二烯丙基二硫活化NADPH氧化酶誘導人白血病K562細胞凋亡
2014-12-07 03:43:32伍尤華李林蔚
中國藥理學通報 2014年8期
關鍵詞:檢測
易 嵐,伍尤華,譚 暉,何 潔,李林蔚,單 健,蘇 琦
(南華大學1.腫瘤研究所、2.藥學與生物科學學院、3.附屬第一醫院腫瘤科,湖南 衡陽 421001)
誘導腫瘤細胞凋亡已經成為治療腫瘤的新方法之一[1]。研究表明[2],眾多的細胞因子和藥物具有誘導腫瘤細胞凋亡的能力。其中,大蒜及其烯丙基硫化物的抗癌作用正日益受到關注。研究顯示,二烯丙基二硫(diallyl disulfide,DADS),大蒜主要有效成分之一,可誘導胃癌、肺癌、結腸癌等多種腫瘤細胞凋亡[3]。我們前期工作已經證實,DADS不僅可誘導人結腸癌等細胞分化與凋亡,而且可誘導人白血病細胞分化與凋亡[4]。DADS對人白血病細胞具有雙效作用,小劑量DADS(<1.25 mg·L-1)誘導人白血病細胞分化,其機制與抑制JAK1/STAT3的酪氨酸激酶活性、下調STAT3與Myc核內表達水平,提高組蛋白乙酰化水平及p21WAF1表達上調有關[5]。而中等劑量 DADS(>1.25 mg·L-1)可誘導人白血病細胞凋亡,其機制與G2/M期阻滯,活性氧,抑制 ERK 和激活 p38,下調 Bag-1、Bcl-2,上調Fas-L、Bax 等有關[6-7]。但具體機制尚不清楚。
在前期工作的基礎上,我們應用蛋白質組學技術篩選出7個與DADS誘導人白血病細胞凋亡的相關的差異蛋白,其中Ras-related C3 botulinum toxin substrate 2(Rac2)表達明顯上調[8],Rac2 是 NADPH氧化酶的6個亞基之一,是NADPH氧化酶活化必不可少的。NADPH氧化酶的激活受高度調控,在此過程中,Rac2與 p67phox相互作用起著關鍵作用[9]。NADPH氧化酶是誘導產生ROS的主要來源之一,而ROS可觸發多種信號轉導途徑,引起細胞凋亡。本研究在前期工作的基礎上,研究DADS活化NADPH氧化酶,通過活性氧途徑誘導人白血病K562細胞凋亡,進一步闡明白血病的發病機制,為白血病的誘導凋亡治療提供新思路。
1 材料與方法
1.1 藥品及試劑 DADS為Fluka公司產品,小牛血清是杭州四季青生物工程公司產品。BCA蛋白定量試劑盒為Pierce公司產品。RNA抽提試劑盒Ⅱ為Omega公司產品,TRIzol試劑為Invitrogen Life Technologies產品。免疫共沉淀試劑盒為Santa Cruz公司產品。2',7'-二氯氫化熒光素二脂(DCFHDA)、phorbol myristate acetate(PMA)、diphenyleneiodonium(DPI)、Rac2、p67phox等試劑為 Santa Cruz公司產品。二抗為Abcam Biotechnology公司產品。
1.2 方法
1.2.1 細胞培養 細胞培養采用常規培養,培養液為含10%小牛血清的RPMI 1640,在37℃,飽和濕度,5%CO2孵箱中培養,每3~4 d換液傳代。取對數生長期細胞用于實驗,DADS用培養基稀釋到所需濃度。
1.2.2 MTT檢測 用含10%胎小牛血清的培養液配成單個細胞懸液,以每孔103~104個細胞接種到96孔板,每孔體積100 μl,設3個復孔。加入處理因素并設0對照、空白對照、溶酶對照和不同濃度DADS處理組。在37℃,飽和濕度,5%CO2孵箱中常規培養,繼續培養72 h。于結束前4 h除對照組外,每孔加 MTT溶液(5 g·L-1用 PBS配制,pH=7.4)20 μl,繼續孵育 4 h,終止培養。離心后再吸棄孔內培養上清液。每孔加150 μl DMSO,振蕩10 min,充分融解結晶物。酶聯免疫檢測儀上測各孔吸光度,檢測波長570 nm,按下列公式計算抑制率(IR%)。
