HBx和c-Src對肝癌細胞HepG2上皮間質轉化的介導作用及其分子機制
2015-01-02 06:28:46呂高波寶雞市中心醫院肛腸外科寶雞7000西安交通大學第二附屬醫院腫瘤科通訊作者mailbluejackiestuxjtueducn
山西醫科大學學報 2015年12期
呂高波,趙 陽(寶雞市中心醫院肛腸外科,寶雞 7000;西安交通大學第二附屬醫院腫瘤科;通訊作者,E-mail:bluejackie@stu.xjtu.edu.cn)
最近幾年,越來越多的研究結果表明乙肝病毒的感染與肝細胞癌(hepatocellular carcinoma,HCC)的復發和轉移密切相關[1],但是乙肝病毒促進HCC的復發和轉移機制目前研究尚不清楚。乙肝病毒基因組有四種不同的開放閱讀框架,X基因所編碼的蛋白HBx與HCC發生和發展關系最為密切,HBx蛋白在HCC組織中的陽性率高達71.8%[2]。HBx本身是一種多功能蛋白,它具有促進正常肝細胞惡性轉化的作用,探明HBx對HCC侵襲轉移功能的影響機制具有十分重要的臨床及科學價值。伴隨著研究的深入,越來越多的HBx相關信號分子被發現,癌基因c-Src也被證明能夠被HBx所活化,而c-Src是重要的上皮間質轉化(epithelial-mesenchymal transition,EMT)調控因子[3]。針對 HBx的促侵襲轉移的分子機制,本研究通過干預HBx的表達以及應用c-Src抑制劑觀察其對肝癌細胞EMT的影響。
1 材料與方法
1.1 抗體和試劑
小鼠抗HBx單克隆抗體購自US Biological Biotechnology,小鼠抗 Vimentin,E-cadherin 和 β-catenin單克隆抗體均購自Santa Cruz Biotechnology,小鼠抗N-cadherin單克隆抗體購自Sigma公司,兔抗c-Src多克隆抗體購自 Calbiochem Biotechnology。C-Src特異性抑制劑PP2及其陰性對照PP3購自Calbiochem Biotechnology。
1.2 細胞系和細胞培養
HepG2—人類HCC細胞系(人乙肝病毒基因陰性)購自北京細胞中心(北京大學生物醫學中心),HepG2細胞培養于DMEM培養基中,培養基包含100 ml/L的胎牛血清(Sigma)、100 U/ml青霉素和100 μg/ml鏈霉素,培養箱內溫度維持在37℃,CO2含量為50 ml/L。
1.3 腺病毒載體的構建
腺病毒包裝試劑盒購自Bioscience公司,依據試劑盒操作指南,我們構建了表達HBx的腺病毒載體。HBx基因首先被插入到腺病毒包裝質粒pAdTrack-CMV上,從而構建了重組腺病毒質粒pAdTrack-CMVHBx,PmeI公司對 pAdTrack-CMV-HBx質粒進行測序,證實質粒重組成功。pAdTrack-CMV-HBx轉染進入BJ5183細胞,在BJ5183細胞中,pAdTrack-CMVHBx與pAdEasy-1進行同源重組,重組后再次測序,證實重組成功。將重組的質粒轉染進入293N細胞,包裝成腺病毒,腺病毒經過純化后,收集并保存于-80℃待用。對照腺病毒載體(Mock)的合成方式與HBx腺病毒載體合成方式相同。
