DENV2感染外周血單個核細胞增加TNF-α基因啟動子區(qū)域CpG位點的甲基化水平*
2014-11-08 02:27:48張英可齊一鳴黃俊琪
中國病理生理雜志 2014年1期
關(guān)鍵詞:區(qū)域
張英可, 張 林, 齊一鳴, 黃俊琪
(中山大學中山醫(yī)學院免疫學研究所,教育部熱帶病防治研究重點實驗室,廣東廣州510080)
登革病毒(dengue virus,DENV)是一種重要的蚊媒病毒,能夠引起登革熱(dengue fever,DF)和重癥登革熱即登革出血熱(dengue hemorrhagic fever,DHF)和登革休克綜合征(dengue shock syndrome,DSS),重癥登革熱死亡率極高。最新文獻報道每年登革病毒感染人數(shù)約有39 000萬人[1]。Jaiyen等[2]指出DENV引起外周血單個核細胞(peripheral blood mononuclear cells,PBMC)凋亡,原因不僅在于病毒本身的毒性作用,也與產(chǎn)生的促炎因子腫瘤壞死因子 α(tumour necrosis factor-alpha,TNF-α)和白細胞介素1β(interleukin-1beta,IL-1β)有關(guān)。本課題組之前對細胞因子的研究發(fā)現(xiàn),2型登革病毒(dengue virus type 2,DENV2)感染 PBMC 0 h、6 h 和 12 h,隨時間增加TNF-α 的產(chǎn)生減少[3]。
DNA甲基化作為表觀遺傳學的重要組成部分之一,能夠通過影響轉(zhuǎn)錄因子的結(jié)合和染色體的結(jié)構(gòu)引起基因表達抑制[4]。在脊椎動物中,DNA甲基化多發(fā)生在含高密度CpG雙核苷酸結(jié)構(gòu)的CpG島區(qū),但是TNF-α啟動子區(qū)不存在典型CpG島結(jié)構(gòu),僅有散在的CpG雙核苷酸[5]。TNF-α的表達調(diào)控受多種因素的影響,DNA甲基化修飾在這個過程中起重要作用。本研究通過在PBMC感染病毒后不同時間檢測TNF-α基因啟動子區(qū)域的甲基化水平,探討甲基化水平對該基因表達調(diào)控的影響。
材料和方法
1 材料
登革病毒2型 New Guinea C株(DENV2,NGC株)來自中山大學中山醫(yī)學院微生物學教研室,由本實驗室進行擴增和保存。健康人外周血由廣州市血液中心提供。RPMI-1640培養(yǎng)基和胎牛血清均購自Gibco。EpiTect Plus DNA Bisulfite Kit購自 Qiagen。凝膠純化試劑盒購自Thermo。
2 方法
2.1 細胞分離培養(yǎng) Ficoll Hypaque密度梯度離心法分離PBMC,10%胎牛血清及1%青、鏈霉素雙抗的RPMI-1640培養(yǎng)基,37℃、5%CO2條件下培養(yǎng)。
2.2 登革病毒感染 DENV2進行擴增定量,病毒滴度為1×107pfu/L,以感染復數(shù)(multiplicity of infection,MOI)為1處理細胞,病毒吸附2 h后棄上清,加入含4%胎牛血清的培養(yǎng)基,于37℃、5%CO2條件下孵育6 h或12 h。
2.3 DNA甲基化轉(zhuǎn)化處理 取登革病毒感染0 h、6 h和12 h的細胞,分別提取DNA,使用EpiTect Plus DNA Bisulfite Kit對各組DNA進行甲基化轉(zhuǎn)化。
2.4 PCR擴增 對各組甲基化轉(zhuǎn)化后的DNA進行擴增,設(shè)計亞硫酸氫鹽測序PCR甲基化檢測引物,正義鏈 5′-AGGGTTTTATATATAAATTAGTTAGTGGTTTAGAAGA-3′,反義鏈:5′-TATAATTACTTCTCTCCCTCTTAACTAATCCTC-3′,擴增產(chǎn)物大小為 353 bp(-294 bp~+58 bp)。PCR反應(yīng)后取3 μL PCR產(chǎn)物進行2%瓊脂糖凝膠電泳分析。對凝膠電泳后的各組DNA片段進行純化回收。