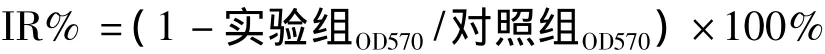
1.2.3 Real-time PCR引物合成 利用 Primer Premier 5.0軟件進行引物設計,送交寶生物工程大連有限公司合成。實時定量PCR使用的管家基金(β-actin)以及NADPH氧化酶的6個亞基引物分別為如下所示。β-actin正義引物序列為5'-CCTGTACGCCAACACAGTGC-3',反義引物序列為:5'-ATACTCCTGCTTGCTGATCC-3',產物長度為211bp;NCF2正義引物序列為:5'-GAAGAAGGGCAATGATAACTGG-3',反義引物序列為:5'-TAGGAGGAGCTGGGATGTCG-3',產物長度為155bp;RAC2正義引物序列為:5'-TCATCTGCTTCTCCCTCGT-3',反義引物序列為:5'-GATGGTGTCCTTGTCGTCC-3',產物長度為143bp;CYBB正義引物序列為:5'-CAAGATGCGTGGAAACTACCT-3',反義引物序列為:5'-TGACTTGAGAATGGATGCGAA-3',產物長度為138bp;CYBA正義引物序列為:5'-ATTGTGGCGGGCGTGTTT-3',反義引物序列為:5'-CACGGCGGTCATGTACTTCTGT-3',產物長度為 102bp;NCF1正義序列為:5'-ACCCTGAGCCCAACTATGC-3',反義序列為:5'-GGACGGGAAGTAGCCTGTG-3',產物長度為170bp;NCF4正義引物序列為:5'-CAGCCTGATGCCTCCTTACTC-3',反義引物序列為:5'-TCTGGAACTCCCGCCTTGT-3',產物長度為120bp。按照Total RNA KitⅡ說明書提取細胞總RNA,用紫外分光光度計測定RNA的純度和濃度。cDNA合成:將2 μg的總RNA 72℃ 10 min預變性,冰上冷卻3 min后,按RT試劑盒說明書依次加入各試劑,建立一個總反應體積20 μl的體系,42℃90 min,冰上冷卻2 min,-20℃保存備用。Realtime PCR擴增:將標準曲線樣品和待測樣品分別加入到Real-time PCR反應液中,進行Real-time PCR擴增和檢測。幾個梯度稀釋的DNA模板以及所有cDNA樣品分別配置總體積為25 μl反應體系,置于Real-time PCR儀上進行PCR反應。每個樣品的目的基因濃度除以其管家基因的濃度,即為此樣品此基因的校正后的相對含量。
1.2.4 細胞內活性氧檢測 收集細胞,用Hanks液洗滌細胞 2 次,加入終濃度為 10 μmol·L-1的DCFH-DA 400 μl,充分混勻,在37℃條件下避光反應50 min,用Hanks液洗滌細胞3次以去除細胞外熒光劑,用500 μl Hank's液重懸細胞,37℃條件下水浴10 min,流式細胞儀分析熒光強度,激發波長488 nm,發射波長530 nm。
1.2.5 Western blot檢測 細胞裂解:分別收集各組細胞,冰PBS洗兩次,每1×107個細胞濃度加入100 ~150 μl裂解液(100 mmol·L-1NaCl,10 mmol·L-1Tris-HCl pH 7.6,1 mmol·L-1EDTA pH 8.0,1 mg·L-1Aprotinin,100 mg·L-1PMSF),冰上裂解1 h,4℃,12 000 r·min-1離心 10 min,上清液即為細胞總蛋白。根據Bradford法進行蛋白含量的測定。蛋白變性后經10%SDS-聚丙烯酰胺凝膠電泳后轉移至PVDF膜上,用含5%牛血清白蛋白的TBST(Tris-HCL 20 mmol·L-1,NaCl 137 mmol·L-1含0.1%Tween-20)封閉1 h,用特異性一抗4℃孵育過夜,TBST洗3次,每次5 min,相應的二抗孵育1 h,TBST洗3次,每次5 min,化學發光劑檢測蛋白質印跡,薄層掃描儀測定印跡區帶的光密度值。
