Orai2參與了HUVEC外鈣敏感受體介導的Ca2+內流和NO生成
2014-11-27 09:23:46王臘梅龐麗娟孫志萍
基礎醫學與臨床 2014年5期
王臘梅,鐘 華,趙 慧,王 靜,龐麗娟,孫志萍,何 芳*
(新疆石河子大學 醫學院 1.新疆地方病與民族高發病教育部重點實驗室;2.病理生理教研室;3.病理教研室; 4.醫學機能實驗中心;新疆 石河子 832002)
研究論文
Orai2參與了HUVEC外鈣敏感受體介導的Ca2+內流和NO生成
王臘梅1,2,鐘 華1,2,趙 慧1,2,王 靜1,2,龐麗娟1,3,孫志萍1,4,何 芳1,2*
(新疆石河子大學 醫學院 1.新疆地方病與民族高發病教育部重點實驗室;2.病理生理教研室;3.病理教研室; 4.醫學機能實驗中心;新疆 石河子 832002)
目的研究鈣釋放激活的鈣調素2(Orai2)在人臍靜脈內皮細胞(HUVEC)細胞外鈣敏感受體(CaR)介導Ca2+內流和一氧化氮(NO)生成中的作用。方法1)用轉染技術將構建的Orai2干擾質粒(Orai2shRNA)轉染入HUVEC,用實時定量RT-PCR和Western blot檢測Orai2 mRNA和蛋白的表達。2)取2~5代HUVEC,分別以精胺激活CaR(鈣池操縱性鈣通道(SOC)和受體操縱性鈣通道(ROC)均激活)、ROC模擬劑12-O-十四烷酰佛波醋酸酯-13(TPA)+CaR負性變構調節劑Calhex231(激活ROC及阻斷SOC)、蛋白激酶C(PKC)抑制劑Ro31-8220、經典型PKCs和PKCμ抑制劑Go6967(阻斷ROC及激活SOC)為細胞模型。實驗分為3組:特異性質粒轉染組(Orai2shRNA組),未轉染組(空白對照組),空質粒組(vehicle組),用熒光探針Fura-2/AM、DAF-FM負載方法同步檢測[Ca2+]i和NO的生成。結果1)與control組相比,shOrai2-71干擾后,Orai2 mRNA和蛋白的表達均明顯降低,抑制率分別為75.75%和70.58%(Plt;0.05)。2)與對照組和空質粒組相比,在4種不同處理因素作用下,Orai2shRNA轉染組[Ca2+]i△ratio值和NO凈熒光強度值均明顯降低(Plt;0.05)。結論Orai2參與了HUVEC中CaR經SOC和ROC激活介導的Ca2+內流和NO生成。
Orai2;一氧化氮;鈣離子;人臍靜脈內皮細胞
隨著鈣通道相關蛋白C-型瞬時型感受器電位通道 (canonical transient receptor potential channels,TRPCs)、基質相互作用分子(stromal interaction molecules,STIMs)、鈣釋放激活的鈣調素(calcium release-activated calcium modulator,Orais)的發現,其單個組件或復合物在不同組織細胞介導的生理和病理生理過程中作用已被進一步研究。Orai家族是鈣池操縱性鈣通道(store-operates cation channels, SOC)的構成蛋白,包括Orai1、Orai2和Orai3[1]。其中,Orai1可以同Orai2,Orai3,甚至可能與TRPCs形成同源/異源復合體介導鈣庫操縱的鈣內流(store operated calcium entry,SOCE)或調節受體操縱鈣內流(receptor-operated calcium entry, ROCE)[2],而由Orai2構成的SOCE或ROCE在不同組織細胞中生物學作用報道甚少。