產木聚糖酶海洋微生物的篩選與誘變育種
2015-01-03 05:52:56黃小云林娟林小洪王國增葉秀云
福州大學學報(自然科學版) 2015年5期
黃小云,林娟,林小洪,王國增,葉秀云
(福州大學生物科學與工程學院,福建省海洋酶工程重點實驗室,福建福州 350116)
0 引言
木聚糖是植物細胞壁半纖維素的主要成分,包括由β-1,4糖苷鍵形成主鏈結構和不同的側鏈取代基,常見的取代基包括4-O-甲基葡萄糖醛酸(4-O-Met-Hylglucuronic)、α-阿拉伯糖(α-arabinofuranose)及O-乙酰基(O-Acetyl group)等[1].木聚糖的完全降解需要多種水解酶的協同作用[2].木聚糖酶(EC 3.2.1.8)通過隨機切割木聚糖主鏈骨架上的β-1,4糖苷鍵產生不同長度的低聚木糖[3],是木聚糖降解酶系中最為關鍵的酶.
木聚糖酶的來源較廣泛,包括陸地植物組織、細菌、真菌和原生動物等[3],但大部分來源于陸地,對于海洋微生物的研究還比較少.海洋微生物所產的酶具有突出的特點和優勢:具有顯著的耐壓、耐堿、耐鹽、耐冷和多物種等特性,這些優越的性質使得海洋酶資源在工業應用和基礎研究等方面具有價值,從而賦予海洋微生物酶獨特的應用前景[4].木聚糖酶可作為食品添加劑用于改善焙烤食品的風味[5],在飼料中添加木聚糖酶可提高動物的生產性能[6],用木聚糖酶對紙漿的預處理會大大減少化學漂白劑的用量[7].由于其在各領域的重要作用,選育高產木聚糖酶的菌株成為目前木聚糖降解菌選育研究的熱點.
1 材料與方法
1.1 樣品
樣品分別來自北極泥樣,福州市平潭海域的水樣和泥樣,寧德市三都澳海域的泥樣和海藻,以及鮑魚內臟等.
1.2 培養基
1)初篩培養基:自制甘蔗渣木聚糖15 g,NH4NO35 g,K2HPO42 g,MgSO4·7H2O 0.5 g,瓊脂20 g,人工海水[8]1 000 mL,pH 7.0;
2)種子培養基:3 g牛肉膏,10 g蛋白胨,5 g NaCl,人工海水1 000 mL,pH 7.0;3)基礎產酶培養基:20 g麩皮,10 g酵母膏,人工海水1 000 mL,pH 7.0.
1.3 主要儀器設備
超凈工作臺(SW-CJ-2FD型,上海博迅實業有限公司醫療設備廠);生化培養箱(MJX-250S型,寧波海曙賽福實驗儀器廠);恒溫氣浴搖床(DHZ-DA型,太倉市實驗設備廠);雙光束分光光度計(U-2910型,日立有限公司);pH計(PHS-3C型,上海精密科學儀器有限公司).
1.4 實驗方法
1.4.1 自制木聚糖
取粉碎過的甘蔗渣加入60 g·L-1NaOH溶液中,固液比為1∶15,100℃條件下浸泡2 h.過濾后的濾液用36% ~38%(質量分數)HCl調pH值至7.0,按濾液 ∶乙醇為1∶0.8的比例(體積分數),加入濃度為95%(體積分數)的工業酒精,所得棕褐色或棕黃色沉淀即為木聚糖.4 000 r·min-1下離心10 min得沉淀,置于75℃條件下烘干后研磨至粉末狀即可使用[9].
1.4.2 樣品處理及木聚糖酶產生菌的篩選
稱取1.1所示的幾份樣品(海藻和鮑魚內臟先用無菌攪拌器研磨)于無菌生理鹽水中進行梯度稀釋.將10-1、10-6、10-7和10-8這4個梯度的樣品懸液分別涂布于木聚糖酶篩選平板,將純化后的單菌落點種于初篩培養基上,測定透明圈直徑(H)和菌落直徑(C)的大小,計算HC值.將初篩獲得的菌株在平板上活化后,轉接至種子培養基中,30℃、200 r·min-1培養12 h,取種子液以10%(體積分數)的接種量接入基礎產酶培養基中,30℃、200 r·min-1培養72 h,發酵液離心后取上清進行酶活測定.