1.2.6 免疫共沉淀 按照Seize Classic Mammalian Immunoprecipitation試劑盒說明書進行,將得到抗體復合物進行SDS-PAGE電泳,再進行Western blot檢測。
1.3 統計學處理 采用SPSS 12.0統計分析軟件,Student't-test法進行差異顯著性檢驗,數值用±s表示,用Sigma Plot10.0軟件進行繪圖。
2 結果
2.1 MTT檢測DADS對K562細胞的增殖抑制作用 如 Tab 1 所示,2.5、5.0、10.0、20.0、40.0 mg·L-1DADS處理人白血病K562細胞,抑制率分別為32.48%、59.34%、66.42%、81.05%、87.09%,抑制率呈濃度依賴性增加(P<0.05)。Tween80對照組對細胞增殖無明顯抑制作用(P>0.05)。這些結果表明,DADS以劑量依賴性抑制K562細胞的生長。IC50值在 5.0 mg·L-1左右。
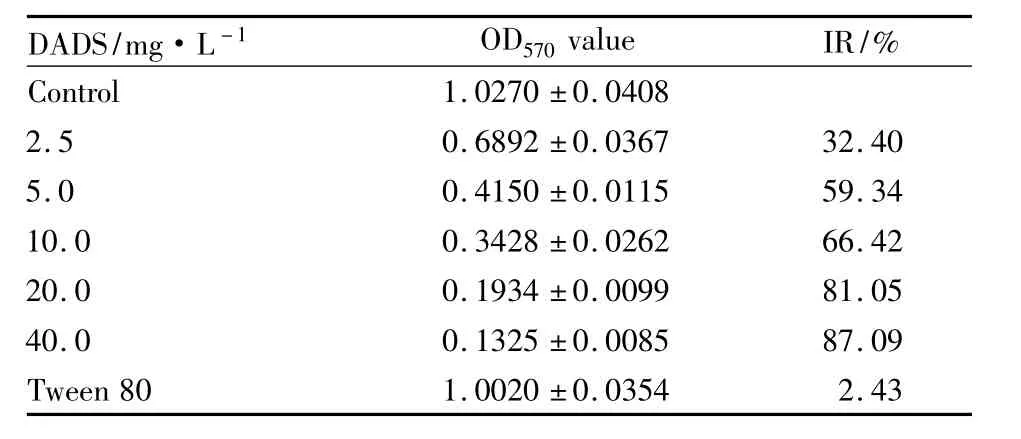
Tab 1 OD570value of different concentrations of DADS treated K562 cells for 48 hours
2.2 DADS誘導K562細胞凋亡過程中 NADPH氧化酶各亞基mRNA表達水平 Real-time PCR結果如Fig 1所示,6 mg·L-1DADS作用K562細胞6 h后,NADPH氧化酶復合物的6個亞基(包括CYBA、CYBB、NCF1、NCF2、NCF4 和 RAC2)相對定量的值分別為1.31×10-3±1.1×10-4、9.85 ×10-4±2.3 ×10-5、1.01 ×10-3±1.1 ×10-4、3.94 ×10-5±3.3 ×10-6、1.65 ×10-3±2.1 ×10-4、1.48 ×10-3±1.8×10-4,較處理前的 1.08 ×10-3±1.3×10-4、4.03×10-4±1.1×10-5、2.57 ×10-4±2.0 ×10-5、1.67×10-5±2.5 ×10-6、1.22 ×10-3±1.2 ×10-4、1.04×10-3±1.4 ×10-4,NADPH 氧化酶各亞基mRNA水平都明顯上調(P<0.05)。
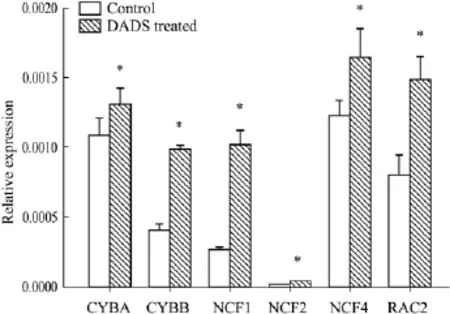
Fig 1 Real-time PCR analysis of mRNA levels of components of the NADPH oxidase in K562 cells treatedby 6 mg·L -1DADS for 6 hours
2.3 Rac2在DADS誘導K562細胞凋亡過程中表達上調 Rac2對NADPH氧化酶(NADPH oxidase)的6個亞基之一,Rac2是NADPH氧化酶活化是必不可少的。Western blot分析結果如Fig 2所示,與對照組相比,5.0、10.0 mg·L-1DADS 作用 K562 細胞24 h后,蛋白 Rac2的表達水平明顯上調(P<0.05)。
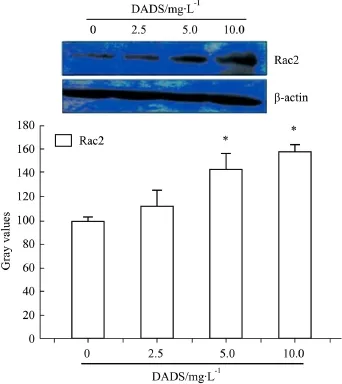
Fig 2 Western blot analysis of expression of Rac2 in K562 cellsinduced by different concentrations of DADS