在人臍靜脈內皮細胞(human umbilical vein endothelial cells, HUVEC)中,鈣敏感受體(Ca-sensing receptor, CaR)激活介導了[Ca2+]i升高及一氧化氮(nitric oxide, NO)的生成過程[3-4],SOC和受體操縱性鈣通道(receptor-operated channels, ROC)以協同的方式參與了此過程,且TRPC1、STIM1和Orai1為上述過程的關鍵組件之一。本研究觀察Orai2基因沉默后對HUVEC中[Ca2+]i和NO生成影響,揭示Orai2在CaR介導[Ca2+]i和NO生成中作用和機制。
1 材料與方法
1.1 材料
健康孕婦剖宮產的新鮮臍帶(華中科技大學同濟醫學院附屬同濟醫院,經倫理道德委員會批準和個人知情同意)。
ECM培養基(Sciencell公司);蛋白酶抑制劑(Calbichem公司);兔抗人Orai2多克隆抗體、鼠抗β-Actin單克隆抗體(Santa Cruze公司);二抗(Protein Tech公司);ECL發光試劑盒(Thermo公司);反轉錄試劑盒與Real time RT-PCR試劑盒(TaKaRa公司);LipofectamineTM2000與Opti-MEM(Invitrogen公司);Fura-2/AM(Invitrogen公司);DAF-FM DA(NO熒光探針)(Beyotime公司);G418 (Biosharp公司)、去內毒素高純度質粒抽提試劑盒(Omega公司);shRNA(上海吉凱基因化學技術有限公司);引物(友名生物技術有限公司);其余均為國產分析純試劑。
1.2 方法
1.2.1 HUVEC的培養與鑒定:按本實驗室以前的方法培養HUVEC 并傳代[3],用細胞內Ⅷ因子相關抗原進行免疫細胞化學染色,鑒定HUVEC,取生長狀態良好的2~5代細胞用于實驗。
1.2.2Orai2基因的shRNA構建及轉染:1)按shRNA的設計原則,GenBan中人Orai2的cDNA序列(NM-032831),以其同源的編碼DNA序列部分設計、合成3條shRNA,分別命名為shOrai2-71、shOrai2-72和shOrai2-73,上述序列經BLAST軟件分析,與人類基因外顯子無同源性,排除對其他基因非特異性干擾,并與載體連接構建成重組質粒。2)轉染,細胞增殖至80%匯合時進行轉染,實驗分為未轉染組即空白對照組(control組)、空質粒組(vehicle組)和特異性質粒轉染組即實驗組(Orai2shRNA)。以6孔板的一個孔為例,首先取一無菌離心管加250 μL OPTI-MEM培養基和5 μL LipofectamineTM2000(Invitrogen),輕輕混勻。取另一無菌離心管加250 μL OPTI-MEM培養基和2 μg個質粒DNA,輕輕混勻。室溫下孵育5 min后,混勻以上2種復合物,室溫孵育20 min。最后將此復合物加入到含有1.5 mL OPTI-MEM的6孔板中,37 ℃培養4~6 h后換培養基繼續培養,孵育24~48 h后,檢測轉染效率。
1.2.3 實驗分組和處理:選取干擾效率最高的shOrai2-71為實驗組,其余兩組分別為未轉染組即空白對照組(control組)與空質粒組(vehicle組)。將3組細胞接種于圓形玻片上,待HUVEC細胞增殖匯合至80%左右時進行轉染,繼續培養48 h后分別與含精胺(2 mmol/L)+Ca2+(2 mmol/L)、含精胺(2 mmol/L)+Ca2+(2 mmol/L)+CaR的抑制劑Calhex231(1 μmol/L)+ROC模擬劑TPA(20 μmol/L)、含精胺(2 mmol/L)+Ca2+(2 mmol/L)+PKC抑制劑Ro31-8220(200 nmol/L)或+PKCs、PKCμ抑制劑Go6967(200 nmol/L)的ECM孵育20 min,將玻片取出放入灌流槽中測定加精胺刺激后[Ca2+]i△ratio值和NO熒光強度值的變化,每組實驗重復3次。