1.4 轉染及細胞處理
HepG2細胞置于6孔板中培養,孵育24 h,使用無血清培養基稀釋純化的腺病毒,將不同稀釋濃度的腺病毒與細胞共孵育2 h,使用含血清培養基替換無血清培養基繼續培養HepG2細胞。轉染后的HepG2細胞在37℃,50 ml/L CO2的培養箱中培養7 d,檢測細胞內目標蛋白的表達。為了探索c-Src對EMT的影響,我們選擇 c-Src特異性抑制劑PP210 μg/ml處理 HepG2細胞24 h,觀察細胞內EMT相關信號分子的表達變化,應用PP3 10 μg/ml作為陰性對照。
1.5 免疫組化
將HepG2細胞置于蓋玻片上,爬片生長24 h,用100%丙酮或者100%乙醇和100%丙酮1:1混合液20 ℃固定細胞 20 min,應用抗 HBx(1∶100),E-cadherin(1∶100)、β-catenin(1∶100)、N-cadherin(1∶100)和vimentin(1∶100)一抗,于4℃孵育細胞過夜,PBS沖洗蓋玻片5 min×3次,應用生物素標記的二抗于37℃孵育30 min,PBS沖洗蓋玻片5 min×3次,顯色劑DAB顯色20 min,自來水沖洗終止反應,之后,蘇木素復染1 min,自來水沖洗30 s,封片保存并拍照。

圖1 HBx對HepG2細胞形態的影響Figure 1 Effect of HBx on the morphology of HepG2 cells
1.6 Western blot檢測
HepG2細胞應用含100 ml/L胎牛血清的DMEM培養基培養至90%融合,細胞裂解液裂解細胞,收集細胞裂解液,應用BCA試劑盒(購自Pierce Chemical公司)檢測蛋白濃度,記錄不同蛋白濃度,蛋白樣本煮沸15 min,使其變性,于-20℃保存。根據蛋白濃度調整上樣量,保證上樣蛋白量一致,經過SDS-PAGE電泳,蛋白印跡轉至NC膜(Millipore公司)上。NC膜于50 g/L脫脂牛奶中4℃孵育過夜,特異性一抗(anti-E-cadherin 1∶1 000,124 kD;anti-βcatenin 1∶1 000,92 kD;anti-N-cadherin 1∶1 000,130 kD;anti-vimentin 1∶1 000 58 kD;1∶1 000;anti-HBx 1∶1 000,17 kD)室溫孵育 NC 膜 2 h,TBST 洗膜 5 min×3次,室溫孵育辣根過氧化物酶標記的二抗2 h,TBST洗膜 5 min×3次,顯色液(購自 Pierce Chemical公司)顯色,壓片定影后拍照,計算灰度值。
1.7 統計學分析
2 結果
2.1 HBx可以改變HepG2細胞的形態
HBx腺病毒載體的轉染提高了HepG2細胞中的HBx表達水平,相比較轉染對照腺病毒的HepG2細胞,外源性HBx轉染的HepG2細胞形態發生了明顯的改變,細胞展現出了成纖維細胞樣形態變化,細胞呈梭形,軸向伸展延長,與周圍細胞松散分布(見圖1)。
2.2 HBx對HepG2細胞EMT標志物的影響