1.4.3 木聚糖酶活力測定
1.4.3.1 木糖標準曲線繪制
配制1.5 μmol·mL-1木糖標準溶液,分別量取 0、0.20、0.25、0.30、0.35、0.40、0.45、0.50、0.60 mL木糖標準溶液,用pH6.0的檸檬酸-磷酸氫二鈉緩沖液補至1 mL,再加入1 mL DNS,沸水浴10 min,冷卻至室溫后在540 nm測定OD值,繪制木糖標準曲線.
1.4.3.2 木聚糖酶活力測定
取0.9 mL含1%(質量分數)木聚糖的底物,加入0.1 mL適當稀釋的酶液,55℃準確反應10 min后加入1 mL DNS溶液終止反應,煮沸10 min顯色,冷卻至室溫后于540 nm測定OD值.一個酶活力單位(U)定義為每min釋放1μmol還原糖所需的酶量.
1.4.4 形態學觀察及16S rDNA序列分析
依據《伯杰細菌鑒定手冊》進行菌落特征和細胞形態的鑒定[10].菌株的分子鑒定以16SrDNA作為指標.以基因組為模板,選用細菌通用引物27F(AGAGTTTGATCMTGGCTCAG)和1492R(TACGGYTACCTTGTTACGACTT)進行16SrDNA擴增,擴增條件為:94℃ 5 min,94℃ 30 s,48℃ 30 s,72℃ 2 min,30個循環;72℃ 10 min.
1.4.5 紫外及微波誘變
吸取5 mL菌懸液(濃度為(107~108)cfu·mL-1)于裝有轉子的無菌培養皿內,紫外誘變是將菌懸液經紫外燈(15 W,照射距離30 cm)照射30、60、90、120、150、180、210 s,微波誘變則是將菌懸液在微波爐最小功率下分別輻照0、1、2、3、4、5、6、7 min.對菌懸液進行梯度稀釋,分別取10-5、10-6、10-7三個稀釋度涂布于篩選平板,以未誘變的菌懸液作為對照.將上述所有平板(紫外誘變的平板需避光培養)在30℃恒溫箱中培養48 h后取出計算平板上的菌落數,繪制致死率曲線.致死率計算公式如下:
致死率(%)=(1-誘變后的菌落數/對照組的菌落數)×100%.
1.4.6 遺傳穩定性實驗
將篩選到的木聚糖酶活力提高較多的正突變菌株,連續傳代5代以上,并通過搖瓶發酵測定其產酶能力,選擇產酶能力較高且穩定遺傳的木聚糖酶正突變株進行后續的實驗.
2 結果與討論
2.1 平板初篩結果
從采集的各樣品中分離到能降解木聚糖底物產生透明圈且菌落形態和細胞形態有差異的菌株60株(見表1),其中菌株編號為B459~B663的來自福州平潭海域的水樣和泥樣;菌株編號為Y的來自鮑魚內臟;菌株編號為Z的來自寧德三都澳海域的海藻;菌株編號為M和N的來自寧德三都澳海域的泥樣;菌株編號為MOR、BOB和BBO的是來自北極的泥樣.
