2對細胞基因的影響 轉染后72 h用Trizol 提取各組細胞中的總RNA。總RNA中逆轉錄出cDNA,再通過PCR法擴增,操作均嚴格按試劑盒說明書進行。以β-actin作為內參,MDM2擴增產物為300 bp,上游5′-AACCACCTCACAGATTCCAG-3′,下游5′-TCAAGGTGACACCTGTTCTC-3′;p53擴增引物為490 bp,上游5′-GTGGTGGTGCCCTATGAG -3′;下游5′-GGGAGGTAGACTGACCCTTT-3′;P21擴增產物為486 bp,上游5′-GCTGAGCCGCGACTGTGAT-3′下游5′-TGAGACTAAGGCAGAAGATGTA-3′;Bcl2擴增產物為382 bp,上游5′-GACCGAAGTCCGCAGAAC-3′,下游5′-CCAACGTGCCATGTGCTA-3′;β-actin 擴增產物為 520 bp,上游:5′-GGGACCTGACAGACTACCT-3′,下游:5′-CGTACTCCTGCTTGCTGA-3′。PCR 產物經1 %瓊脂糖凝膠系統電泳(含溴乙啶),拍照鑒定。用Pharmacia ImageMaster 凝膠成像儀觀察,并用 ImageMaster 軟件分析條帶峰面積,轉染各組信號值相對于對照組的百分率進行統計作圖。
1.5MTT法檢測siMDM2對細胞增殖的影響 轉染siMDM-1、2和control siRNA,每組4復孔,培養24,48,72 h 后每孔加入5 g/ L MTT 溶液15 μl,37℃孵箱中繼續孵育4 h后吸棄孔內培養上清液,每孔加150 μl二甲基亞砜(DMSO),振蕩10 min,用酶聯免疫檢測儀570 nm測定各孔吸光度,記錄結果。以時間、吸光度繪制細胞增殖曲線,細胞增殖抑制率(%)=(1-處理組吸光度/未處理組吸光度)×100%,計算出抑制率。
1.6流式細胞儀檢測siMDM2對細胞凋亡的影響 轉染后繼續培養72 h后收集各組的HepG2細胞磷酸鹽緩沖液(PBS)洗滌,70%冰乙醇固定,碘化丙啶染色,流式細胞儀檢測。通過軟件Multicycle進行分析,以DNA含量為橫坐標,受檢細胞為縱坐標作圖,用累積曲線分割法計算各期細胞所占比例。G1峰前面出現的亞2倍體峰即為凋亡峰,計算各組凋亡率。
1.7劃痕實驗檢測siMDM2對細胞遷移能力的影響 將HepG2細胞種于24孔培養板中,用含10%胎牛血清(FBS)的IMDM培養基常規培養。次日待細胞融合度達到80%~90%時,用中號槍頭進行劃痕,PBS洗3次以去除劃下的細胞,加入新鮮的培養液,轉染,培養48 h。熒光顯微鏡拍照紀錄。比較轉染前后劃痕寬度的變化。
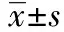
2 結 果
2.1重組質粒酶切和測序鑒定 PGCsilencerTM-MDM2 siRNA經Hind Ⅲ和BamH Ⅰ雙酶切后呈2條帶,分成大的載體片段和小的目的片段。將重組質粒PGCsilencerTM-MDM2-siRNA送上海生工測序(測序號D02.CS060503093_0253.SD13D229-1.F;D04.CS060503093_0255.SD13D230-1.F),引物序列根據載體序列,采用Primer-Premier5.0設計為:5′-AAGAAGAGAGTGTGGAATCTA-3′。結果表明siRNA表達模板成功構建于PGCsilencer載體上,序列完全正確,與目的序列相同。見圖1。

1:Marker;2:siMDM2-1;3:siMDM2-1/HindⅢ+BamHⅠ;4:siMDM2-2;5:siMDM2-2/HindⅢ+BamHⅠ;6:control siRNA;7:control siRNA / Hind Ⅲ+ BamH Ⅰ;8:Marker
2.2siMDM2轉染后細胞基因表達的變化 以β-actin作為內參照,對靶基因mRNA進行RT-PCR結果定性定量分析,各組引物的擴增基因片段經電泳和HE染色后,結果顯示擴增片段大小與所設計的大小完全一致。siMDM2-1,2對基因表達影響趨勢一致,轉染2組siMDM2后MDM2和Bcl2 mRNA表達水平有顯著的下調,p53和p21 mRNA表達水平有顯著上調,而轉染陰性質粒組各基因表達水平無明顯改變。半定量掃描分析,和空白組比較轉染2組si-MDM2的HepG2中MDM2 mRNA的表達量平均下調61.7%;p53 mRNA表達量平均上調46.9%;p21 mRNA表達量平均上調41.8%;Bcl2 mRNA平均下調39.5%,和空白組比均具有顯著性差異(P<0.05,n=4); control siRNA組各基因表達量與空白對照組無明顯差異(P>0.05,n=4)。見圖2。
2.3siMDM2 轉染后細胞增殖的變化 MTT法檢測顯示轉染siMDM2后HepG2細胞增殖受到不同程度的抑制,轉染2組si MDM2后24、48、72 h的平均抑制率為(30.9±2.99)%、(45.6±3.15)%和(52.1±3.29)%。抑制率隨時間的增加而增高,各轉染組抑制率明顯高于control siRNA組,control siRNA組細胞增殖未見明顯改變。而且siMDM2-1組抑制率稍高于siMDM2-2組,但差異無統計學意義(P>0.05)。
2.4siMDM2轉染后細胞凋亡的變化 轉染siMDM2可以誘導HepG2細胞凋亡,轉染2組siMDM2的細胞凋亡率分別為(41.9±2.59)%和(34.9±1.98)%,明顯高于control siRNA組(2.6±0.39)%(P<0.05)。同時,和空白組比siMDM2組細胞處于S期的細胞比例明顯減少;而處于G0/G1期的細胞比例明顯增加。
2.5siMDM2轉染后細胞遷移能力的變化 control組與control siRNA組遷移較快,而siMDM2組遷移速率變慢,control組與control siRNA組之間差異無顯著性(P>0.05),各組siMDM2之間差異無顯著性(P>0.05)。siMDM2組遷移距離與空白組和control siRNA組相比,差異有顯著性意義(P<0.05)。見圖3。