為了探索HBx與EMT之間的相關性,我們觀察了上皮細胞標志E-cadherin和β-catenin,間質細胞標志N-cadherin和Vimentin的表達。外源性HBx蛋白表達上調,顯著上調了上皮細胞標志E-cadherin的表達,下調了間質細胞標志N-cadherin和Vimentin的表達(圖2),這些結果符合EMT改變。
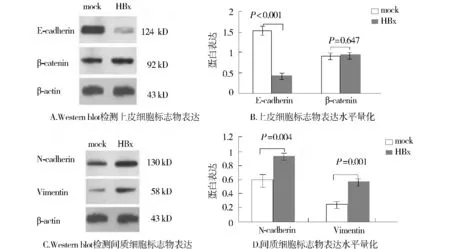
圖2 HBx介導HepG2細胞EMT標志物的表達Figure 2 HBx induces epithelial and mesenchymal markers expression in HepG2 cells
我們進一步應用免疫組織化學方法觀察了上皮細胞及間質細胞標志物在HepG2細胞中的表達特點,E-cadherin呈現為近膜分布,N-cadherin和Vimentin為細胞質分布,β-catenin在HBx轉染的細胞胞質和胞核中均有分布,在對照細胞中β-catenin集中分布在細胞質中。HBx顯著改變了HepG2細胞的形態,呈現出了松散分布和梭形改變的特點,下調了上皮細胞標志E-cadherin表達,上調了間質細胞標志物N-cadherin和Vimentin(見圖3)。
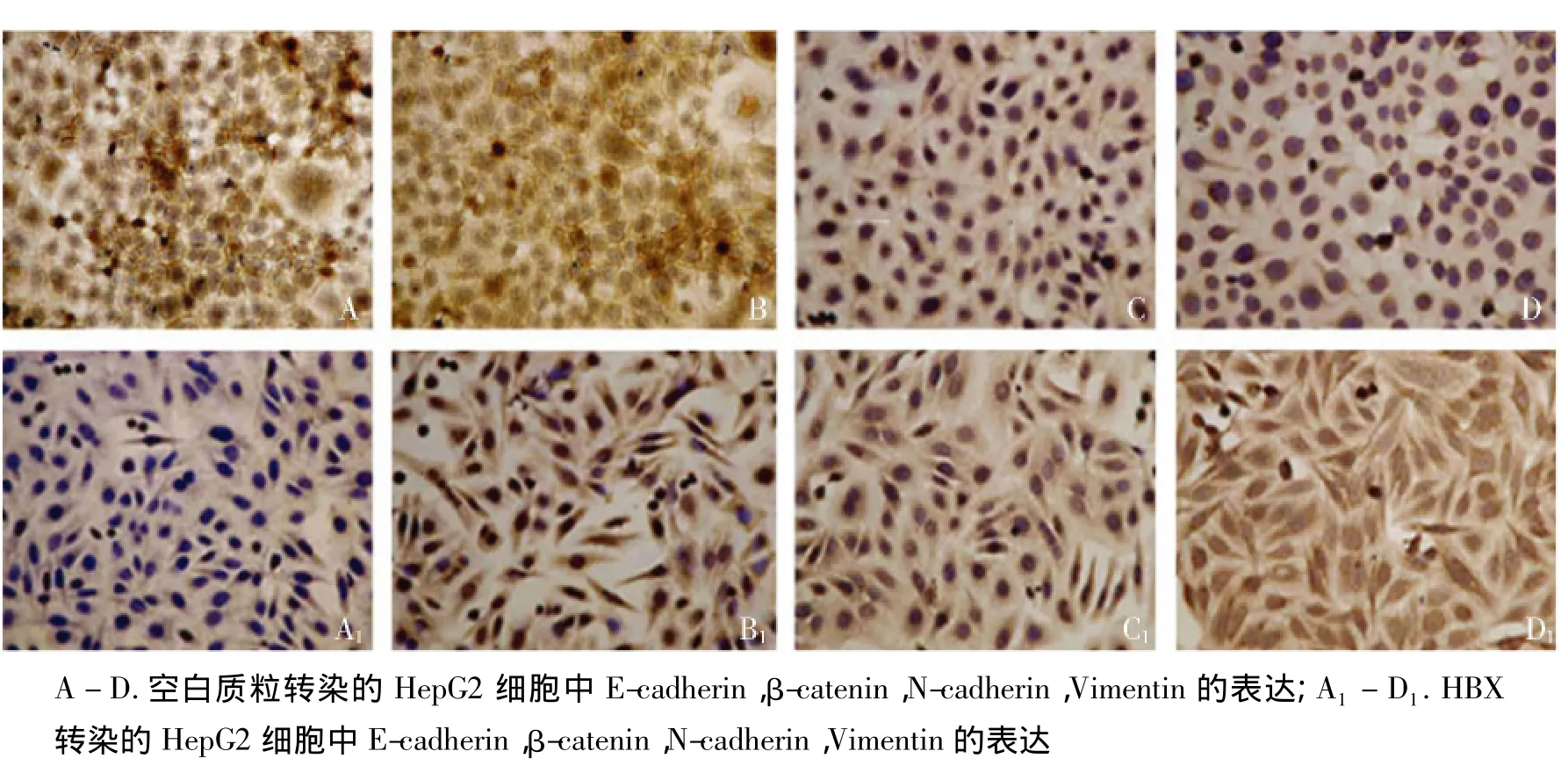
圖3 免疫組化檢測HBx對HepG2細胞EMT標志物的表達Figure 3 EMT markers expression in HepG2 cells by immunohistochemistry
2.3 c-Src抑制劑阻斷HBx介導的HepG2細胞形態改變
為進一步探索c-Src活性對HBx介導的EMT改變的影響,我們應用c-Src特異性抑制劑PP2處理HBx轉染的HepG2細胞,應用PP3作為PP2的陰性對照。PP2處理的HBx轉染的HepG2細胞失去了梭形形態,細胞間的連接緊密,而PP3處理的HBx轉染的HepG2細胞未見明顯的細胞形態改變(見圖4),c-Src特異性抑制劑逆轉了HBx介導的HepG2細胞的EMT改變。

圖4 PP2抑制HBx介導的HepG2細胞形態改變Figure 4 PP2 inhibits the HBx-induced morphologic changes of HepG2 cells
2.4 c-Src抑制劑阻斷 HBx介導的 HepG2細胞EMT標志分子表達改變
基于以上的形態觀察,我們更進一步探索了PP2對HBx轉染的HepG2細胞中EMT標志物的表達情況。與之前的觀察結果類似,在HBx轉染的HepG2細胞中,PP2恢復了上皮細胞標志E-cadherin的表達,下調了間質細胞標志N-cadherin和Vimentin的表達,而PP3并未對E-cadherin,N-cadherin和Vimentin的表達產生影響(見圖5)。免疫組化的觀察也支持PP2對HBx介導的EMT信號通路的阻斷作用,同時PP2的應用顯著破壞了β-catenin的胞核分布(見圖6)。

圖5 PP2抑制HBx轉染的HepG2細胞EMTFigure 5 PP2 inhibits HBx-induced EMT in HepG2 cells