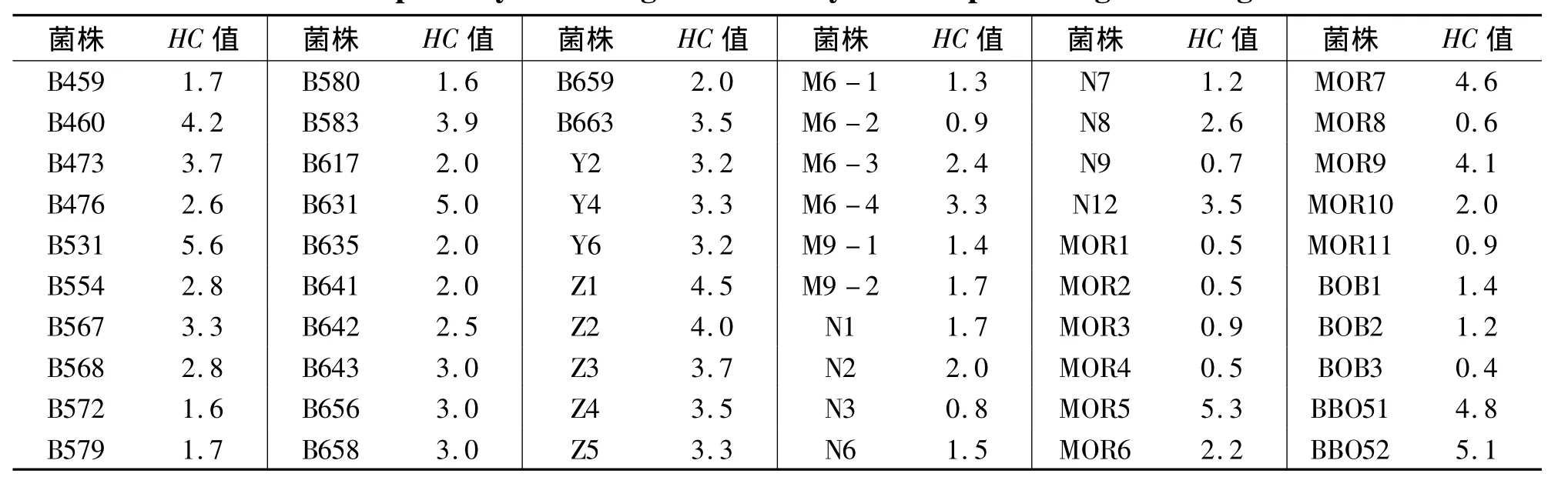
表1 產木聚糖酶微生物的初篩結果Tab.1 The primary screening results of xylanase-producing microorganisms
2.2 發酵復篩結果
對初篩得到的60株菌進行搖瓶發酵測定木聚糖酶活力,結果顯示,有22株菌不具有產木聚糖酶的能力,38株菌能產木聚糖酶(見表2).
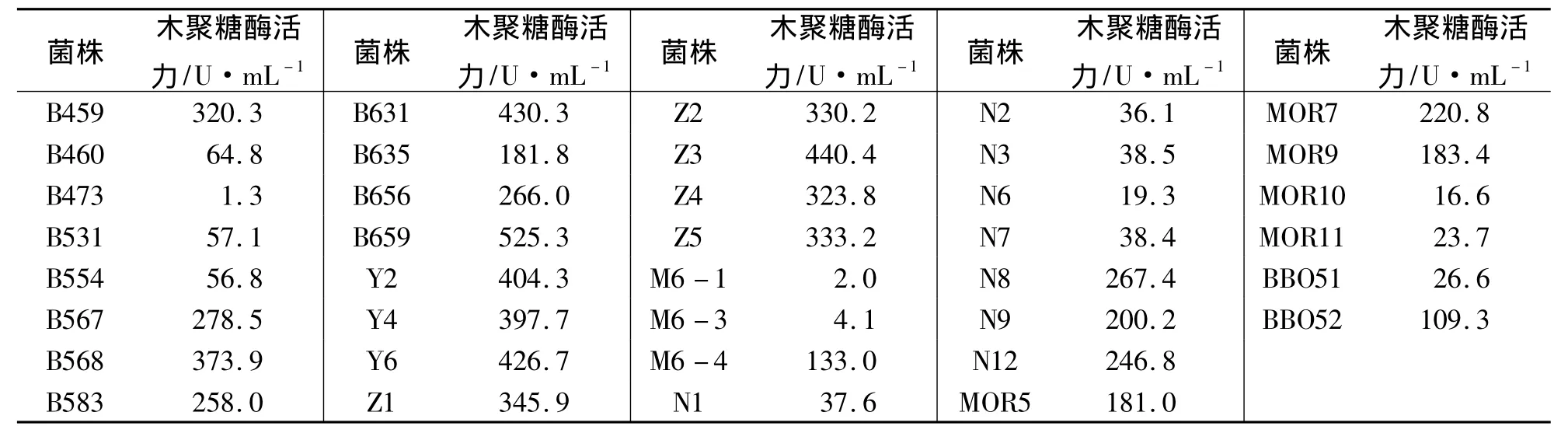
表2 產木聚糖酶微生物的復篩結果Tab.2 The secondary screening results of xylanase-producing microorganisms
產酶能力在0~100 U·mL-1的有14株菌,100~200 U·mL-1的有5株菌;200~300 U·mL-1的有7株菌;大于300 U·mL-1的有12株菌,其中B659菌株的木聚糖酶活力最高,達到525.3 U·mL-1.
