鎘脅迫對(duì)不同鎘富集能力海水養(yǎng)殖貝類抗氧化能力的影響
——以扇貝和菲律賓蛤仔為例
2014-09-21 11:59:48趙艷芳吳繼法翟毓秀寧勁松盛曉風(fēng)丁海燕尚德榮
生態(tài)毒理學(xué)報(bào) 2014年2期
關(guān)鍵詞:差異
趙艷芳,吳繼法,翟毓秀,寧勁松,盛曉風(fēng),丁海燕,尚德榮,*
1. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所,青島266071 2. 青島市黃島區(qū)海洋與漁業(yè)局,青島 266400
鎘脅迫對(duì)不同鎘富集能力海水養(yǎng)殖貝類抗氧化能力的影響
——以扇貝和菲律賓蛤仔為例
趙艷芳1,吳繼法2,翟毓秀1,寧勁松1,盛曉風(fēng)1,丁海燕1,尚德榮1,*
1. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所,青島266071 2. 青島市黃島區(qū)海洋與漁業(yè)局,青島 266400
為研究具有不同鎘(Cd)富集能力的扇貝和菲律賓蛤仔在相同鎘脅迫環(huán)境下的抗氧化能力反應(yīng)的差異,將2種貝類于0.05 mg·L-1Cd環(huán)境中暴露10 d,分別于第0天、2天、4天、6天、8天、10天分別取樣檢測(cè)內(nèi)臟團(tuán)中SOD、CAT、GPx、GST酶活性和GSH含量,同時(shí)分析內(nèi)臟團(tuán)中Cd的含量。結(jié)果表明,對(duì)于Cd脅迫組,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中SOD和CAT酶活反應(yīng)相似,均呈先被誘導(dǎo)后被抑制的規(guī)律,且2種貝類的SOD和CAT活性差異較小。2種貝類內(nèi)臟團(tuán)中GSH含量、GST和GPx活性變化差異較大:扇貝內(nèi)臟團(tuán)中GSH含量顯著降低(p<0.05),GST和GPx活性均在第2天和第4天時(shí)處于顯著誘導(dǎo)狀態(tài)(p<0.05),從第6天時(shí)處于抑制狀態(tài),而菲律賓蛤仔內(nèi)臟團(tuán)中GSH含量、GST和GPx活性在Cd暴露期間無顯著變化,且2種貝類間比較,扇貝內(nèi)臟團(tuán)中GSH含量和GST活性在整個(gè)實(shí)驗(yàn)期間始終顯著高于菲律賓蛤仔(p<0.05),GPx活性從第4天開始高于菲律賓蛤仔;對(duì)于無鎘污染的對(duì)照組,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中SOD、CAT和GPx活性無顯著差異,但扇貝內(nèi)臟團(tuán)中GSH含量和GST活性顯著高于菲律賓蛤仔,其中前者GSH含量約為后者的22倍。研究同時(shí)表明在相同鎘環(huán)境下扇貝內(nèi)臟團(tuán)對(duì)Cd的富集濃度和富集速率均遠(yuǎn)高于菲律賓蛤仔。本研究明確了2種貝類在相同鎘環(huán)境下的抗氧化反應(yīng)的差異,推斷出扇貝內(nèi)臟團(tuán)中高含量的GSH以及較高的GST和GPx活性可能在扇貝高富集、高耐受Cd方面起重要作用。
鎘;扇貝;菲律賓蛤仔;抗氧化能力;鎘富集能力
鎘(cadmium, Cd)是一種易在生物體內(nèi)蓄積且難代謝的有害元素,被美國毒物管理委員會(huì)(ATSDR)列為第6位危及人類健康的有毒物質(zhì)。鎘廣泛存在于自然環(huán)境中,也可通過食物鏈傳遞,對(duì)消費(fèi)者的身體健康造成威脅。大洋中鎘的濃度很低(0.01~0.25 μg·L-1),近岸海域鎘的濃度可達(dá)5.38 μg·L-1[1]。雙殼貝類能夠從環(huán)境中富集高濃度的鎘。但是不同貝類具有不同的富集能力,如扇貝對(duì)鎘的富集能力約是蛤仔的10倍甚至更高[2],而且扇貝在正常環(huán)境中也能積累高濃度的鎘[3-5]。相同環(huán)境中雙殼貝類對(duì)重金屬的富集能力不同可能是由于其對(duì)鎘具有不同的富集能力和解毒策略。目前已有研究指出,扇貝能夠富集高濃度的鎘,可能是由于扇貝對(duì)鎘具有較高的消化吸收率、同化利用率和釋放速率[6-8],以及具有有效的解毒策略包括溶酶體的區(qū)隔化作用、與磷酸鹽顆粒的共沉淀及與金屬硫蛋白(metallothioneins, MT)結(jié)合等[3]。
重金屬鎘對(duì)水生生物的毒性作用機(jī)制之一是造成生物體內(nèi)的過氧化脅迫,在長(zhǎng)期進(jìn)化中,需氧生物發(fā)展了防御過氧化損害的酶系統(tǒng),在生理狀態(tài)下,由代謝產(chǎn)生的活性氧可為抗氧化防御系統(tǒng)所控制,機(jī)體抗氧化防御系統(tǒng)中的一些成分會(huì)改變,從而使機(jī)體免受氧化傷害[9]。由自由基介導(dǎo)的生物體內(nèi)的氧化損傷是鎘離子毒性效應(yīng)的主要體現(xiàn)。其過氧化脅迫主要有2種方式:直接誘導(dǎo)生物體內(nèi)產(chǎn)生活性氧(ROS)、自由基代謝物;間接降低生物體清除氧自由基的能力。超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GPx)、谷胱甘肽S-轉(zhuǎn)移酶(GST)等抗氧化物酶能夠清除ROS和多種自由基,在生物體解毒機(jī)制中發(fā)揮著至關(guān)重要的作用[10-11]。因此,筆者推測(cè)2種具有不同鎘富集能力的貝類的抗氧化反應(yīng)肯定存在重要差異。本實(shí)驗(yàn)試圖通過比較研究扇貝和菲律賓蛤仔在相同鎘脅迫環(huán)境下的抗氧化反應(yīng)差異,從抗氧化能力角度解釋2種貝類對(duì)鎘的蓄積和耐受能力差異。本研究選擇的抗氧化反應(yīng)指標(biāo)包括SOD、CAT、GPx、GST活性以及谷胱甘肽(GSH)含量。
1 材料與方法(Materials and methods)
1.1 實(shí)驗(yàn)材料