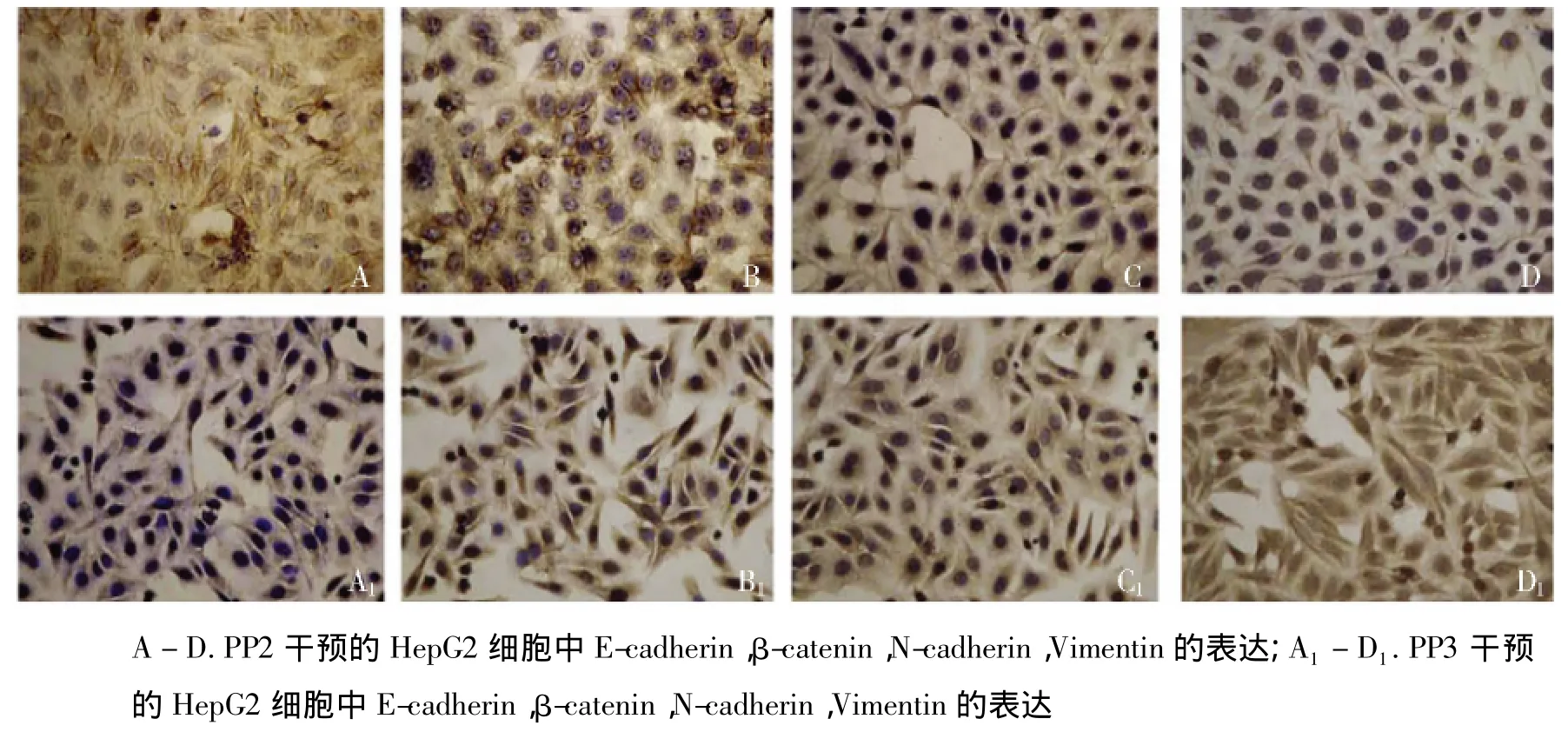
圖6 免疫組化檢測PP2和PP3對HBx轉染的HepG2細胞EMT標志物表達Figure 6 EMT markers expression in HepG2 cells treated by PP2 or PP3 by immunohistochemistry
3 討論
在眾多惡性腫瘤促侵襲轉移的機制當中,EMT是近期的研究熱點。EMT主要的功能特點是使惡性腫瘤細胞失去上皮細胞功能,而展現間質細胞的功能,功能改變導致惡性腫瘤細胞的細胞間連接趨于松散,細胞骨架進行重組,細胞形態呈現出成纖維細胞樣改變,這一系列改變導致惡性腫瘤細胞侵襲轉移能力增強[3,4]。在分子水平上,EMT的主要特征是上皮細胞標志物—E-cadherin等表達下調,而間質細胞標志物—N-cadherin,Vimentin等表達上調[3,4]。
在我們的研究當中,肝細胞癌HepG2細胞展現出了典型的上皮細胞形態特點,穩定轉染HBx基因的HepG2細胞形態發生了明顯的改變,呈現出了梭形改變,類似成纖維細胞,細胞與細胞之間的連接松散,這些觀察表明外源性HBx基因轉染介導HepG2細胞發生EMT形態改變。進一步Western blot和免疫組化結果表明:HBx基因轉染導致上皮細胞標志E-cadherin表達下調,而間質細胞標志N-cadherin和Vimentin表達上調,這些結果進一步支持了HBx具有介導HepG2細胞EMT改變的重要作用。EMT標志物表達的改變也被稱為“cadherin開關效應”,這種cadherin開關效應最初被Tomita等在前列腺癌中觀察到,E-cadherin下調是EMT的標志改變[5]。
E-cadherin在維持上皮細胞表型和同源細胞間的黏附功能至關重要,在多種惡性腫瘤細胞系中,E-cadherin表達下調有效地上調了細胞侵襲轉移能力[6],另一方面,E-cadherin表達下調間接促進了其他類型 cadherin表達,如 N-cadherin,N-cadherin表達上調增加了細胞的遷移能力,促進了細胞與內皮細胞、細胞與細胞外基質間的新黏附結構形成,從E-cadherin到N-cadherin改變有效促進了惡性腫瘤細胞的侵襲轉移和血管侵犯[7,8]。
我們的免疫組化結果表明,HBx基因轉染的HepG2細胞呈現了EMT改變,同時,β-catenin在其胞質和胞核中均有較高水平的表達,對照細胞中,βcatenin主要分布在近細胞膜處。這些結果與之前的報道類似[9],說明β-catenin在EMT中發揮了轉錄因子作用。在Wnt信號通路中,已經證實伴隨E-cadherin表達下調,β-catenin發生細胞質到細胞核的定位改變,在細胞核中,β-catenin發揮了轉錄因子的作用,上調了N-cadherin和vimentin表達,導致EMT 的發生[9,10]。