2.3 木聚糖酶的酶學性質初步研究
選取酶活力相對較高的6株菌(B250、B631、B659、Y6、Z3和MOR7),對其所產的木聚糖酶進行酶學性質的初步研究,包括最適反應pH值和最適反應溫度.實驗結果見圖1.由圖1可知,這6株菌所產的木聚糖酶的最適反應pH值均為6.0,最適反應溫度也很相近,分別是55℃和60℃;而且酶的pH值和溫度作用范圍也都相差不大.因此,選擇產酶能力最高的B659菌株作為下一步研究的實驗菌株.
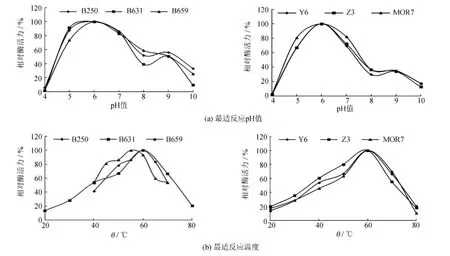
圖1 反應pH值和溫度對木聚糖酶活力的影響Fig.1 Effect of pH and temperature on xylanase activity
2.4 菌株B659的形態鑒定及16S r DNA鑒定結果
B659菌株在篩選培養基平板上30℃培養48 h,菌落呈乳白色,小而突起,表面濕潤皺褶,邊緣圓整(圖2(a)).顯微鏡下觀察,細胞呈短桿狀,隨機分散不成鏈狀,細胞中有橢圓形芽孢,位置中生(圖2(b)).經形態觀察初步鑒定B659菌株為芽胞桿菌屬.
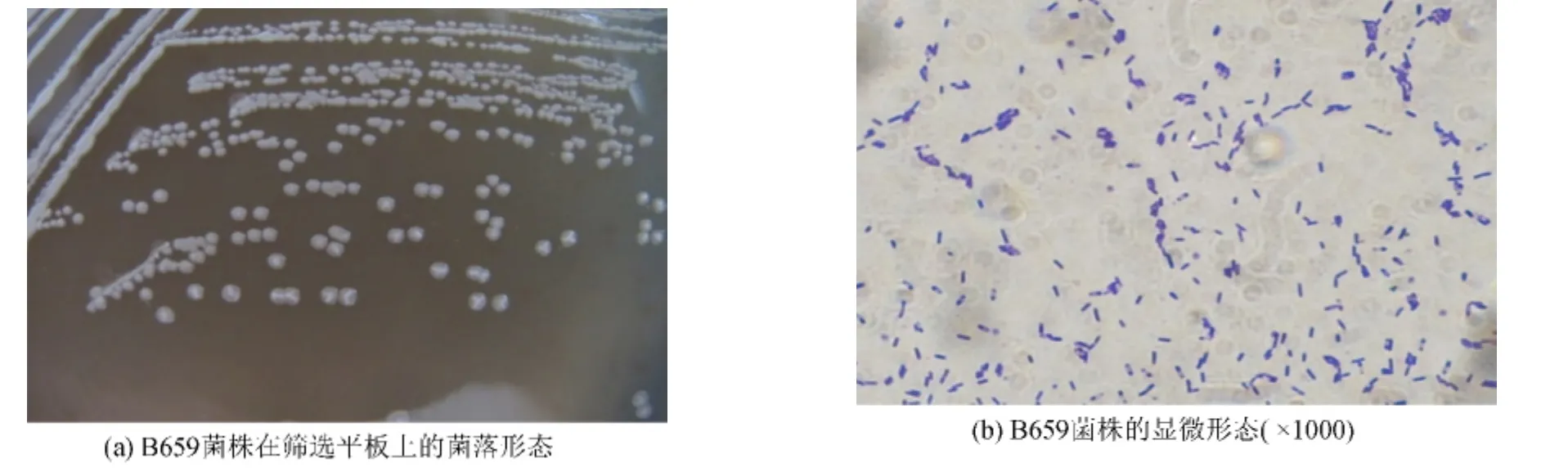
圖2 B659菌株的形態觀察Fig.2 The morphology of strain B659
基于BLAST分析,菌株B659的部分16S rDNA(1 510 bp)與Bacillus stratosphericus strain MCCC 1A08157(序列號:JX680139)的16SrDNA一致性為99%,構建其系統發育樹(見圖3),發現與同溫層芽胞桿菌(Bacillus stratosphericus)、嗜氣芽胞桿菌(Bacillus aerophilus)、短小芽胞桿菌(Bacillus pumilus)和沙福芽胞桿菌(Bacillus safensis)的親緣關系最近,因此鑒定B659菌株屬于Bacillus屬.