扇貝(Chlamys farreri)和菲律賓蛤仔(Ruditapes philippinarum)于2012年10月取自青島一貝類養(yǎng)殖廠。扇貝平均殼高(5.8±0.34) cm,殼長(zhǎng)(5.5±0.26) cm,菲律賓蛤仔平均殼高(2.1±0.26) cm,殼長(zhǎng)(3.0±0.28) cm,在60 L玻璃缸中用自然海水暫養(yǎng)1周,保持水體連續(xù)充氣,每日換水1次,水溫(20.0±1.0) ℃,pH=8.0,自然光照。
1.2 實(shí)驗(yàn)設(shè)計(jì)
鎘脅迫實(shí)驗(yàn)養(yǎng)殖容器為無毒白色塑料周轉(zhuǎn)箱565 mm×420 mm×300 mm,將每個(gè)實(shí)驗(yàn)容器中加入30 L自然海水,根據(jù)實(shí)驗(yàn)生物個(gè)體大小差異,分別隨機(jī)放入扇貝和菲律賓蛤仔各30和150只。
CdCl2(CdCl2·2.5H2O)為分析純,購自國藥集團(tuán)化學(xué)試劑有限公司。首先將其配制成1 g·L-1母液,再稀釋至實(shí)驗(yàn)所需濃度。鎘的實(shí)驗(yàn)濃度為0.05 mg·L-1。取樣時(shí)間分別為0、2、4、6、8和10 d。各處理組每個(gè)實(shí)驗(yàn)容器分別取櫛孔扇貝6只個(gè)體,菲律賓蛤仔30只個(gè)體。空白對(duì)照組為在不添加鎘離子的正常自然海水,實(shí)驗(yàn)期間不投餌,每天定時(shí)換水,實(shí)驗(yàn)養(yǎng)殖條件同暫養(yǎng)條件,并每天檢查貝的存活情況。
1.3 樣品制備
隨機(jī)從Cd脅迫處理組和空白對(duì)照組中取出活貝進(jìn)行解剖。迅速取出內(nèi)臟團(tuán),用濾紙吸凈表面水分,稱取5.000 g組織即刻放入-80 ℃冰箱冷凍保存。
1.4 指標(biāo)測(cè)定方法
SOD、CAT、GPX、GST活性和GSH含量測(cè)定采用南京建成生物工程研究所試劑盒方法測(cè)定。蛋白質(zhì)含量測(cè)定采用南京建成生物工程研究所試劑盒測(cè)定方法-考馬斯亮藍(lán)法(Bradford法)測(cè)定。
1.5 鎘含量分析
稱取2.000 g內(nèi)臟團(tuán)試樣于100 mL玻璃消化管中,加入混合酸(V(硝酸)∶V(高氯酸)= 4∶1)浸泡過夜后,用控溫消化爐消化,同時(shí)做試劑空白對(duì)照。試樣中Cd含量采用石墨爐原子吸收(Perkin Elmer, AA600)測(cè)定。
1.6 數(shù)據(jù)處理
試驗(yàn)結(jié)果用平均值±標(biāo)準(zhǔn)差(Mean±SD)表示,實(shí)驗(yàn)數(shù)據(jù)用Duncan' s多重比較法將各處理組進(jìn)行比較,并利用SPSS 16.0統(tǒng)計(jì)分析軟件,采用單因素方差分析(One-Way ANOVA)進(jìn)行統(tǒng)計(jì),當(dāng)p<0.05時(shí),認(rèn)為差異顯著。
2 結(jié)果(Results)
2.1 扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中Cd含量
在0.05 mg·L-1Cd脅迫下,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中Cd含量變化趨勢(shì)見表1。從表中可見,2種貝類內(nèi)臟團(tuán)均對(duì)Cd有顯著富集作用。對(duì)于空白對(duì)照組,扇貝內(nèi)臟團(tuán)中Cd含量為6.89 μg·g-1(濕重),約為菲律賓蛤仔內(nèi)臟團(tuán)中含量的43倍。在Cd暴露實(shí)驗(yàn)期間內(nèi),2種貝類內(nèi)臟團(tuán)中Cd的含量一直呈上升趨勢(shì),并且鎘含量相對(duì)于暴露時(shí)間呈強(qiáng)正相關(guān)關(guān)系(扇貝組:R2=0.9868,p<0.01;菲律賓蛤仔組:R2=0.9663,p<0.01)。暴露2、4、6、8和10 d后,扇貝內(nèi)臟團(tuán)對(duì)Cd的富集速率分別為1.46、1.74、1.52、1.42、1.49 μg·(g·d)-1,菲律賓蛤仔內(nèi)臟團(tuán)對(duì)Cd的富集速率分別為0.037、0.059、0.12、0.17、0.19 μg·(g·d)-1。因此,在相同Cd濃度環(huán)境下,扇貝內(nèi)臟團(tuán)對(duì)Cd的富集速率遠(yuǎn)大于菲律賓蛤仔。
2.2 Cd對(duì)扇貝和菲律賓蛤仔SOD活性的影響
2種貝類內(nèi)臟團(tuán)SOD的活性變化趨勢(shì)相似,均呈先誘導(dǎo)后抑制的規(guī)律,并且在脅迫4 d后達(dá)到最高。另外,空白對(duì)照組中,2種貝類內(nèi)臟團(tuán)SOD的活性相似(p>0.05)。第4天和第8天,扇貝中SOD活性顯著高于菲律賓蛤仔(p<0.05),其余時(shí)間點(diǎn)2種貝類內(nèi)臟團(tuán)的SOD活性均相似(p>0.05)。
2.3 Cd對(duì)扇貝和菲律賓蛤仔CAT活性的影響
Cd脅迫下扇貝和菲律賓蛤仔CAT活性的變化見圖2,可知2種貝類的CAT活性變化趨勢(shì)相同,且與SOD活性變化規(guī)律相似(見圖1),均呈先誘導(dǎo)后抑制的趨勢(shì),脅迫4 d后CAT活性達(dá)到最高。空白對(duì)照組中,2種貝類內(nèi)臟團(tuán)CAT的活性無顯著差異(P>0.05),且均是在第4天和第8天,扇貝中CAT活性顯著高于菲律賓蛤仔(p<0.05),其余取樣時(shí)間點(diǎn)2種貝類的CAT活性相似(p>0.05)。

表1 扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中Cd含量Table 1 Cd contents in visceral mass of Chlamys farreri and Ruditapes philippinarum
注:Cd含量單位為μg·g-1(濕重)。
Note: the unit of Cd content is μg·g-1, wet weight.