c-Src是細胞內具有重要功能的一個非受體酪氨酸激酶,c-Src活性升高能夠顯著增強多種惡性腫瘤細胞的侵襲轉移,其促侵襲轉移的分子機制十分復雜[11]。c-Src能夠通過活化 Ras/Raf-1/MAPK信號通路減弱細胞間的黏附作用,促進惡性腫瘤細胞的侵襲轉移[12],另一方面,c-Src可以在近細胞膜處活化小分子GTP酶,導致細胞骨架重組,增強了惡性腫瘤細胞的遷移能力[13]。近期的研究結果表明,c-Src可以磷酸化E-cadherin,導致 E-cadherin泛素化降解,介導EMT改變發生[14]。我們的研究結果進一步支持了c-Src對肝癌HepG2細胞EMT具有重要的促進作用。
本研究結果表明,HBx基因轉染顯著提高了HepG2細胞的c-Src活性,在應用c-Src特異性抑制劑PP2作用于轉染HBx基因的HepG2細胞后,EMT改變被有效的逆轉,細胞形態恢復上皮樣改變,E-cadherin表達恢復,N-cadherin和Vimentin表達下調,β-catenin細胞核表達顯著減少,這些結果表明c-Src在惡性腫瘤細胞EMT中發揮了重要的作用。由于c-Src相關信號分子十分復雜,由此推測不同信號分子間可能存在cross-talk,最終介導惡性腫瘤細胞的EMT發生。
4 結論
HBx基因的轉染導致肝細胞癌HepG2細胞發生EMT改變,c-Src分子在HBx介導的EMT改變中發揮了至關重要的作用,更進一步的分子機制的探索對于揭示更深層的EMT改變是必要的,EMT分子機制的明確將為控制惡性腫瘤細胞的侵襲轉移提供新的思考。
[1]Kubo S,Takemura S,Tanaka S,et al.Management of hepatitis B virus infection during treatment for hepatitis B virus-related hepatocellular carcinoma[J].World J Gastroenterol,2015,21(27):8249-8255.
[2]Chan CF,Yau TO,Jin DY,et al.Hepatitis B virus X protein(HBx)is responsible for resistance to targeted therapies in hepatocellular carcinoma:ex vivo culture evidence[J].Clin Cancer Res,2015,21(5):8140-8149.
[3]Zheng H,Kang Y.Multilayer control of the EMT master regulators[J].Oncogene,2014,33(14):1755-1763.
[4]Puisieux A,Brabletz T,Caramel J.Oncogenic roles of EMT-inducing transcription factors[J].Nat Cell Biol,2014,16(8):488-494.
[5]van Roy F.Beyond E-cadherin:roles of other cadherin superfamily members in cancer[J].Nat Rev Cancer,2014,14(2):121-134.