圖3 B659菌株的系統發育樹Fig.3 The phylogenetic tree of strain B659
2.5 菌株B659的誘變育種
2.5.1 紫外誘變
2.5.1.1 誘變劑量的選擇
繪制B659菌株的生長曲線.由圖4可知,菌株B659在10~14 h時處于對數生長期的中期,此時細胞數量多且生長速率恒定、細胞各成分平衡增長、整個菌群的生理特性比較一致[11],因此選擇培養12 h的B659菌株制備菌懸液進行誘變.
不同照射劑量所對應的致死率如圖5所示.當照射時間為150 s時,致死率達到了79.4%;照射時間為180 s時,致死率達到94.1%;210 s時,致死率達到了100%.致死率在70% ~80%時有利于正突變的產生[12],故實驗選用150 s作為照射劑量對菌株進行紫外誘變處理.
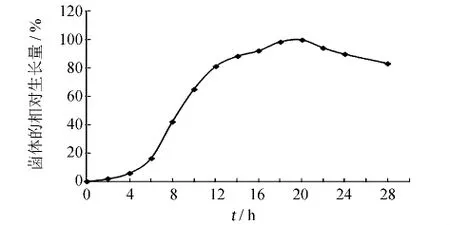
圖4 B659菌株的生長曲線Fig.4 The growth curve of strain B659
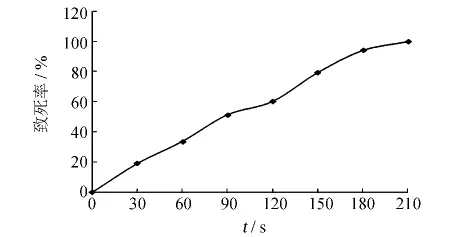
圖5 B659菌株的紫外誘變曲線Fig.5 The UV mutation curves of strain B659
2.5.1.2 正突變菌株的篩選
出發菌株B659在150 s的照射劑量下,經過2輪紫外誘變在初篩平板上共得到了236株菌,挑選118株菌通過搖瓶發酵測定其酶活力.結果表明,118株突變菌株中有5株菌的產木聚糖酶能力高于出發菌株,詳見表3.
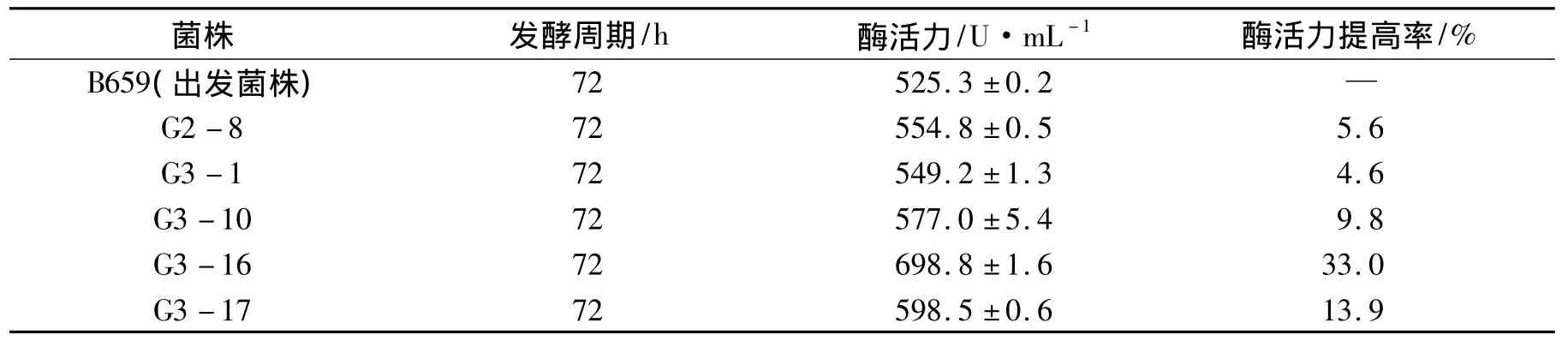
表3 出發菌株與突變菌株的木聚糖酶活力比較Tab.3 The comparison of xylanase activity between the original and the mutant strain
其中G3-16菌株的產酶能力最高,可達到698.8 U·mL-1,比出發菌株B659提高33.0%;其次是G3-17和G3-10菌株,其產酶能力分別是598.5 U·mL-1和577.0 U·mL-1,較出發菌株分別提高了13.9%和 9.8%.