2.4 Cd對(duì)扇貝和菲律賓蛤仔GST活性的影響
Cd脅迫對(duì)2種貝類內(nèi)臟團(tuán)中GST活性的影響見圖3,從圖中可見,扇貝內(nèi)臟團(tuán)中GST活性在暴露4 d內(nèi)處于顯著誘導(dǎo)狀態(tài),第4天時(shí)達(dá)到最高為629.5 U·mg-1prot,約是空白對(duì)照組的2倍,隨后逐漸被抑制,10 d后,降至空白對(duì)照組水平(p>0.05)。而對(duì)于菲律賓蛤仔,在整個(gè)實(shí)驗(yàn)期間,GST活性變化較小。

圖1 Cd脅迫對(duì)扇貝和菲律賓蛤仔內(nèi)臟團(tuán)SOD活性的影響注:圖中不同小寫字母表示p<0.05差異顯著。Fig. 1 SOD activity in the visceral mass of C. farreri and R. philippinarum under 0.05 mg·L-1 cadmium exposureNote: the different small letters indicate a significant difference at p<0.05.

圖2 Cd脅迫對(duì)扇貝和菲律賓蛤仔內(nèi) 臟團(tuán)CAT活性的影響注:圖中不同小寫字母表示p<0.05差異顯著。Fig. 2 CAT activity in visceral mass of C. farreri and R. philippinarum under 0.05 mg·L-1 cadmium exposureNote: the different small letters indicate a significant difference at p<0.05.
另外在整個(gè)暴露實(shí)驗(yàn)期間,扇貝內(nèi)臟團(tuán)中GST的活性均顯著性高于菲律賓蛤仔(P<0.05)。扇貝空白對(duì)照組的GST活性(303 U·mg-1prot)約為菲律賓蛤仔對(duì)照組(157 U·mg-1prot)的2倍。
2.5 Cd對(duì)扇貝和菲律賓蛤仔GPx活性的影響 0.05 mg·L-1Cd脅迫下,扇貝內(nèi)臟團(tuán)GPx活性的活性變化規(guī)律(見圖4)與GST的活性變化規(guī)律相似,在第2天和第4天時(shí)處于顯著誘導(dǎo)狀態(tài)(p<0.05),第4天時(shí)活性達(dá)到最高值為100 U·mg-1prot,約為空白對(duì)照組(30.2 U·mg-1prot)的3倍多。對(duì)于菲律賓蛤仔,整個(gè)暴露實(shí)驗(yàn)期間,GPx活性變化較小,未達(dá)到差異顯著水平(p>0.05)。另外,不同于GST活性,空白對(duì)照組2種貝類內(nèi)臟團(tuán)GPx活性無顯著差異(p>0.05)。

圖3 Cd脅迫對(duì)扇貝和菲律賓蛤仔內(nèi)臟團(tuán)GST活性的影響注:圖中不同小寫字母表示p<0.05差異顯著。Fig. 3 GST activity in visceral mass of C. farreri and R. philippinarum under 0.05 mg·L-1 cadmium exposureNote: the different small letters indicate a significant difference at p<0.05.

圖4 Cd脅迫對(duì)扇貝和菲律賓蛤仔 內(nèi)臟團(tuán)GPx活性的影響注:圖中不同小寫字母表示p<0.05差異顯著。Fig. 4 GPx activity in visceral mass of C. farreri and R. philippinarum under 0.05 mg·L-1 cadmium exposureNote: the different small letters indicate a significant difference at p<0.05.
2.6 Cd對(duì)扇貝和菲律賓蛤仔GSH含量的影響
圖5顯示了Cd脅迫下,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)GSH含量的變化。從圖中可見,2種貝類GSH含量變化存在很大差異。首先,對(duì)于空白對(duì)照組,扇貝內(nèi)臟團(tuán)中GSH含量為78 nmol·mg-1prot,而菲律賓蛤仔內(nèi)臟團(tuán)中GSH含量為3.5 nmol·mg-1prot,前者約為后者的22倍。且在暴露實(shí)驗(yàn)期間,扇貝內(nèi)臟團(tuán)中的GSH含量始終顯著高于菲律賓蛤仔(p<0.05)。0.05 mg·L-1Cd脅迫下,扇貝中GSH含量急劇下降,2 d后降至24.5 nmol·mg-1prot,且6 d后降至最低點(diǎn)。而對(duì)于菲律賓蛤仔,空白對(duì)照組和Cd暴露組在實(shí)驗(yàn)期間內(nèi)臟團(tuán)中GSH含量無顯著變化(p>0.05)。2.7 扇貝和菲律賓蛤仔抗氧化能力比較
為了更清楚地表明扇貝和菲律賓蛤仔2種貝類,在相同濃度Cd脅迫下的抗氧化能力反應(yīng)差異,將扇貝空白組的4種抗氧化酶(SOD、CAT、GST、GPx)酶活性和GSH含量定義為100%,比較扇貝Cd處理組不同時(shí)間點(diǎn)、菲律賓蛤仔空白和Cd處理組不同時(shí)間點(diǎn)的抗氧化酶活性和GSH含量的相對(duì)變化。結(jié)果見表2,從表中可見,對(duì)于SOD和CAT,扇貝和菲律賓蛤仔的酶活性高低和變化規(guī)律均相似;對(duì)于GST,相對(duì)于扇貝空白組,菲律賓蛤仔空白組僅占52%,并且在Cd脅迫下,菲律賓蛤仔的GST活性
最高升至73%,至暴露結(jié)束后,降至空白組水平(53%),而扇貝在Cd脅迫下,GST活性于暴露4 d時(shí)升高了2倍(207.6%),約為菲律賓蛤仔(73.3%)的3倍;對(duì)于GPx,在Cd脅迫4 d時(shí),扇貝處理組最高為菲律賓蛤仔的2倍;對(duì)于GSH含量,相對(duì)于扇貝空白組,菲律賓蛤仔空白組的GSH含量?jī)H占4.5%,在Cd脅迫下,扇貝內(nèi)臟團(tuán)中GSH含量急劇下降,脅迫2 d時(shí)即下降至空白組31.3%,脅迫6 d時(shí),含量約占空白組的13.8%,而菲律賓蛤仔于Cd脅迫10 d后,GSH含量最高,占扇貝空白對(duì)照組的5.0%。

圖5 Cd脅迫對(duì)扇貝和菲律賓蛤仔內(nèi)臟團(tuán)GSH含量的影響注:圖中不同小寫字母表示P<0.05差異顯著。Fig. 5 GSH content in visceral mass of C. farreri and R. philippinarum under 0.05 mg·L-1 cadmium exposureNote: the different small letters indicate a significant difference at p<0.05.