[6]Lecuit T,Yap AS.E-cadherin junctions as active mechanical integrators in tissue dynamics[J].Nat Cell Biol,2015,17(5):533-539.
[7]Radice GL.N-cadherin-mediated adhesion and signaling from development to disease:lessons from mice[J].Prog Mol Biol Transl Sci,2013,116:263-289.
[8]Wang F,Li XK,Xu HY,et al.N-cadherin participated in invasion and metastasis of human esophageal squamous cell carcinoma via taking part in the formation of vasculogenic mimicry[J].Med Oncol,2015,32(2):480.
[9]Liu J,Lian Z,Han S,et al.Downregulation of E-cadherin by hepatitis B virus X antigen in hepatocellular carcinoma[J].Oncogene,2006,25(7)1008-1017.
[10]Fernandez-Sanchez ME,Barbier S,Whitehead J,et al.Mechanical induction of the tumorigenic beta-catenin pathway by tumour growth pressure[J].Nature,2015,523(7558):92-95.
[11]Bordoli MR,Yum J,Breitkopf SB,et al.A secreted tyrosine kinase acts in the extracellular environment[J].Cell,2014,158(5):1033-1044.
[12]McLachlan RW,Kraemer A,Helwani FM,et al.E-cadherin adhesion activates c-Src signaling at cell-cell contacts[J].Mol Biol Cell,2007,18(8):3214-3223.
[13]Frame MC.V-SRC informs integrin signaling[J].Nat Rev Mol Cell Biol,2013,14(9):548.
[14]Wilson C,Nicholes K,Bustos D,et al.Overcoming EMT-associated resistance to anti-cancer drugs via Src/FAK pathway inhibition[J].Oncotarget,2015,5(17):7328-7341.
