修文縣喀斯特示范區退化森林群落數量特征研究
2015-01-27 09:04:01蔡品迪
吉林農業·下半月 2014年9期
關鍵詞:結構

摘要:采用“空間代替時間”的方法對修文縣喀斯特示范區植被群落進行調查研究,結果表明:雖然示范區植物種類相對豐富,但是物種組成以矮、小高位芽植物為主,并且萌生植株比例較高,株數密度過大,群落演替到頂級群落難度大;物種多樣性指數在不同層次的變化存在著一定差異;隨著演替進展,總生物量呈上升趨勢,但各層次生物量分布較不合理,在喬木林階段喬木層生物量所占比例較低,而灌木層卻依然占有很高的比例,群落總體處于一個較低的演替階段,需要進行結構調整,優化物種組成結構,才能促進植被恢復。
關鍵詞:植被退化;組成;結構;多樣性;生物量
中圖分類號:S718.54 文獻標識碼: A 文章編號: 1674-0432(2014)-18-22-4
中國西南喀斯特地區,因地質環境脆弱性大、敏感度高,且面臨人口超載和經濟社會落后的雙重壓力,致使生態環境嚴重退化[1],植被退化現象日益加劇,石漠化土地面積不斷增大,已經成為該地區最主要的生態地質環境問題。本研究主要對不同演替階段群落主要特征參數進行統計分析,掌握群落演替過程中組成結構動態變化趨勢,為喀斯特地區開展結構調整技術研究提供理論依據。
1 研究區概況
研究區域位于貴州省貴陽市修文縣龍場鎮城邊,烏江支流的貓跳河右岸,海拔1100~1500米,屬亞熱帶季風氣候區,年均溫13.6℃,≥10℃積溫4097.4℃,年降雨量1235毫米,分布不均,易出現干旱現象。試驗示范區面積共1500畝,包括強、中、輕度三類石漠化等級,其中強度石漠化面積為260畝,中度石漠化面積為515畝,輕度石漠化面積為725畝。母巖以白云質灰巖為主。土壤為發育在白云質灰巖上的黃色石灰土和黑色石灰土,pH4~7.4,土層淺薄,石礫含量高。現存植被為各種次生喬林、灌木林、藤刺灌叢、草坡以及石漠化荒地,蓋度15%。主要植物種類有白櫟(Quercus fabri Hance)、川榛(Corylus heterophylla Fisch. ex Trautv. var. sutchuenensis Franch)、茅栗(Castanea seguinii Dode)、火棘(Pyracantha fortuneana(Maxim.)Li)、光皮樺(Betula luminifera H. Winkl.)、紅葉木姜子(Litsea rubescens Lec.)、響葉楊(Populus adenopoda Maxim.)、云南鼠刺(Itea yunnanensis Franch.)、鹽膚木(Rhus chinensis Mill.)、麻櫟(Quercus acutissima Carr.)等。當地村民經濟條件較差,以農業為主要經濟來源,副業以挖鋁土礦為主。
2 研究方法
2.1 群落樣地調查
采用“以空間序列代替時間序列”的方法[2],在草坡階段、灌草階段、灌叢灌木階段、灌木林階段、喬灌階段、喬木林階段六個演替階段設置樣地,共計21個樣地,進行常規群落學調查,樣地面積根據群落最小面積確定,其大小分別為4×5米、10×10米、10×10米、10×10米、8×20米、20×20米。喬林群落樣地中分為10個5×8米喬木層樣方,且在每個喬木層樣方內設置1個2×2米灌木層樣方和1個1×1米草本層樣方。調查記錄每個樣方內喬、灌木及幼樹的種類、數量、胸徑、地徑、高度、冠幅、蓋度,并判定每株樹木是實生或萌生起源;草本植物調查其種類、多度、高度、蓋度。