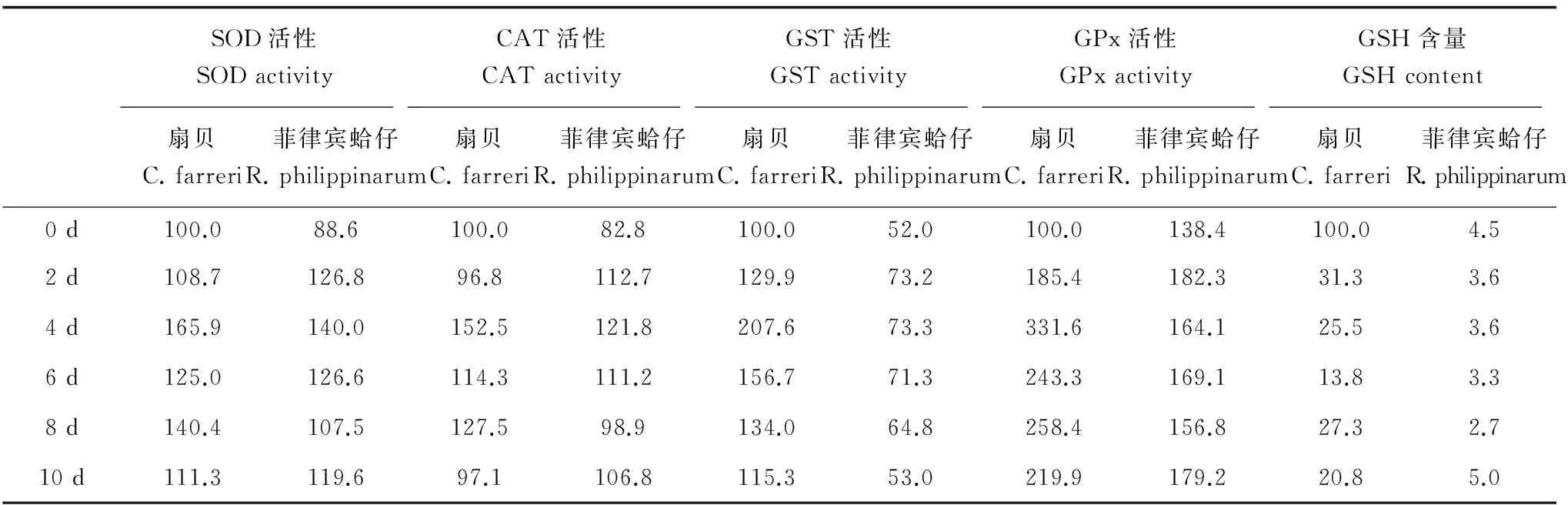
表2 扇貝和菲律賓蛤仔4種抗氧化酶活性和GSH含量隨暴露時(shí)間的相對(duì)變化(%)Table 2 Comparative changes in activities of four antioxidant enzymes and GSH content for both C. farreri and R. philippinarum with the extension of exposure time (%)
3 討論(Discussion)
海水水質(zhì)標(biāo)準(zhǔn)GB 3097—1997的漁業(yè)標(biāo)準(zhǔn)中,Cd≤0.005 mg·L-1,已有研究表明0.005 mg·L-1Cd脅迫下,貝類組織中抗氧化酶在實(shí)驗(yàn)期間與對(duì)照組相比無顯著變化,當(dāng)鎘濃度為標(biāo)準(zhǔn)濃度的10倍時(shí),貝類產(chǎn)生抗氧化反應(yīng)[12]。本研究目的是試圖通過比較2種貝類對(duì)鎘的抗氧化能力差異,來解釋2種貝類對(duì)鎘蓄積和耐受能力的差異,因此選擇0.05 mg·L-1Cd為脅迫實(shí)驗(yàn)濃度。
實(shí)驗(yàn)分析了空白對(duì)照組2種貝類中Cd的蓄積量,結(jié)果顯示扇貝整貝中Cd含量為9.7 mg·kg-1,約為菲律賓蛤仔整貝中Cd含量(0.27 mg·kg-1)的35倍(數(shù)據(jù)未展示),再次證明了扇貝在相同無鎘污染的正常環(huán)境中能夠在體內(nèi)積累高濃度的鎘,含量約為蛤仔的幾十倍[2-5]。扇貝內(nèi)臟團(tuán)中Cd含量為6.89 mg·kg-1,菲律賓蛤仔內(nèi)臟團(tuán)中Cd含量為0.16 mg·kg-1,因此,內(nèi)臟團(tuán)是扇貝和菲律賓蛤仔富集鎘的主要組織。0.05 mg·L-1Cd脅迫10 d,2種貝類內(nèi)臟團(tuán)中的鎘均沒有達(dá)到飽和,內(nèi)臟團(tuán)中鎘含量與暴露時(shí)間呈顯著正相關(guān)關(guān)系。這可能是由于在慢性鎘暴露初期,鎘進(jìn)入貝類體內(nèi)后首先進(jìn)入內(nèi)臟團(tuán),與MT、GSH、Cys等巰基化合物結(jié)合[13],使其毒性降低,并蓄積在內(nèi)臟團(tuán)內(nèi),同時(shí)導(dǎo)致內(nèi)臟團(tuán)內(nèi)SOD、CAT、GST、GPX等抗氧化系統(tǒng)指標(biāo)發(fā)生變化。
3.1 扇貝和菲律賓蛤仔在相同Cd環(huán)境下抗氧化反應(yīng)相同之處
在0.05 mg·L-1Cd脅迫下,2種貝類內(nèi)臟團(tuán)中SOD和CAT活性變化趨勢(shì)相似,均呈先誘導(dǎo)后抑制的變化規(guī)律。已有研究表明,Cd2+能夠誘導(dǎo)機(jī)體產(chǎn)生大量的自由基和活性氧(ROS),引起脂質(zhì)、蛋白質(zhì)和DNA的氧化變性,誘發(fā)組織損傷[14]。SOD是生物體內(nèi)唯一一種以自由基為底物的抗氧化酶,它可將超氧陰離子自由基歧化為H2O2和O2,以達(dá)到清除體內(nèi)活性氧自由基的作用,避免細(xì)胞受到氧自由基的損害[15-16]。CAT是機(jī)體抗氧化體系中又一關(guān)鍵非特異抗氧化酶,能夠清除SOD催化活性氧自由基所產(chǎn)生的H2O2,使其轉(zhuǎn)化為對(duì)機(jī)體無毒的H2O和O2,它們的作用都是使機(jī)體免受氧自由基的攻擊[17]。本實(shí)驗(yàn)中,0.05 mg·L-1Cd脅迫下,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中SOD活性均先升高后降低,在暴露4 d后,SOD活性達(dá)到最高值,顯示其被顯著誘導(dǎo),之后呈下降的趨勢(shì),但是相對(duì)于空白組仍然被顯著誘導(dǎo)。隨著SOD活性增加,CAT活性也有增加的趨勢(shì),并在暴露4 d后達(dá)到最大值,之后逐漸降低。這說明,扇貝與菲律賓蛤仔在Cd的脅迫下,機(jī)體會(huì)產(chǎn)生適應(yīng)性誘導(dǎo)效應(yīng),當(dāng)較少的鎘離子進(jìn)入機(jī)體并誘導(dǎo)其產(chǎn)生活性氧自由基時(shí),SOD的活性升高以清除氧自由基,從而促使機(jī)體內(nèi)H2O2含量升高。H2O2的升高誘導(dǎo)了機(jī)體內(nèi)CAT的基因表達(dá)及活力,提高了H2O2的分解速率。但是如果暴露時(shí)間增加,扇貝和菲律賓蛤仔體內(nèi)鎘離子含量升高,大量的不能被及時(shí)清除的氧自由基被誘發(fā)產(chǎn)生,SOD、CAT的合成途徑被阻礙或者其亞單位間的裝配發(fā)生了改變,從而導(dǎo)致其活力開始下降,這種低濃度時(shí)的“毒物興奮效應(yīng)”與在真鯛、長(zhǎng)江華溪蟹、褶紋冠蚌、尖紫蛤、泥蚶等生物中觀察到的結(jié)果類似[12,18-21]。另外,在無鎘污染的正常環(huán)境中,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中2種酶的活性無顯著差異,本研究結(jié)果可以推斷出SOD、CAT酶可能不是影響2種貝類具有不同積累和耐受鎘能力差異的主要因素。
3.2 扇貝和菲律賓蛤仔在相同Cd環(huán)境下抗氧化反應(yīng)不同之處
在0.05 mg·L-1Cd環(huán)境下,扇貝和菲律賓蛤仔內(nèi)臟團(tuán)中的GSH含量、GST和GPx活性變化存在一定差異。在整個(gè)實(shí)驗(yàn)期間,扇貝內(nèi)臟團(tuán)中GSH含量和GST酶活均顯著高于菲律賓蛤仔(p<0.05)。而且在無鎘污染的正常環(huán)境下,扇貝內(nèi)臟團(tuán)中的GSH含量和GST酶活性顯著高于菲律賓蛤仔(p<0.05)。其中差異最大的是GSH含量,前者約為后者的22倍。GSH是細(xì)胞內(nèi)含巰基最豐富的物質(zhì),參與物質(zhì)代謝和轉(zhuǎn)運(yùn)過程,對(duì)各種外源(如重金屬)和內(nèi)源性物質(zhì)(如活性氧)對(duì)細(xì)胞的損害都有保護(hù)作用[22]。GSH可在GST的催化下與細(xì)胞內(nèi)的有害物質(zhì)結(jié)合,生成無毒或毒性小的GSH硫結(jié)合物,具有消除體內(nèi)脂質(zhì)過氧化物和解毒的雙重作用。有研究表明生物體由于污染的暴露產(chǎn)生適應(yīng)性誘導(dǎo)反應(yīng)而使GSH含量升高,也可以由于污染的毒性作用產(chǎn)生中毒反應(yīng)而使GSH含量降低[23]。如比薩茶蝸牛(Theba pisana)受到以Cu2+為主要成分的農(nóng)藥刺激后,消化腺中的GSH含量出現(xiàn)了明顯下降,并且刺激的Cu2+濃度越高,GSH含量下降越明顯[24]。在鎘脅迫下,扇貝內(nèi)臟團(tuán)中GSH含量先下降后上升,暴露開始GSH含量下降說明大量的GSH與Cd結(jié)合,以降低Cd對(duì)生物體的毒性,隨著暴露時(shí)間的延長(zhǎng),GSH含量逐漸升高,說明扇貝產(chǎn)生可適應(yīng)性誘導(dǎo)效應(yīng)。而GST酶活性則呈現(xiàn)與GSH相反的變化趨勢(shì),這是由于GSH是GST的自然底物,Cd脅迫初期,大量GST酶被激活,催化GSH與Cd2+及過氧化物結(jié)合,使生物體內(nèi)還原性的GSH消耗增加。而對(duì)于菲律賓蛤仔,Cd暴露組的蛤仔內(nèi)臟團(tuán)中GSH含量、GST酶活性無論是與空白對(duì)照組比較,還是不同暴露時(shí)間點(diǎn)之間比較,變化均較小,且均顯著低于扇貝。