1.2.4 Real time RT-PCR檢測HUVEC中Orai2 mRNA的表達:按SYBR greenⅠRt-PCR試劑盒說明書操作。Orai2的上游引物:5′-GGTGTAGTGGGAGG TGAGGA-3′,下游引物:5′-TGGCTTTACCACCATGTC AA-3′。β-actin的上游引物:5′-ACGGTCAGGTCATC ACTATCG-3′,下游引物:5′-GGCATAGAGGTCTTT ACGGATG-3′。用Trizol法抽提細胞RNA,紫外分光光度儀分別在260 nm和280 nm波長下檢測計算提取的總RNA濃度及純度。取總RNA 2 μg反轉錄成cDNA,以此為模板以β-actin作為內參照進行Real-time PCR擴增,反應體系為25 μL,其中SYBR Premix Ex Tap12.5 μL,PCR Forward Primer 0.5 μL,PCR Reverse Primer 0.5 μL,樣品cDNA 2.0 μL,滅菌蒸餾水9.5 μL。反應條件為預變性95 ℃,30 s;PCR反應(95 ℃ 5 s、60 ℃ 20 s),40個循環。隨之由電腦自動分析系統進行定量分析,每組實驗重復4次。
1.2.5 免疫印跡法檢測HUVEC中Orai2的蛋白表達及干擾效率:各轉染組轉染48 h后,用G418進行穩篩,預冷的PBS沖洗細胞3次,冰上加入含3 μL蛋白酶抑制劑的細胞裂解液300 μL,刮下細胞收集于離心管中。冰上靜置60 min,12 000r/min,離心15 min。取上清液棄沉淀。取5 μL上清液測蛋白質濃度并標化。SDS-PAGE電泳分離蛋白,轉膜,封閉,加入抗Orai2抗體,抗β-actin抗體置于4 ℃過夜。二抗室溫孵育1 h,化學發光試劑盒處理,顯影、定影,重復3次。
1.2.6 HUVEC中Ca2+濃度測定:參照文獻[5]測定HUVEC中Ca2+水平。通過鈣熒光成像系統IPA software進行數據分析,每組實驗重復3次。
1.2.7 HUVEC中NO含量的檢測:參照文獻[5]測定HUVEC中NO含量。通過鈣熒光成像系統IPA software進行數據分析,每組實驗重復3次。
1.3 統計學分析
2 結果
2.1 shRNA成功轉染HUVEC
細胞轉染FITC標記的shOrai2后,鏡下可見成功轉染的細胞內發出可反映轉染效率的綠色熒光。轉染48 h后加入G418穩篩后,可獲取90%以上的陽性克隆細胞(圖1)。
2.2 Orai2 mRNA的表達及干擾效率
轉染各組與control組相比,可見vehicle組mRNA表達無明顯變化,其余各轉染組mRNA的表達均有降低(n=4,Plt;0.05),尤其是shOrai2-71組中Orai2 mRNA表達顯著降低(抑制率為75.75%)(圖2)。
2.3 Orai2的蛋白表達及干擾效率
轉染各組與control組相比,可見vehicle組的蛋白表達無明顯變化,其余各轉染組蛋白的表達均有降低(n=3,Plt;0.05),尤其是shOrai2-71組中Orai2的蛋白表達顯著降低(抑制率為70.58%)(圖3)。
2.4不同處理因素對HUVEC中[Ca2+]i和NO生成的影響