2.4 免疫共沉淀檢測NADPH氧化酶亞基Rac2與p67phox結合 免疫共沉淀結果如Fig 3所示,通過用Anti-Rac2沉淀K562細胞的裂解蛋白,Western blot檢測到 p67phox存在。實驗結果表明在DADS誘導的 K562細胞凋亡過程中,有 Rac2和p67phox的結合,即存在NADPH氧化酶的活化。

Fig 3 Western blot analysis p67phox in K562 cells after subsided by Anti-Rac2
2.5 NADPH氧化酶激活劑和抑制劑對DADS誘導K562細胞凋亡的影響 我們采用NADPH氧化酶的激活劑PMA和NADPH氧化酶的黃素蛋白抑制劑DPI來進一步研究NADPH氧化酶對DADS誘導K562細胞凋亡的影響。
流式細胞術檢測凋亡細胞百分率結果如Fig 4所示,PMA單獨作用K562細胞24 h,其凋亡細胞百分率為(10.2±1.04)%,PMA與6 mg·L-1DADS聯合處理K562細胞24 h,凋亡細胞百分率為(40.5±2.03)%,與 DADS單獨處理組(30.9% ±1.34)%相比,PMA能明顯提高DADS誘導K562細胞凋亡的作用 (P<0.05),DPI單獨作用K562細胞24 h,凋亡細胞百分率為(3.35±0.76)%,可見,DPI并未能誘導K562細胞凋亡。DPI與6 mg·L-1DADS聯合處理K562細胞24 h,凋亡細胞百分率為(17.3±1.03)%,實驗結果表明DPI能抑制DADS誘導K562細胞凋亡(P<0.05)。
2.6 NADPH氧化酶的活化是DADS誘導K562細胞活性氧主要來源 流式細胞術檢測DCF的熒光強度結果如Fig 5所示,對照組、6 mg·L-1DADS處理K562細胞8 h組、PMA單獨處理組、DADS和PMA聯合處理組、DPI單獨處理組、DADS和DPI聯合處理組,流式細胞術檢測DCF的熒光強度分別為10.2±0.98、33.0±1.65、30.5±1.73、43.7±1.76、10.0±0.98、11.8 ±0.95,可見,PMA 能提高DADS誘導K562細胞活性氧的水平(P<0.05),而DPI明顯抑制了DADS作用的K562細胞活性氧的產生(P<0.05)。
3 討論
誘導腫瘤細胞凋亡已經成為治療腫瘤的一種新的可能性[10]。眾多的細胞因子和藥物都能誘導腫瘤細胞凋亡,其中,大蒜及其烯丙基硫化物的抗腫瘤作用已日益引起人們的關注。DADS作為大蒜提取物中最有效的成分,可以誘導多種腫瘤細胞包括人慢性髓細胞性白血病K562細胞系凋亡,但是其具體機制還有待于進一步研究。我們前期工作已經表明,低劑量(<1.25 mg·L-1)的DADS具有抗增殖作用,能誘導K562細胞分化[11]。本研究用中等濃度(2.5~20 mg·L-1)DADS來誘導K562細胞凋亡。在我們前期工作的基礎上,本研究進一步證實了DADS對K562細胞的生長抑制作用。MTT實驗結果表明,DADS以劑量依賴方式抑制K562細胞的活性。
本研究 Real-time PCR結果顯示,6 mg·L-1DADS作用K562細胞6h后,NADPH氧化酶復合物的6 個亞基(包括 CYBA、CYBB、NCF1、NCF2、NCF4和RAC2)的mRNA水平都明顯上調(P<0.05)。Rac2是NADPH氧化酶(NADPH oxidase)的6個亞基之一,Rac2的活化是NADPH氧化酶活化是必不可少的。Western blot分析結果表明,與對照組相比,5.0、10.0 mg·L-1DADS 作用人白血病 K562 細胞24 h后蛋白 Rac2的表達水平明顯上調(P<0.05)。這些研究結果表明DADS誘導K562細胞凋亡的過程中有NADPH氧化酶的活化。