群落生物量按喬木層、灌木層、草本層分別調查。其中喬木層生物量采用朱守謙等[3]的研究公式進行計算,灌木層采用樣本法,根據調查結果,按樹種、地徑選取樣本,用收獲法測定樣木樹干、枝、葉的鮮重,共獲取樣本107株。在每塊樣地內設置2個1×1米,采用刈割法獲取草本層地上部分鮮重。取樣品在80℃恒溫烘至恒重,計算含水率并換算成干重。
2.2 數據處理
不同恢復階段群落物種多樣性主要采用以下指標測度:
(1)Gleason豐富度指數:
(2)Simpson多樣性指數:
(3)Shannon-wiener多樣性指數: ;
(4)Pielou均勻度指數:
(5)生態優勢度指數:
式中:S為物種數,A為樣方面積,Pi為種i相對重要值,N為總個體數,Ni為種i個體數。
灌木層采用
喬木層生物量的計算直接用公式(1) [3]。
植被生活型采用瑙基耶爾(Raunkiaer)[4]的生活型系統進行劃分。
3 結果與分析
3.1 群落組成結構分析
植物群落結構的復雜程度在一定程度取決于構成群落的植物種類的多少,一般而言植物種類多,群落結構就復雜。調查結果顯示:本研究區域內不同演替階段植物共分布有67個科,122個屬,共計152個種,植物種類多,但少數物種優勢明顯,如白櫟、光皮樺等,植被類型較為單一。
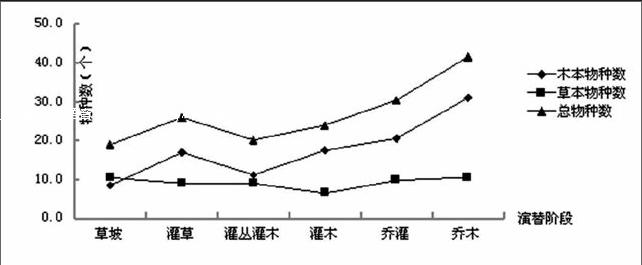
3.1.1群落物種數變化 從圖1可以看出隨著植被恢復過程,群落的總物種數總體呈現上升趨勢,在灌叢灌木階段由于遭受人為破壞,萌生能力較強的物種,如白櫟、小果南燭等植被大量萌生,獲取有限的空間資源,抑制了其他植被生長,物種數略微減少,當進入演替中期,由于萌生植株出現生長衰退,而適應能力更強的實生物種開始逐漸占領空間資源,促使一些新的物種出現,物種數又開始上升;木本物種數與總物種數的變化趨勢相同,兩者呈極顯著相關;而草本物種數從草坡階段到灌木階段種數不斷減少,當進入喬灌后種數又逐漸增多,呈現出先下降后升高的趨勢,可能是由于從草坡階段演替到灌木階段過程中,植物種間競爭激烈,木本植物的逐漸增多,不斷地吸取養分、光照等資源,抑制了草本植物的生長,導致一些競爭力較弱的草本物種死亡并消失,當進入喬灌階段以后,群落逐漸形成了喬、灌、草的分層結構,生態系統也逐漸穩定,環境條件得到了較好的改善,這就給林下草本植物生長提供了良好的生長環境,促使一些耐陰性草本植物的出現,故草本種類又呈現上升趨勢。
圖1 不同演替階段群落物種組成變化
3.1.2 群落生活型組成 生活型(Life form)是植物對生態環境適應的外部表現形式,同一生活型的植物,不但形態相似,而且其生態適應特點也相似。植物群落中不同組成成分生活型的劃分,是研究群落結構的基礎。采用瑙基耶爾(Raunkiaer)的生活型系統對不同演替階段植物生活型進行劃分,具體劃分結果見表1。
從表1可以看出,各演替階段植被均以高位芽植物占優勢,其在各演替階段的生活型譜中所占的比例分別為34.3%、55.7%、51.5%、67.9%、62.5%、64.7%。高位芽中主要以矮、小高位芽為主,其次為中高位芽,很少有大高位芽分布,層間分布有大量的藤本植物,高位芽植物中常綠的平均占21.6%,落葉的平均占34.5%,常綠中各高位芽比例大小為小高位芽>矮高位芽>大高位芽>中高位芽,而落葉中各高位芽比例大小為小高位芽>中高位芽>矮高位芽>大高位芽;草本植物中以地面芽植物為主,平均所占比例為18.4%,其次分別為一年生植物、隱芽植物,還分布有少量地上芽植物。群落演替過程實質就是不同物種間相互替代的過程,促使群落朝頂級群落發展,但從群落整個生活型譜中不難發現,以小、矮高位芽占優勢的群落,其無法形成高大的頂級喬木林群落,故需要適時地引入一些大高位芽植物,完善群落整體結構,促進其朝頂級群落演替。