2.5 連接 目的片段 DNA 2 μL(約 150 ng),PMD19-T 載體(TaKaRa)1 μL(約50 ng),T4 快速連接酶 1 μL,10× Buffer用滅菌去離子水補足10 μL,22℃連接10 min。
2.6 轉(zhuǎn)化 取連接產(chǎn)物加到50 μL T1感受態(tài)細胞中,混勻,冰浴30 min;將上述轉(zhuǎn)化液置于42℃水浴30 s,取出后立即置于冰浴中放置2~3 min;向其中加入900 μL 37℃預熱的LB培養(yǎng)基(不含抗生素),150 r/min、37℃振蕩培養(yǎng)45 min;2 500 r/min離心5 min,將上清液吸走,留 100 μL混勻菌液,加到含AMP抗生素LB固體瓊脂培養(yǎng)基上(抗生素終濃度50 mg/L),用無菌的彎頭玻棒輕輕將細胞均勻涂開;待平板表面干燥后,倒置平板,37℃培養(yǎng)12~16 h。
2.7 基因測序 挑取上述平板上的單菌落,每組挑選5個克隆送中美泰和生物技術(shù)(北京)有限公司測序。
3 統(tǒng)計學處理
甲基化率以每組擴增序列發(fā)生甲基化的位點數(shù)占總甲基化位點數(shù)的百分率表示。各組間甲基化率差異的比較采用 χ2檢驗。以P<0.05為差異有統(tǒng)計學意義。
結(jié) 果
1 擴增序列
通過在線工具(http://www.urogene.org/methprimer/index1.html)預測甲基化位點,人工設(shè)計檢測引物。如圖1所示,箭頭所指為轉(zhuǎn)錄起始位點(transcriptional start site,TSS),擴增序列為(-294 bp~+58 bp),覆蓋11個甲基化位點。
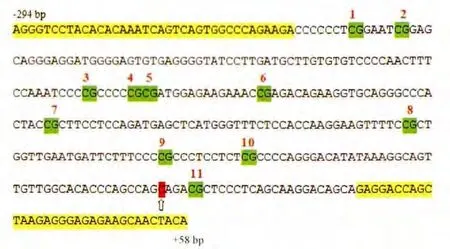
Figure 1.The sequence of PCR product.The sequence was from-294 bp to+58 bp,including eleven CpG sites.The arrow indicated the transcriptional start site(TSS).圖1 擴增序列
2 瓊脂糖凝膠電泳鑒定各樣本擴增產(chǎn)物
PCR反應(yīng)結(jié)束后取各樣本3 μL PCR產(chǎn)物進行2%瓊脂糖凝膠電泳分析,結(jié)果顯示擴增產(chǎn)物的大小與理論相符合,見圖2。
3 硫化修飾后的TNF-α基因啟動子區(qū)域部分互補序列的測序結(jié)果
挑取上述平板上的單菌落,每組挑選5個克隆送測序公司測序。序列中未甲基化的C脫氨基變成U,而甲基化的C保持不變,見圖3。
4 TNF-α基因啟動子區(qū)域甲基化水平變化
基因測序結(jié)果顯示,TNF-α基因啟動子區(qū)域-294 bp到+58 bp區(qū)域內(nèi),11個甲基化位點中,DENV2感染PBMC 0 h和6 h有2個位點發(fā)生甲基化,感染12 h有6個甲基化位點發(fā)生甲基化,甲基化水平呈增高趨勢,見圖4。
感染0 h、6 h和12 h平均甲基化率分別為10.3%、12.1%和25.5%。0 h與12 h及6 h與12 h之間的甲基化率差異有統(tǒng)計學意義(P<0.05)。
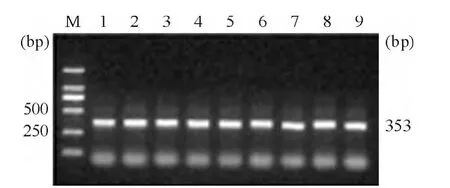
Figure 2.Identification of PCR product of each sample by agarose gel electrophoresis analysis.The size of PCR product was 353 bp,from-294 bp to+58 bp.The PCR products were consistent with the theoretical size.M:DL2000 DNA marker;1~9:PCR products of the samples.圖2 瓊脂糖凝膠電泳鑒定各樣本擴增產(chǎn)物