有研究表明,NADPH氧化酶有6個亞基,在NADPH氧化酶活化過程中,Rac2與p67phox結合是NADPH氧化酶的活化非常關鍵的環節[12]。因此我們應用免疫共沉淀方法來檢測DADS誘導K562細胞凋亡的過程中Rac2與p67phox是否結合,進一步研究在DADS誘導人白血病K562細胞凋亡過程中,是否有NADPH氧化酶的活化。免疫共沉淀結果顯示,通過用Anti-Rac2沉淀K562細胞的裂解蛋白,Westernblot檢測到p67phox存在。實驗結果表明在DADS誘導的K562細胞凋亡過程中,有Rac2和p67phox的結合,也進一步說明有NADPH氧化酶的活化。
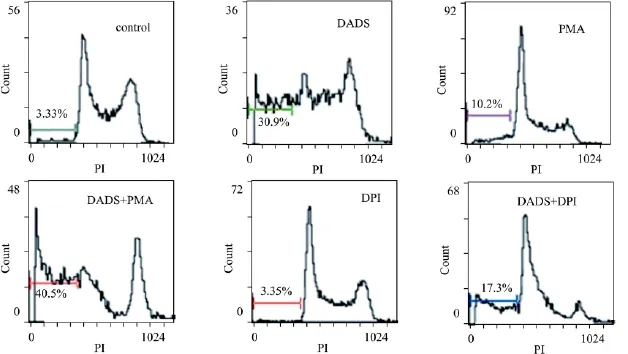
Fig 4 Flow cytometry analysis of percentage of apoptosis cells in K562 cells
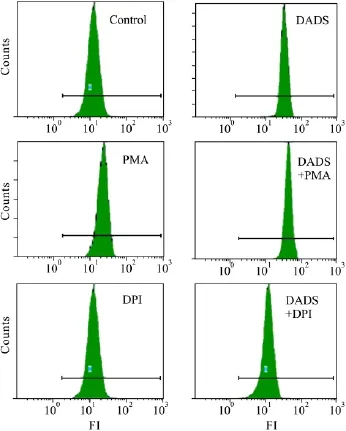
Fig 5 Flow cytometry analysis DCF fluorescence intensities of K562 cells
為進一步研究NADPH氧化酶與DADS誘導K562細胞凋亡的關系,我們采用NADPH氧化酶的激活劑PMA和NADPH氧化酶的黃素蛋白抑制劑DPI來研究NADPH氧化酶對DADS誘導K562細胞凋亡的影響。流式細胞術檢測凋亡細胞百分率結果顯示,PMA與DADS聯合處理組凋亡細胞百分率明顯高于與DADS單獨處理組,而DPI與DADS聯合處理組凋亡細胞百分率為明顯低于DADS單獨處理組。實驗結果表明,PMA能明顯提高DADS誘導K562細胞凋亡的作用,而DPI能抑制DADS誘導K562細胞凋亡。可見,NADPH氧化酶與DADS誘導K562細胞凋亡密切相關。
研究表明,DADS能誘導腫瘤細胞產生活性氧,產生的活性氧和腫瘤細胞的進一步的命運包括凋亡密切相關[13-14]。我們的前期研究結果顯示,DADS誘導K562細胞凋亡過程中活性氧水平明顯提高[7]。但是活性氧有多種來源,這些活性氧的來源并不確定,NADPH氧化酶的活化是活性氧的一個重要來源之一[15]。盡管我們的研究已經表明,在DADS誘導人白血病細胞凋亡的過程既有NADPH氧化酶的活化,又有活性氧的產生,但是NADPH氧化酶的活化是不是活性氧的主要來源還有待于進一步研究。因此我們用PMA及DPI預處理細胞,以便檢測DADS誘導K562細胞凋亡過程中活性氧的水平,以確定NADPH氧化酶的活化是否DADS誘導K562細胞凋亡過程中活性氧主要來源。流式細胞術檢測DCF的熒光強度結果顯示,PMA明顯提高了DADS誘導K562細胞活性氧的水平,而DPI預處理的細胞卻沒有活性氧的產生。這些結果表明,NADPH氧化酶是DADS誘導K562細胞凋亡過程中活性氧的主要來源。
NADPH氧化酶在多種腫瘤組織中的表達異于組織來源相同的正常組織,其催化產生的活性氧,可以誘導一系列細胞因子,調節細胞周期,調節腫瘤發生、發展和血管生成等等,這些為研究腫瘤發生和治療提供了新的思路。本研究結果不僅表明DADS誘導K562細胞凋亡的過程中有NADPH氧化酶的活化,而且證實了NADPH氧化酶的活化是DADS誘導K562細胞凋亡過程中活性氧產生的主要來源。因此,NADPH氧化酶的活化在DADS誘導人白血病K562細胞凋亡中起著重要作用。當然,DADS誘導人白血病細胞凋亡的確切機制還有待于進一步研究。
[1]Schulze-Bergkamen H,Krammer P H.Apoptosis in cancer-implications for therapy[J].Semin Oncol,2004,31(1):90 -119.