2.5.1.3 突變菌株的遺傳穩定性
將酶活提高相對較多的突變菌株G3-10、G3-16和G3-17進行5次傳代培養,取第1代、第3代和第5代的菌株于產酶培養基中進行搖瓶發酵,72 h后取樣測定各菌株的酶活力,結果如表4所示.由表可知,G3-16菌株雖然誘變后酶活力提高最多,但很不穩定,到第3代時酶活力大幅度下降,甚至低于出發菌株;G3-10菌株和G3-17菌株具有較好的遺傳穩定性,而G3-17菌株的產酶能力高于G3-10菌株,因此最后確定G3-17菌株作為下一輪誘變的出發菌株.
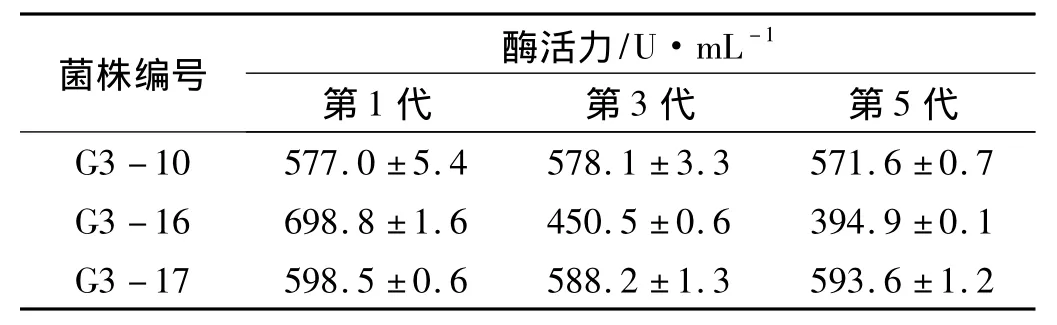
表4 突變菌株的遺傳穩定性Tab.4 Genetic stability of mutant strain
2.5.2 微波誘變
2.5.2.1 誘變劑量的選擇
本輪誘變以紫外誘變所得突變菌株G3-17作為出發菌株,對其進行微波誘變,不同劑量所對應的致死率曲線如圖6所示.由圖6可知,出發菌株G3-17的致死率隨著誘變時間的延長而增加.在輻射時間為4 min時,致死率達到83.2%;輻射時間為5 min時,致死率達到90.1%;輻射6 min以后,致死率達到100%.本實驗選用4 min作為輻射劑量對G3-17菌株進行微波誘變處理.
2.5.2.2 出發菌株與正突變菌株的酶活力比較
出發菌株G3-17在4 min的輻射劑量下,經過2輪的微波輻射后在初篩平板上共得到了183株菌,挑選92株突變菌株全部進行搖瓶發酵測定其產酶能力.結果表明,92株突變菌株中有6株菌的產木聚糖酶能力高于出發菌株G3-17,見表5.其中W1-40菌株的產酶能力最高,可達到662.8 U·mL-1,比出發菌株G3-17提高了11.6%;W1-28和 W1-41次之,分別達到了654.3 U·mL-1和646.8 U·mL-1,其余3株菌酶活提高幅度較小.