GPx也是生物體普遍存在的一種抗氧化物酶。在GST的催化下,GPx作為親電子集團(tuán)的結(jié)合靶位點(diǎn),可以降低內(nèi)、外源毒物對(duì)機(jī)體產(chǎn)生的毒性作用,也可以分解過氧化氫和過氧化物,清除細(xì)胞內(nèi)脂質(zhì)和有機(jī)過氧化反應(yīng)的產(chǎn)物,阻斷脂質(zhì)過氧化連鎖反應(yīng),達(dá)到保護(hù)細(xì)胞膜機(jī)構(gòu)和功能完整的作用,與CAT酶在細(xì)胞內(nèi)具有相似的作用[25]。本實(shí)驗(yàn)也證明了扇貝中GPx活性變化趨勢(shì)與CAT變化趨勢(shì)相似,均于4 d后活性達(dá)到最高,約為空白對(duì)照組的3倍,并且顯著高于菲律賓蛤仔。Cd2+還可以與GPx分子中的Se形成Cd-Se復(fù)合物,破壞GPx活性中心Se代半胱氨酸的機(jī)構(gòu),一方面降低了鎘離子對(duì)機(jī)體的毒性影響,達(dá)到解除重金屬鎘毒性的目的;另一方面使GPx分子活性中心的Se代半胱氨酸結(jié)構(gòu)被破壞而使GPx活性降低[26]。因此,隨著暴露鎘時(shí)間的增加,扇貝中GPx活性逐漸降低。但是菲律賓蛤仔內(nèi)臟團(tuán)中GPx活性在Cd暴露期間變化不顯著,并且空白對(duì)照組中2種貝類GPx活性無顯著差異。
生物體對(duì)逆境反應(yīng)存在的差異與抗氧化系統(tǒng)的整體活力相關(guān)。重金屬污染地區(qū)的貝類、魚、沙蠶等的抗氧化能力遠(yuǎn)遠(yuǎn)強(qiáng)于未污染地區(qū)的個(gè)體[27-29],重金屬污染地區(qū)的植物的抗氧化反應(yīng)強(qiáng)于非污染地區(qū)的植物[30]。如砷/鎘超富集植物比非砷/鎘超富集植物具有更強(qiáng)的抗氧化能力,且較強(qiáng)的抗氧化能力在超富集植物解毒、蓄積重金屬的機(jī)制中起關(guān)鍵作用[31-33]。本研究結(jié)果明確顯示,在相同Cd暴露環(huán)境下,扇貝的抗氧化能力比菲律賓蛤仔強(qiáng),主要表現(xiàn)在扇貝內(nèi)臟團(tuán)具有較高的GSH含量、GST活性和GPx活性,且扇貝對(duì)鎘的富集量和富集速率遠(yuǎn)高于菲律賓蛤仔。因此可以推斷扇貝中較高的GSH含量、GST和GPx活性可能使扇貝具有較高的Cd解毒能力,也可能是其具有高富集Cd能力的一個(gè)主要原因。
[1] Ferreira A C, Costa A C S, Korn M G A. Preliminary evaluation of the cadmium concentration in seawater of the Salvador city, Brazil [J]. Microchemical Journal, 2004, 78(1): 77-83
[2] Pigeot J, Miramand P, Guyot T, et al. Cadmium pathways in an exploited intertidal ecosystem with chronic cadmium inputs (Marennes-Oleron, Atlantic coast, France) [J]. Marine Ecology Progress Series, 2006, 307: 101-114
[3] Bustamante P, Miramand P. Subcellular and body distributions of 17 trace elements in the variegated scallop Chlamys varia from the French coast of the Bay of Biscay [J]. Science of the Total Environment, 2005, 337(1-3): 59-73
[4] Bustamante P, Miramand P. Interspecific and geographical variations of trace element concentrations in Pectinidae from European waters [J]. Chemosphere, 2004, 57(10): 1355-1362
[5] Pan K, Wang W X. The subcellular fate of cadmium and zinc in the scallop Chlamys nobilis during waterborne and dietary metal exposure [J]. Aquatic Toxicology, 2008, 90(4): 253-260
[6] Metian M, Warnau M, Oberh?nsli F, et al. Interspecific comparison of Cd bioaccumulation in European Pectinidae (Chlamys varia and Pecten maximus) [J]. Journal of Experimental Marine and Ecology, 2007, 353(1): 58-67
[7] Wang W X, Rainbow P S. Comparative approaches to understand metal bioaccumulation in aquatic animals [J]. Comparative Biochemistry and Physiology - Part C, 2008, 148(4): 315-323
[8] Ng T Y T, Wang W X. Detoxification and effects of Ag, Cd and Zn pre-exposure on metal uptake kinetics in the clam Ruditapes philippinarum [J]. Marine Ecology Progress Series, 2004, 268: 161-172
[9] 劉慧, 王曉蓉, 張景飛, 等. 銅及其EDTA配合物對(duì)彭澤鯽魚肝臟抗氧化系統(tǒng)的影響[J]. 環(huán)境化學(xué), 2004, 23(3): 263-267
Liu H, Wang X R, Zhang J F, et al. Effects of copper and complex (Cu-EDTA) on the antioxidant system of Carassius auratos liver [J]. Environmental Chemistry, 2004, 23(3): 263-267 (in Chinese)
[10] Zikic R V, Stajn A S, Saicic Z S, et al. The activities of superoxide dismutase, catalase and ascorbic acid content in the liver of goldfish (Carassius auratus gibelio Bloch) exposed to cadmium [J]. Physiological Research, 1996, 45(6): 479-481