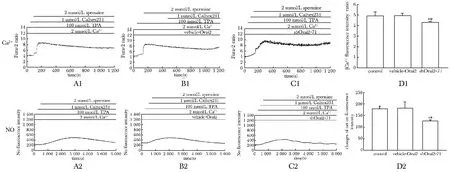
2.4.1Orai2基因沉默使精胺介導的HUVEC中[Ca2+]i和NO生成減少:與control組及vehicle組相比,實驗組[Ca2+]i△ratio值和NO凈熒光強度值明顯降低(n=3,Plt;0.05),vehicle組中[Ca2+]i△ratio值和NO凈熒光強度值無顯著差異(圖4)。

A.confocal microscopy showed the HUVEC profile in nature light; B.confocal microscopy showed FITC-labelled green fluorescence in HUVEC; C.merged image of A,B (scale bar=50 μm)
圖1化學熒光FITC標記的shOrai2轉染進入細胞
Fig1FITC-labelledshOrai2transfectedHUVEC(×20)

*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle group圖2 HUVEC中轉染組Orai2 mRNA的干擾效率Fig 2 Interference ratio of Oraiz mRNA and inhibition ratio after transfection in HUVEC(±s, n=4)
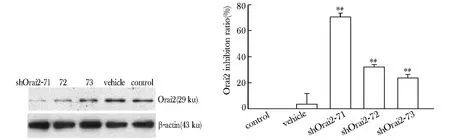
*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle group圖3 各轉染組HUVEC中Orai2蛋白的表達及抑制效率Fig 3 Expression of Orai2 protein examined by Western blot and inhibition ratio in HUVEC(±s, n=3)
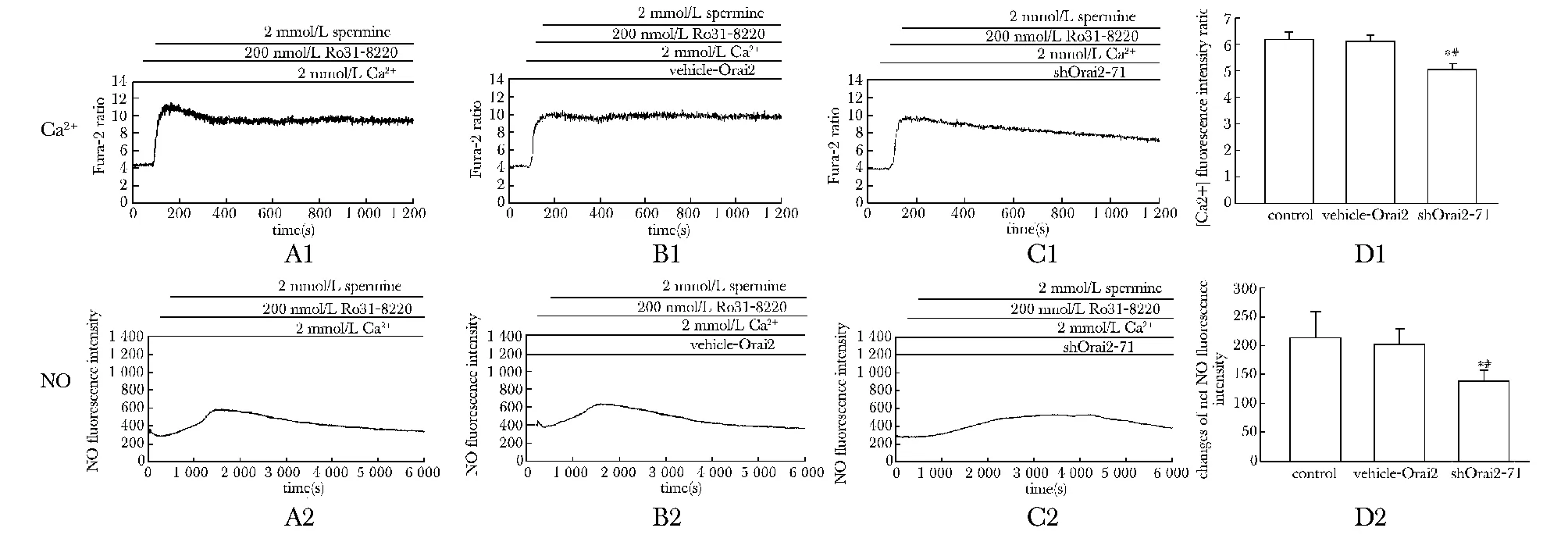
2.4.2Orai2基因沉默使TPA+Calhex231介導的HUVEC中[Ca2+]i和NO生成減少:與control組及vehicle組相比,實驗組中[Ca2+]i△ratio值和NO凈熒光強度值明顯降低(n=3,Plt;0.05),vehicle組中[Ca2+]i△ratio值和NO凈熒光強度值無顯著差異 (圖5)。
2.4.3Orai2基因沉默使Ro31-8220介導的HUVEC中[Ca2+]i和NO生成減少:與control組及vehicle組相比,實驗組中[Ca2+]i△ratio值和NO凈熒光強度值明顯降低(n=3,Plt;0.05),vehicle組中[Ca2+]i△ratio值和NO凈熒光強度值無顯著差異 (圖6)。
2.4.4Orai2基因沉默使Go6967介導的HUVEC中[Ca2+]i和NO生成減少:與control組及vehicle組相比,實驗組中[Ca2+]i△ratio值和NO凈熒光強度值明顯降低(n=3,Plt;0.05),vehicle組中[Ca2+]i△ratio值和NO凈熒光強度值無顯著差異 (圖7)。
3 討論