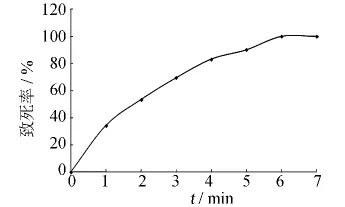
圖6 微波誘變的致死率曲線Fig.6 The lethality rate of microwave mutagenesis
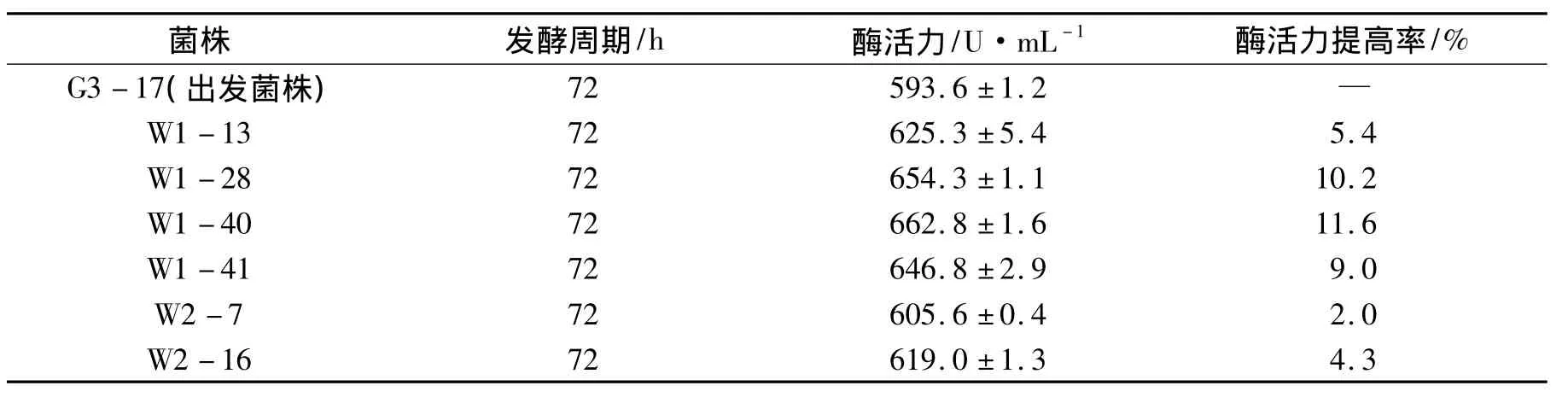
表5 出發菌株與突變菌株的木聚糖酶活力比較Tab.5 The comparison of xylanase activity between the original and the mutant strain
2.5.2.3 突變菌株遺傳穩定性分析
將酶活提高相對較多的突變菌株W1-13、W1-28、W1-40和W1-41進行5次傳代培養,結果如表6所示.
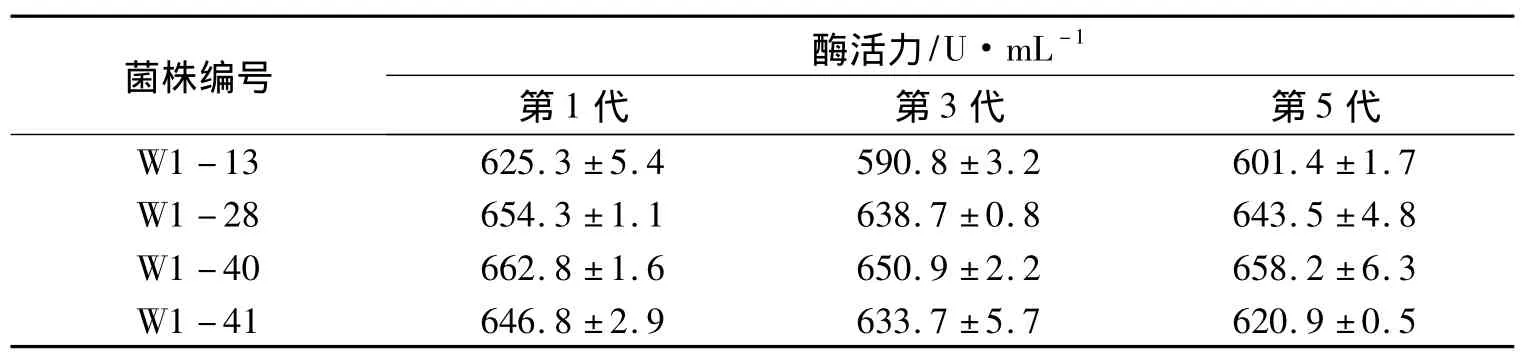
表6 遺傳穩定性的測定Tab.6 Genetic stability of mutant strain
這4株菌中W1-13遺傳不穩定,其它3株都具有較好的遺傳穩定性.其中W1-40的產酶能力最高,達到658.2 U·mL-1,且遺傳穩定性良好,因此確定W1-40菌株作為后續實驗的研究對象.