[11] Almeidaa J A, DinizbY S, Marquesa S F G, et al. The use of the oxidative stress responses as biomarkers in Nile tilapia (Oreochromis niloticus) exposed to in vivo cadmium contamination [J]. Environment International, 2002, 27(8): 673-679
[12] 鄧思平, 趙云濤, 朱春華, 等. 鎘對(duì)尖紫蛤抗氧化酶活性及脂質(zhì)過氧化的影響[J]. 水生生物學(xué)報(bào), 2012, 36(4): 689-695
Deng S P, Zhao Y T, Zhu C H, et al. Effect of cadmium on the antioxidant enzyme activity and lipid perosidation in Sanguinolaria acuta [J]. Acta Hydrobiologica Sinica, 2012, 36(4): 689-695 (in Chinese)
[13] 趙艷芳, 尚德榮, 寧勁松, 等. 體積排阻高效液相色譜-電感耦合等離子體質(zhì)譜法測(cè)定海產(chǎn)貝類中鎘的形態(tài)[J]. 分析化學(xué), 2012, 40(5): 681-686
Zhao Y F, Shang D R, Ning J C, et al. Speciation analysis of cadmium in marine bivalves by size exclusion chromatography-high performance liquid chromatography-inductively coupled plasma mass spectrometry [J]. Chinese Journal of Analytical Chemistry, 2012, 40(5): 681 -686 (in Chinese)
[14] Hegedus A, Erdei S, Horvath G. Comparative studies of H2O2detoxifying enzymes in green and greening barley seedlings under cadmium stress [J]. Plant Science, 2001, 160(6): l085-l093
[15] Pastor A, Medina J, Delramo J. Determination of lead in treated crayfish Procambarus clarkia: Accumulation in different tissues [J]. Bulletin of Environmental Contamination and Toxicology, 1988, 41(3): 412-418
[16] Papadimitriou E, Loumbourdis N S. Exposure of the frog Rana ridibunda to copper impact on two biomarkers, lipid peroxidation, and glutathione [J] . Bulletin of Environmental Contamination and Toxicology, 2002, 69(6): 885-891
[17] Sarkar S, Yadav P, Trivedi R. Cadmium induced lipid peroxidation and the status of the antioxidant system in rat tissues [J]. Journal of Trace Elements in Medicine and Biology, 1995, 9(3): 144-149
[18] Mourente G, Diaz-Salvago E, Bell J G. Increased activities of hepatic antioxidant defense enzymes in juvenile gilthead sea bream (Sparus aurata L.) fed dietary oxidized oil: Attenuation by dietary vitamin E [J]. Aquaculture, 2002, 214 (1-4): 343-361
[19] 李琰, 吳丹, 文春根, 等. 鎘對(duì)褶紋冠蚌抗氧化因子的影響[J]. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào), 2012, 31(11): 2108-2113
Li Y, Wu D, Wen C G, et al. Effects of cadmium on the antioxidant factors of Cristaria plicata [J]. Journal of Agro-Environment Science, 2012, 31(11): 2108-2113 (in Chinese)
[20] 金芬芬, 徐團(tuán), 秦圣娟, 等. 鎘對(duì)長(zhǎng)江華溪蟹肝胰腺線粒體抗氧化酶活力和脂質(zhì)過氧化水平的影響[J]. 水生生物學(xué)報(bào), 2011, 35(6): 1019-1024
Jin F F, Xu T, Qin S J, et al. Effects of cadmium on hepatopancreas mitochondrial antioxidant enzyme activity and lipid peroxidation level in freshwater carb Sinopotamon yangtsekiense [J]. Acta Hydrobilogica Sinica, 2011, 35(6): 1019-1024 (in Chinese)
[21] 孫彬, 葛奇?zhèn)? 陸慧賢, 等. 鎘對(duì)泥蚶抗氧化酶系統(tǒng)的影響[J]. 生態(tài)科學(xué), 2011, 30(4): 383-388
Sun B, Ge Q W, Lu H X, et al. Effects of cadmium exposure on antioxidant enzyme system of Tegillarca granosa [J]. Ecological Science, 2011, 30(4): 383-388 (in Chinese)
[22] 惠天朝, 王家剛, 朱蔭湄. 鎘對(duì)羅非魚肝組織中GSH代謝的影響[J]. 浙江大學(xué)學(xué)報(bào): 農(nóng)業(yè)與生命科學(xué)版, 2001, 27(5): 575-578