本研究前期實驗發現在HUVEC中CaR激活引發持續Ca2+內流是NO生成的重要條件,但參與Ca2+內流關鍵分子組件尚不清楚。應用RNA干擾(RNAi)技術,發現了兩個與SOC功能密切相關的通道蛋白:STIM1和Orai1。Orai1的同源蛋白Orai2和Orai3也參與構成SOC,但是這些通道具有與Orai1構成的通道不完全一致的生物物理特性[6]。在異源表達STIM1、Orai1、Orai2和Orai3的HEK293細胞中發現,所有Orai蛋白都能增加SOCE,且增強SOC的作用功效依次是Orai1gt;Orai2gt;Orai3,其中Orai3能部分代償Orai1敲除對細胞產生的影響[7]。對Orai2的研究表明其特點與Orai1相似,Orai2單獨表達時抑制HEK293細胞形成SOCE;當同STIM1聯合表達時,則可以增強SOCE。

*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle-Orai2 group圖4 CaR激動劑精胺刺激HUVEC后各轉染組中Ca2+和NO熒光強度動態變化
*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle-Orai2 group圖5 ROC模擬劑TPA+CaR負性變構調節劑Calhex231刺激HUVEC后各轉染組中Ca2+和NO熒光強度動態變化
*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle-Orai2 group圖6 PKC抑制劑Ro31-8220刺激HUVEC后各轉染組中Ca2+和NO熒光強度動態變化

*Plt;0.05 compared with control group; #Plt;0.05 compared with vehicle-Orai2 group圖7 經典型PKCs和PKCμ抑制劑Go6967刺激HUVEC后各轉染組中Ca2+和NO熒光強度動態變化
HUVEC中CaR為功能性表達,其在介導Ca2+內流、eNOS活性和NO生成中起著重要作用[8],且SOC與ROC是以協同方式參與CaR介導的Ca2+內流及NO生成。而鈣通道相關蛋白Orai2是否為其關鍵分子組件尚不清楚。研究首先采用脂質體法將設計好的shRNA轉染到HUVEC中,將抑制效率最高的shRNA轉染HUVEC,在保證沉默Orai2基因有效抑制Orai2表達前提下,以精胺與原代培養HUVEC共孵育激活CaR為細胞模型,證實了在HUVEC中沉默Orai2后對CaR介導的[Ca2+]i升高和NO生成均有抑制作用,此抑制效應由shRNA特異抑制Orai2基因表達所介導。而后又以ROC模擬劑TPA+CaR負性變構調節劑Calhex231、PKC抑制劑Ro31-8220,經典型PKCs和PKCμ抑制劑Go6967共孵育HUVEC為細胞模型,通過分別激活SOC或ROC最終證明在精胺刺激激活CaR介導[Ca2+]i和NO的生成過程中,Orai2既作為SOC的關鍵組件,又作為ROC的關鍵組件參與了CaR介導Ca2+內流及NO生成。
綜上所述,本研究證明了Orai2為CaR經SOC及ROC途徑介導鈣內流和NO生成的關鍵組件之一。這不僅從SOC及ROC角度揭示了CaR在血管內皮細胞中部分功能及作用機制,且進一步發現了參與上述過程的關鍵分子組件。本研究將有助于以鈣信號系統障礙的多病理機制為靶點,為心腦血管疾病防治提供一條新思路。
志謝:衷心感謝華中科技大學同濟醫學院病理生理學系/衛生部呼吸疾病重點實驗室提供實驗條件和技術指導。
[1] Ji W, Xu P, Li Z,etal. Functional stoichiometry of the unitary calcium-release-activated calcium channel[J]. Proc Natl Acad Sci USA, 2008, 105: 13668-13673.