[2]Ferreira I C,Vaz J A,Vasconcelos M H,Martins A.Compounds from wild mushrooms with antitumor potential[J].Anticancer A-gents Med Chem,2010,10(5):424-36.
[3]Yi L,Su Q.Molecular mechanisms for the anti-cancer effects of diallyl disulfide[J].Food Chem Toxicol,2013,57:262 -70.
[4]Huang Y S,Xie N,Su Q,et al.Diallyl disulfide inhibits the proliferation of HT-29 human colon cancer cells by inducing differentially expressed genes[J].Mol Med Report,2011,4(3):553 -9.
[5]黃衛國,譚 暉,易 嵐,等.二烯丙基二硫上調p21、STAT1和CAMTA1誘導人白血病K562細胞分化[J].中國藥理學通報,2010,26(4):513-6.
[5]Huang W G,Tan H,Yi L,et al.Diallyl disulfide up-regulate p21,STAT1 and CAMTA1 induce differentiation of human leukemia HL-60 cells[J].Chin Pharmacol Bull,2010,26(4):513 -6.
[6]Tan H,Ling H,He J,et al.Inhibition of ERK and activation of p38 are involved in diallyl disulfide induced apoptosis of leukemia K562 cells[J].Arch Pharm Res,2008,31(6):786 -93.
[7]易 嵐,譚 暉,吉曉霞,等.DADS通過活性氧介導的JNK信號通路誘導人白血病K562細胞凋亡[J].中國藥理學通報,2013,29(4):473-7.
[7]Yi L,Tan H,Ji X X,et al.Diallyl disulfide induces apoptosis in human leukemia K562 cells through activation of JNK mediated by reactive oxygen[J].Chin Pharmacol Bull,2013,29(4):473 -7.
[8]Yi L,Zeng X,Tang H,et al.Proteomics analysis of apoptosis initiation induced by diallyl disulfide in human leukemia K562 cells[J].Chin J Cancer,2009,28(1):43 -8.
[9]Wen S,Zhu D,Huang P.Targeting cancer cell mitochondria as a therapeutic approach[J].Future Med Chem,2013,5(1):53-67.
[10]László K,István P.Selected aspects of cancer progression:metastasis,apoptosis and immune response[J].Apopt Cancer,2008,11:103-13.
[11]武明花,唐 黎,李利平,等.二烯丙基二硫對K562細胞的生長抑制和誘導分化作用[J].中華血液學雜志,2004,25(5):300-2.
[11]Wu M H,Tang L,Li L P,et al.The effect of Diallyl disulfide on cell growth inhibition and differentiationof K562[J].Chin J Hematol,2004,25(5):300 -2.
[12]Elnakish M T,Hassanain H H,Janssen P M,et al.Emerging role of oxidative stress in metabolic syndrome and cardiovascular diseases:important role of Rac/NADPH oxidase[J].J Pathol,2013,231(3):290-300.
[13]Wu W S.The signaling mechanism of ROS in tumor progression[J].Cancer Metastasis Rev,2006,25(4):695 -705.
[14]Kadara H,Tahara E,Kim H J,et al.Involvement of Rac in Fenretinide-Induced Apoptosis[J].Cancer Res,2008,68(11):4416-23.
[15]El-Benna J,Dang P M,Gougerot-Pocidalo M A.Priming of the neutrophil NADPH oxidase activation:role of p47phox phosphorylation and NOX2 mobilization to the plasma membrane[J].Semin Immunopathol,2008,30(3):279 -89.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