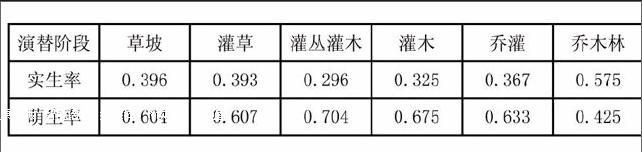
3.1.3 群落起源分析 從表2可以看出,植株實生比例呈現“V”字型變化趨勢,在灌叢灌木階段達到最低,僅為0.296,此階段主要以萌生的植株為主,出現一叢多株的現象,植株長勢較差,粗度小,高度低,而株數密度大,演替到更高一級群落階段所需的時間長,大多需要輔以人工措施,進行適宜疏伐,多留實生植株,到了灌木階段,木本植物大量增多,密度達到了最大值,物種對有限資源的競爭日益加劇,而實生植株具有根系發達、生長旺盛的特點,比萌生植株具有更強的競爭力,從而存活了下來,故其比例開始增大,并在喬木階段達到最大值,高達0.575,占到總植株數的一半之多,實生植株所具有的生長優勢也在植被恢復后期得以體現。
表2 不同演替階段植物起源統計
3.2 群落結構分析
森林群落的結構是指群落的組成結構、垂直結構和水平空間結構[5]。從表3可以看出林分在垂直結構上,隨著植被演替進展,喬木層高度、蓋度均逐漸增大,其中喬木層高度在喬木林階段突然上升,但高度還是較低,平均高度僅為6.64米;灌木層蓋度總體呈現先上升再下降的趨勢,在喬木林階段卻有所上升,可能原因是該區域的喬木林曾遭到人為砍伐,出現多處林窗,給林下灌木生長提供了有利條件,其次由于母樹天然下種,林下分布有大量的實生幼苗,導致灌木蓋度較大;草本層蓋度變化與喬木層基本相反,只是在進入喬灌階段以后,由于生境條件有所改善,林分尚未完全郁閉,給林下一些耐陰性較強的草本提供了生長空間,蓋度有所回升,但總體蓋度相對較低。
從水平結構上看,群落植株密度總體呈現先上升后降低,在灌叢灌木階段達到最大值,但灌木地徑較小,僅為0.71厘米,當進入喬灌和喬木林階段時灌木層植株密度依然很大,群落結構較差,需要進行適當的撫育,降低灌木層植株密度,優化群落結構。
3.3 群落物種多樣性
物種多樣性是指種的數目及其個體分配均勻度兩者的綜合,它能有效地表征生物群落和生態系統結構的復雜性[6]。從表4可以看出,喬木層豐富度指數呈現上升趨勢,但由于喬木層物種較為單一,豐富度指數相對較低,灌木層豐富度指數總體呈現上升趨勢,在喬木林階段達到最大值,草本層豐富度指數呈現先下降,在灌木階段達到最小值,而后又有所回升;喬木層的Simpson多樣性指數和Shannon-wiener多樣性指數均呈上升趨勢,而灌木層呈現先上升后下降,而后又上升的趨勢,在喬木林階段達到最大值,分別達到0.9119和2.9010,草本層多樣性指數在草坡階段達到最低,而后上升,在進入灌叢灌木后又有所回落,當進入喬灌階段后有又所回升;喬木層均勻度指數總體呈現上升趨勢,而灌木層呈“V”型變化,在灌叢灌木階段達到最小值,草本層的均勻度變化較不明顯,只是在草本階段相對小些,主要是由于草本群落大多是由火燒跡地演替而來,主要是由白茅、密毛蕨等物種組成,這些草且高度均較大,抑制了其他草本的生長,導致分布不均,多樣性指數低。
3.4 群落生物量分布特征
生物量的積累是退化喀斯特森林生態系統生產力發展的結果,也是喀斯特森林生態系統功能恢復的基礎[7]。
退化喀斯特森林植被主要以早期的灌叢灌木為主,對于研究小徑階的灌木植被生物量計算模型具有極其重要的意義,本研究共砍36種共107株樣木,通過生物量與地徑平方和高的研究,建立地上部分生物量與地徑的相對生長關系預測模型擬合,得出公式:
灌木層生物量采用以上公式進行計算。
從表5可以看出,群落總生物量呈逐漸上升趨勢,由于研究區植被常年遭到人為破壞,植被演替進程緩慢,所形成的喬木林大多為早期型喬木林群落,植被總生物量小;草本層生物量呈現先上升后下降,進入喬灌階段以后又略微有所回升的趨勢,而后林分開始郁閉,形成了高大的喬木林,林下的植被難以獲取足夠的光照,大量死亡,僅剩下一些耐陰性較強的草本能夠生長,草本層生物量達到最低值,僅為1.924/公頃;灌木層生物量呈現先上升,后下降的趨勢,在草坡階段由于很少有木本植物分布,灌木層生物量最小,僅為0.648噸/公頃,隨著生境條件的改善,在灌草階段出現了一些先鋒樹種,灌木層生物量開始上升到3.209噸/公頃,到灌叢灌木階段便迅速上升到10.067噸/公頃,而后在灌木段階達到最大值,進入喬灌階段以后,一些優勢樹種迅速長成喬木,不斷占用營養資源,抑制了其他灌木樹種的生長,灌木層生物量開始下降喬木層生物量變化趨勢與群落總生物量相同,在部分灌叢灌木階段便分布有少許喬木,這些喬木種類比較單一,大多為馬尾松,并且大多為以前人工栽植的,被人們砍伐后所遺留的干型長勢較差的喬木,隨著植被恢復到更高階段,特別是喬灌階段,植被開始分化,一部分競爭力較強的樹種生長變成喬木,喬木層生物量開始增大,在此階段初步形成喬、灌、草三層的群落結構。從每個層生物量所占比重中可以發現,草本層生物量比重總體呈下降趨勢,在喬木林階段達到最低,僅為5.96%,灌木層生物量比重呈現先上升后下降的趨勢,在灌木階段達到最大值,為57.96%,喬木層生物量比重呈單調上升趨勢,在喬木林階段達到最大值,但由于喬木林多為早期型,高大的喬木較少,喬木層生物量比重也僅為54.66%,從不同演替階段不同層次生物量分布情況可以看出,示范區大多數群落的生物量分布不合理,尚未形成合理的喬灌草層結構,需要進行適當的結構調整。
4 結語
雖然示范區的不同演替階段群落物種相對豐富,但其群落組成結構較不合理,生活型主要以矮、小高位芽為主,群落層次結構存在較大問題,各層生物量分布不合理,灌木株數密度過高,而喬木樹種稀少,所形成的喬木林大多以早期型為主,群落總體處于一個較低的演替階段,需要進行結構調整,優化物種組成結構,才能促進群落朝頂級群落演替。
參考文獻
[1] 何才華,熊康寧,粟茜.貴州喀斯特生態環境脆弱性類型及其開發治理研究[J].貴州師范大學學報(自然科學版),1996,14(1):1-9.
[2] D.Muller-Dombois H.Ellenberg著.(鮑顯誠等譯).植被生態學的目的和方法[M].北京:科學出版社,1986:247-252.
[3] 朱守謙,魏魯明,陳正仁,等.茂蘭喀斯特森林生物量構成初步研究[J].植物生態學報,1995,19(4):358-367.
[4] 楊持.生態學實驗與實習[M].北京:高等教育出版社,2003.
[5] 胡傳偉,孫冰,陳勇,等.深圳次生林群落結構與植物多樣性[J].南京林業大學學報(自然科學版),2009,33(5):21-26.
[6] 沈年華,萬志洲,湯庚國,等.紫金山栓皮櫟群落結構及物種多樣性[J].浙江林學院學報,2009,26(5):696-700.
[7] 劉映良,薛建輝.貴州茂蘭退化喀斯特森林群落的數量特征[J].南京林業大學學報(自然科學版),2005,29(3):23-27.
作者簡介:蔡品迪,碩士學歷,貴州林業勘察設計有限公司,助理工程師,研究方向:林業調查規劃設計。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