2.5.3 原始菌株和突變菌株的產酶曲線比較
把原始菌株B659和突變菌株W1-40分別接入產酶培養基進行發酵試驗,每隔12 h取樣測定酶活,比較2株菌的產酶變化,繪制產酶進程曲線.由圖7可知,B659菌株和W1-40菌株的酶活都是隨著發酵時間的延長而上升,在12~36 h時上升最快,72 h時產酶量達到最高(B659菌株和W1-40菌株分別為517.9和645.2 U·mL-1);72 h之后可能由于木聚糖酶失活速率高于產酶速率,酶活力開始下降.
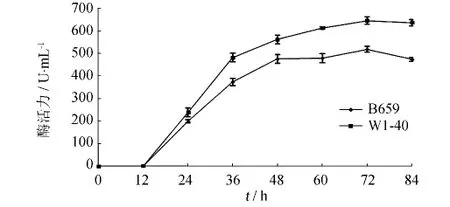
圖7 B659菌株和W1-40菌株的產酶曲線Fig.7 Effect of incubation time on xylanase production by strain B659 and strain W1-40
3 結論
采用透明圈法從多個海洋來源的樣品中共篩到60株有透明圈的菌株,從中復篩得到38株具有產木聚糖酶能力的菌株,其中經鑒定為芽孢桿菌屬的菌株B659產酶能力最高(酶活力為525.3 U·mL-1).對菌株B659進行紫外誘變得到遺傳穩定且酶活提高13.9%的突變菌株G3-17,對突變菌株G3-17進行微波誘變得到遺傳穩定且酶活較菌株G3-17提高11.6%的突變菌株W1-40.發酵72 h時,突變菌株W1 -40的酶活力(645.2 U·mL-1)比原始菌株 B659(酶活為517.9 U·mL-1)提高24.6%.
[1]Biely P.Microbial xylanolytic systems[J].Trends in Biotechnology,1985,3(11):286 -290.
[2]Beg Q K,Kapoor M,Mahajan L,et al.Microbial xylanases and their industrial applications:a review[J].Applied Microbiology and Biotechnology,2001,56(3/4):326-338.
[3]Collins T,Gerday C,Feller G.Xylanases,xylanase families and extremophilic xylanases[J].FEMS Microbiology Reviews,2005,29(1):3-23.
[4]侯運華.適冷海洋青霉纖維素酶和半纖維素酶的分子生物學研究[D].濟南:山東大學,2006.
[5]Camacho N A,Aguilar O G.Production,purification,and characterization of a low -molecular-mass xylanase from Aspergillus sp.and its application in baking[J].Applied Biochemistry and Biotechnology,2003,104(3):159 -172.
[6]Wong K K,Tan L U,Saddler JN.Multiplicity of beta-1,4-xylanase in microorganisms:functions and applications[J].Microbiological Reviews,1988,52(3):305 -317.
[7]Bajpai P.Application of enzymes in the pulp and paper industry[J].Biotechnology Progress,1999,15(2):147 -157.
[8]Takai K,Sugai A,Itoh T,et al.Palaeococcus ferrophilus gen.nov.,sp.nov.,a barophilic,hyperthermophilic archaeon from a deep - sea hydrothermal vent chimney[J].International Journal of Systematic and Evolutionary Microbiology,2000,50(2):489-500.
[9]李秀婷,孫寶國,宋煥祿,等.玉米芯水不溶性木聚糖的堿法提取及酶解分析[J].中國食品學報,2010,5(10):171-176.
[10]R.E.布坎南,N.E.吉本斯.伯杰細菌鑒定手冊[M].北京:科學出版社,1984:729-758.
[11]周德慶.微生物學教程[M].北京:高等教育出版社,2002:152-159.
[12]雷肇祖,錢志良,章健.工業菌種改良述評[J].工業微生物,2004,34(1):39-51.