Hui T C, Wang J G, Zhu Y M. Effects of Cd on hepatic GSH metabolism in Tilapia nilotica [J]. Journal of Zhejiang University: Agriculture and Life Science, 2001, 27(5): 575-578 (in Chinese)
[23] Stebbing A R. Hormesis-the stimulation of growth by low levels of inhibitors [J]. Science of the Total Environment, 1982, 22(3): 213-234
[24] EI-Gendy K S, Radwan M A, Gad A F. In vivo evaluation of oxidative stress biomarkers in the land snail, Theba pisana exposed to copper based pesticides [J]. Chemosphere, 2009, 77(3): 339-344
[25] 閆博, 王蘭, 李涌泉, 等. 鎘對(duì)長(zhǎng)江華溪蟹肝胰腺抗氧化酶活性的影響[J]. 動(dòng)物學(xué)報(bào), 2007, 53(6): 1121-1128
Yan B, Wang L, Li Y Q, et al. Effects of cadmium on hepatopancreatic antioxidant enzyme activity in a freshwater crab Sinopotamon yangtsekiense [J]. Acta Zoologica Sinica, 2007, 53(6): 1121-1128 (in Chinese)
[26] Iszard M B, Liu J, Klaassen C D. Effect of several metallothionein inducers on oxidative stress defense mechanisms in rats [J]. Toxicology, 1995, 104(1-3): 25-33
[27] Yildirim N C, Benzer F, Danabas D. Evaluation of environmental pollution at Munzur river of Tunceli applying oxidative stress biomarkers in Capoeta trutta (Heckel, 1843) [J]. The Journal of Animal and Plant Sciences, 2011, 21(1): 66-71
[28] Box A A, Sureda F, Galgani A, et al. Assessment of environmental pollution at Balearic Islands applying oxidative stress biomarkers in the mussel Mytilus galloprovincialis [J]. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology, 2007, 146(4): 531-539
[29] Geracitano L A, Bocchetti R, Monserrat J M, et al. Oxidative stress responses in two population of Laeonereis acuta (Polychaeta, Nereididae) after acute and chronic exposure to copper [J]. Marine Environmental Research, 2004, 58(1): 1-17
[30] Kandziora C M, Nadgorska S A, Barczyk G. A comparative study of heavy metal accumulation and antioxidant responses in Vaccinium myrtillus L. leaves in polluted and non-polluted areas [J]. Environmental Science and Pollution Research, 2013, 20(7): 4920-4932
[31] Srivastava M, Ma L Q, Singh N, et al. Antioxidant responses of hyper-accumulator and sensitive fern species to arsenic [J]. Journal of Experimental Botany, 2005, 56(415): 1335-1342
[32] 謝飛, 王宏鑌, 王海娟, 等. 砷脅迫對(duì)不同砷富集能力植物葉片抗氧化酶活性的影響[J]. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào), 2009, 28(7): 1379-1385
Xie F, Wang H B, Wang H J, et al. Effects of arsenic stress on activities of antioxidant enzymes in the fronds of plants with different abilities to accumulate arsenic [J]. Journal of Agro-Environment Science, 2009, 28(7): 1379-1385 (in Chinese)
[33] 張軍, 束文圣. 植物對(duì)重金屬鎘的耐受機(jī)制[J]. 植物生理與分子生物學(xué)學(xué)報(bào), 2006, 32(1): 1-8
Zhang J, Shu W S. Mechanisms of heavy metal cadmium tolerance in plants [J]. Journal of Plant Physiology and Molecular Biology, 2006, 32(1): 1-8 (in Chinese)
◆
EffectsofCadmiumStressontheAntioxidantResponsesinTwoBiovalveswithDifferentAccumulationAbilitiesforCadmium:ACaseStudyontheScallopChlamysfarreriandtheClamRuditapesphilippinarum
Zhao Yanfang1, Wu Jifa2, Zhai Yuxiu1, Ning Jinsong1, Sheng Xiaofeng1, Ding Haiyan1, Shang Derong1,*
1. Yellow Sea Fishery Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China 2. Ocean and Fishery Bureau of Huangdao District of Qingdao City, Qingdao 266400, China
8 March 2013accepted25 June 2013