[2] Liao Y, Plummer NW, George MD,etal. A role for Orai in TRPC-mediated Ca2+entry suggests that a TRPC: Orai complex may mediate store and receptor operated Ca2+entry[J]. Proc Natl Acad Sci USA, 2009,106: 3202-3206.
[3] 王振煥, 胡清華, 鐘華, 等. 小凹蛋白-1在臍靜脈內皮細胞CaR介導NO生成中的作用和機制[J]. 中國病理生理雜志, 2011, 27: 934-938.
[4] 王振煥, 胡清華, 鐘 華, 等. 小凹蛋白-1下調人臍靜脈內皮細胞外鈣敏感受體介導的鈣內流[J]. 生理學報, 2011, 63: 39-47.
[5] Ziegelstein RC, Xiong Y, He C,etal. Expression of a functional extracellular calcium-sensing receptor in human aortic endothelial cells[J]. Biochem Biophys Res Commun, 2006, 342: 153-163.
[6] Lis A, Peinelt C, Beck A,etal. CRACM1, CRACM2, and CRACM3 are store-operated Ca2+channels with distinct functional properties [J]. Curr Biol, 2007, 17: 794-800.
[7] Mercer JC, Dehaven WI, Smyth JT,etal. Large store-operated calcium selective currents due to co-expression of Orail or Orai2 with the intracellular calcium sensor,Stiml[J]. J Biol Chem, 2006, 281: 24979-24990.
[8] 梁霄, 羅小林, 鐘華, 等. 小干擾RNA沉默細胞外鈣敏感受體抑制人臍靜脈內皮細胞鈣內流和NO生成[J]. 生理學報, 2012, 64: 289-295.
Orai2 participates in CaR mediated Ca2+entryand NO generation in human umbilical vein endothelial cells
WANG La-mei1,2, ZHONG Hua1,2, ZHAO Hui1,2, WANG Jing1,2, PANG Li-juan1,3,SUN Zhi-ping1,4, HE Fang1,2*
(1.Key Laboratory of Xinjiang Endemic and Ethnic Diseases, Ministry of Education; 2.Dept. of Pathophysiology;3.Dept. of Physiology; 4.Centre of Medical Functional Experiments, Medical College of Shihezi University, Shihezi 832002, China)
ObjectiveTo study the role of calcium release-activated calcium modulator 2(Orai2) in extracellular Ca2+-sensing receptor (CaR)-induced extracellular Ca2+influx and the production of nitric oxide (NO) in human umbilical vein endothelial cells (HUVEC).Methods1) We silenced the expression of Orai2 genes in HUVEC by transfection constructed Orai2 RNA interference plasmids. The expression of Orai2 protein and mRNA levels were determined by Western blotting and real time RT-PCR respectively. 2) The second to fifth passage of HUVEC were
incubated with CaR agonist spermine(activating store-operates cation channels (SOC) and examined for receptor-operated channels (ROC)), CaR negative allosteric modulator Calhex231 and ROC analogue TPA , protein kinase C (PKC) inhibitor Ro31-8220, PKCs and PKCμ inhibitor Go6967.Those cell models were divided into three groups: Orai2-71 short hairpin RNA group (Orai2shRNA); control group and vehicle group. Intracellular Ca2+concentration was detected and the production of NO was determined by DAF-FM.Results1)shRNA targeted at the Orai2 genes decreased Orai2 protein and mRNA levels by 70.58% and 75.75% respectively (Plt;0.05). (2)[Ca2+]iratio and the net NO fluorescence intensity values of Orai2shRNA group in four different treatments were significantly reduced (Plt;0.05).ConclusionsOrai2 participates in CaR-mediated Ca2+influx and NO production through SOC and ROC activation in HUVEC.
Orai2; nitric oxide; Ca2+; human umbilical vein endothelial cells
2013-08-26
2013-11-25
國家自然科學基金(31160239,81160018)
*通信作者(correspondingauthor): fangf2002shz@126.com
1001-6325(2014)05-0595-07
R363
A