圖2 轉染后各組MDM2、P53、P21和Bcl2基因表達的變化

圖3 轉染后HepG2細胞遷移能力的變化
3 討 論
目前原癌基因、抑癌基因、凋亡基因等癌癥相關基因的過度表達被認為是肝癌發生的主要原因。一些研究表明,可以用RNAi技術抑制原癌基因,突變的抑癌基因,細胞周期相關基因,抗凋亡相關基因等的表達來抑制肝癌的發生和發展〔1〕。肝癌的發生是多基因參與的結果,隨著肝癌分子機制研究的不斷深入,可成為RNAi技術靶位點的基因也越來越多,因此,如能利用RNAi技術抑制癌基因的表達,從而影響上下游基因表達變化,就可能抑制肝癌的發生和發展。
MDM2作為一種癌基因,其生物學作用是增強細胞的增殖活力,使細胞生存期延長,促進細胞增生及腫瘤的生長〔5〕。MDM2能與p53結合,抑制p53的抑癌功能。p53和Bcl-2同屬于細胞凋亡的重要調控因子,在調節細胞凋亡的過程中有相互聯系、共同作用的關系〔6〕。Farah等〔7〕認為p53基因誘導細胞凋亡的機制可能為:降低細胞內源性Bcl-2蛋白表達和抑制其功能,p53基因與Bcl-2蛋白表達呈明顯負相關。另外,MDM2蛋白還可以與p21結合,終止p21被p53活化的抑制細胞增生的作用〔8,9〕。P21是ras基因家族中一種共同表達產物,是細胞周期負性調控基因,可被野生型p53蛋白誘導表達而使細胞周期停滯于G1期。MDM2過表達可以下調p53和p21表達,上調Bcl-2的表達從而刺激細胞無限制性生長,這可能在某些腫瘤的發生及發展中起著關健性作用。
本文結果顯示siMDM2可抑制MDM2基因的表達從而促進p53表達,最終導致p21表達增加和Bcl-2表達降低,這可能是siMDM2抑制腫瘤細胞增殖和誘導其凋亡的主要機制之一。
本課題組在siRNA MDM2對前列腺癌增殖和凋亡的影響研究基礎上〔10〕,對干擾MDM2在肝癌治療中作用的進一步研究。研究結果基本一致,進一步證明了本研究組設計合成的siRNA MDM2質粒可以抑制腫瘤細胞增殖和誘導凋亡,同時可以抑制腫瘤細胞的遷移能力。siRNA技術為腫瘤基因治療提供了一個新的可供選擇的策略和技術平臺,本研究亦表明MDM2可能是腫瘤治療的一個新的靶點。
4 參考文獻
1Zeng XX,Zheng RL,Zhou T,etal.Novl thienopyridine derivatives as specific anti- hepatocellular carcinoma (HCC) agents:synthesis,preliminary structure-activity relationships,and in vitro biological evaluation〔J〕.Bioorg Med Chem Lett,2010;20(21):6281-5.
2Akkiz H,Sümbül AT,Bayram S,etal.MDM2 promoter polymorphism is associated with increased susceptibility to hepatocellular carcinoma in Turkish population〔J〕.Cancer Epidemiol,2010;34(4):448-52.
3Kang KF,Zhang X,Chen XW,etal.Expressions of MDM2 and MMP-7 in normal liver,cirrhosis,hepatocellular carcinoma tissues〔J〕.Zhonghua Gan Zang Bing Za Zhi,2010;18(1):67-8.
4Elbashir SM,Harborth J,Weber K,etal.Analysis of gene function in somatic mammalian cells using small interfering RNAs〔J〕.Methods,2002;26(2):199-213.
5劉冬滿,何世東,張志勇.子宮內膜癌中p15及MDM2蛋白表達〔J〕.中國老年學雜志,2010;30(4):1936-7.
6Esposito F,Tornincasa M,Chieffi P,etal.High-mobility group A1 proteins regulate p53- mediated transcription of Bcl-2 gene〔J〕.Cancer Res,2010;70(13):5379-88.
7Farah IO,Begum RA,Ishaque AB,etal.Differential protection and transactivation of P53,P21,Bcl2,PCNA,cyclin G,and MDM2 genes in rat liver and the HepG2 cell line upon exposure to pifithrin〔J〕.Biomed Sci Instrum,2007;43:116-21.
8Rong JJ,Hu R,Song XM,etal.Gambogic acid triggers DNA damage signaling that induces p53/p21(Waf1/CIP1) activation through the ATR-Chk1 pathway〔J〕.Cancer Lett,2010;296(1):55-64.
9Xu H,Zhang Z,Li M,etal.MDM2 promotes proteasomal degradation of p21Waf1 via a conformation change〔J〕.J Biol Chem,2010;285(24):18407-14.
10李然偉,趙燕穎,楊澤城,等.siRNA抑制人前列腺癌細胞PC3的MDM2表達及細胞增殖〔J〕.腫瘤防治研究,2009;36(1):24-7.
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