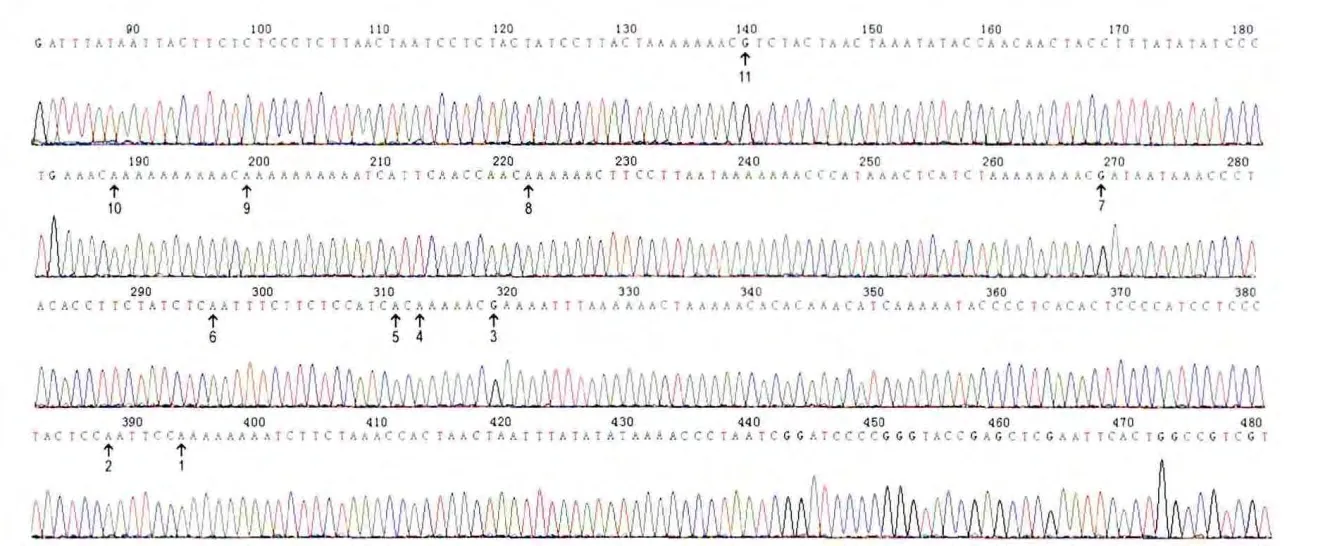
Figure 3.Partial result of complementary sequence of bisulfite-converted TNF-α promoter region.Me-thylated loci were G and unmethylated loci were A in the complementary sequence because methylated C remained unchanged and unmethylated C turned into U,which was combined with A.圖3 硫化修飾的TNF-α啟動子區(qū)域部分互補序列測序結(jié)果
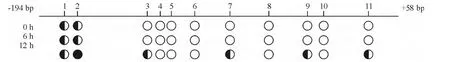
Figure 4.The changes of DNA methy-lation at CpG sites in the promoter region of TNF-α gene.The first and second sites were methylated at 0 h and 6 h,and the first,second,third,seventh,ninth and eleventh sites were methylated at 12 h.●:≥80%methylation at a given site in the sequencing of five individual clones;■○:between 20%and 80%methylation at a given site;○:≤20%methylation at a given site.圖4 TNF-α基因啟動子區(qū)域甲基化水平
討 論
目前登革熱的發(fā)病機制尚未完全明確,細胞因子風暴為重癥登革熱發(fā)病機制的相關(guān)假說之一[6],登革病毒感染后多種細胞因子的水平會受到影響。Bozza等[7]的研究證實在重癥登革熱患者中IL-1、IL-4、IL-6、IL-13、IL-7 和 IFN-γ 等明顯增高,IL-1β、IL-8和TNF-α與血小板的減少有明顯的相關(guān)性。Wati等[8]發(fā)現(xiàn)TNF-α能夠通過多種機制誘導登革病毒感染的細胞發(fā)生凋亡。Jaiyen等[2]提出 DENV引起PBMC凋亡,除了病毒本身的毒性作用,也與產(chǎn)生的促炎因子TNF-α和IL-1β有關(guān);他們還發(fā)現(xiàn)DHF患者TNF-α顯著高于DF患者,提示TNF-α在重癥登革病毒感染中起重要作用。TNF-α基因?qū)儆谠绶磻?yīng)基因范疇,TNF-α蛋白呈現(xiàn)快速產(chǎn)生、早期分泌、快速下降的規(guī)律。有研究證實LPS刺激THP-1細胞TNF-α分泌在4 h達峰之后下降[9]。本課題組之前的研究發(fā)現(xiàn)正常人PBMC被DENV2感染0 h、6 h和12 h,隨時間增加 TNF-α 的產(chǎn)生減少[3]。這似乎提示了病毒初次感染與重癥感染的不同,可能是機體在初次病毒感染早期對自身的一種保護機制,避免重癥登革熱的發(fā)生。
TNF-α的表達調(diào)控受多種因素的影響,DNA甲基化修飾在這個過程中起重要作用。本研究采用亞硫酸氫鹽測序PCR法檢測TNF-α基因啟動子區(qū)的甲基化狀態(tài),我們的結(jié)果表明病毒感染12 h TNF-α啟動子區(qū)域11個位點發(fā)生甲基化的水平明顯增加,感染6 h與0 h相比甲基化水平相當。我們之前的報道[3],DENV2 感染 PBMC 0 h、6 h 和 12 h,隨時間增加TNF-α的產(chǎn)生減少。這種不同可能是由多因素共同調(diào)控 TNF-α的表達引起的。郭堯等[5]發(fā)現(xiàn)THP-1細胞在LPS刺激之前TNF-α啟動子區(qū)域的CpG雙核苷酸均處于甲基化狀態(tài),而本實驗中PBMC在病毒感染0 h時就出現(xiàn)多個位點的去甲基化,這可能是細胞種類差異導致的不同。