In order to find out the difference in the antioxidant responses in two bivalves (the scallop Chlamys farreri and the clam Ruditapes philippinarum) which have different abilities for accumulating cadmium (Cd), two bivalves were exposed to 0.05 mg L-1Cd for 2, 4, 6, 8 and 10 days. The activities of SOD, CAT, GPx and GST, the content of GSH and the concentration of cadmium in the visceral mass of two bivalves were analyzed. Results showed that for the treated groups, the activities of SOD and CAT in the two bivalves displayed similar response variability, which were induced firstly and then were inhibited during the period of 10 days. In addition, the activities of SOD and CAT were similar in two bivalves. However, GSH content, GST and GPx activities had great differences in two bivalves. The GSH content in C. farreri significantly decreased (p<0.05), while the GST and GPx activities were significantly induced on the second and fourth day (p<0.05), and were inhibited from the sixth day. In contrast, the GST and GPx activities, and GSH content in R. philippinarum had no significant changes. In addition, GSH content and GST activity in C. farreri were significantly higher than those of R. philippinarum during the 10-day exposure. The GPx activity in C. farreri was also higher (p<0.05) than that of R. philippinarum from the fourth day. For the control groups, the activities of SOD, CAT and GPx had no significant differences between C. farreri and R. philippinarum, while the GSH content and GST activity in C. farreri were much higher than that of R. philippinarum. Specifically, the GSH content in C. farreri was almost 22 times of that of R. philippinarum in the control groups. Moreover, C. farreri had higher Cd accumulation rate and concentration than those of R. philippinarum. It is found that the two bivalves with different abilities for accumulating Cd did show different responses in antioxidant systems under the same Cd-stress conditions. High content of GSH, and higher GST and GPx activities may help to explain the scallop C. farreri have higher ability to defense and detoxify the Cd than the clam R. philippinarum.
cadmium; Chlamys farreri; Ruditapes philippinarum; antioxidant defenses: Cd accumulation abilities
國家自然科學(xué)基金(41206101)
趙艷芳(1979-),女,博士,研究方向?yàn)橛泻χ亟饘偕锘瘜W(xué),E-mail: yanfang_z1979@163.com;
*通訊作者(Corresponding author),E-mail: shangdr@ysfri.ac.cn
10.7524/AJE.1673-5897.20130308004
趙艷芳,吳繼法,翟毓秀,等. 鎘脅迫對(duì)不同鎘富集能力海水養(yǎng)殖貝類(扇貝和菲律賓蛤仔)抗氧化能力的影響[J]. 生態(tài)毒理學(xué)報(bào), 2014, 9(2): 224-232
Zhao Y F, Wu J F, Zhai Y X, et al. Effects of cadmium stress on the antioxidant responses in two biovalves (the scallop Chlamys farreri and the clam Ruditapes philippinarum) with different abilities to accumulate cadmium [J]. Asian Journal of Ecotoxicology, 2014, 9(2): 224-232 (in Chinese)
2013-03-08錄用日期2013-06-25
1673-5897(2014)2-224-09
X171.5
A
尚德榮(1960—),女,副研究員,主要研究方向?yàn)槲廴旧锘瘜W(xué),發(fā)表學(xué)術(shù)論文20余篇。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50

- 生態(tài)毒理學(xué)報(bào)的其它文章
- 鄰苯二甲酸二(2-乙基己)酯(DEHP)和鄰苯二甲酸單乙基己基酯(MEHP)長(zhǎng)期暴露對(duì)海洋青鳉(Oryzias melastigma)內(nèi)分泌干擾效應(yīng)的評(píng)價(jià)
- 微囊藻毒素在太湖白鰱體內(nèi)的累積規(guī)律及其影響因素
- 打印機(jī)使用過程中顆粒物等有害因素釋放及其毒理學(xué)效應(yīng)研究進(jìn)展
- 我國六價(jià)鉻淡水水生生物安全基準(zhǔn)推導(dǎo)研究
- 多氯聯(lián)苯對(duì)我國土壤微生物的生態(tài)毒理效應(yīng)
- 沉積物中六氯苯對(duì)搖蚊幼蟲的慢性毒性效應(yīng)