本研究證明了PBMC在DENV2感染早期存在TNF-α基因甲基化水平的改變。甲基化能夠調(diào)控蛋白的表達,且TNF-α蛋白水平與登革熱病情的嚴重程度相關(guān),這可能為治療或緩解登革病毒感染提供新的思路。
[1] Bhatt S,Gething PW,Brady OJ,et al.The global distribution and burden of dengue[J].Nature,2013,496(7446):504-507.
[2] Jaiyen Y,Masrinoul P,Kalayanarooj S,et al.Characteristics of dengue virus-infected peripheral blood mononuclear cell death that correlates with the severity of illness[J].Microbiol Immunol,2009,53(8):442-450.
[3] Qi Y,Li Y,Zhang L,et al.MicroRNA expression profiling and bioinformatic analysis of dengue virus-infected peripheral blood mononuclear cells[J].Mol Med Rep,2013,7(3):791-798.
[4] Bergman Y,Cedar H.DNA methylation dynamics in health and disease[J].Nat Struct Mol Biol,2013,20(3):274-281.
[5] 郭 堯,楊智勇,王春友.啟動子區(qū)域甲基化狀態(tài)對THP-1細胞腫瘤壞死因子α分泌的影響[J].中國病理生理雜志,2011,27(6):1206-1209.
[6] 黃俊琪.登革熱發(fā)病機制的研究進展[J].實用醫(yī)學雜志,2011,27(19):3464-3465.
[7] Bozza FA,Cruz OG,Zagne SM,et al.Multiplex cytokine profile from dengue patients:MIP-1β and IFN-γ as predictive factors for severity[J].BMC Infect Dis,2008,8:86.
[8] Wati S,Rawlinson SM,Ivanov RA,et al.Tumour necrosis factor alpha(TNF-α)stimulation of cells with established dengue virus type 2 infection induces cell death that is accompanied by a reduced ability of TNF-α to activate nuclear factor κB and reduced sphingosine kinase-1 activity[J].J Gen Virol,2011,92(Pt 4):807-818.
[9] 郭 堯.啟動子區(qū)域甲基化狀態(tài)與炎癥細胞TNF-α分泌關(guān)系及調(diào)控因素的研究[D].武漢:華中科技大學,2011.
猜你喜歡
發(fā)明與創(chuàng)新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農(nóng)墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經(jīng)大學學報(2015年3期)2015-12-10 03:49:15
