利用隸屬函數法對不同基因型甘薯耐鹽堿能力的分析與評價
2015-03-02 03:37:16張勝利朱弘博韓忠才
東北師大學報(自然科學版) 2015年2期
關鍵詞:植物
孫 凱,張勝利,朱弘博,韓忠才,徐 飛
(吉林省蔬菜花卉科學研究院,吉林 長春130033)
土壤的鹽堿化是一個世界性的生態問題,全球超過10%的陸地正遭受著土壤鹽堿化的侵蝕.根據聯合國糧農組織以及教科文組織的不完全統計,全世界鹽堿地的面積約為9.54億hm2,其中我國約為1.3億hm2[1].土壤鹽堿化程度加重,次生鹽堿土地面積的不斷擴大,嚴重影響和制約了農業的可持續發展.在人口不斷增長和能源緊缺的今天,如何利用鹽堿化土地,已成為政府和科學界關注的熱點問題[2].
甘薯(Ipomoea betatas Lam.),也稱為地瓜、甜薯、紅苕、番薯等,屬旋花科甘薯屬草本植物,是重要的糧食作物、能源原料作物和飼料作物,此外也是一種耐逆性較強的作物.已有的研究主要集中在甘薯的中性鹽脅迫上,表明不同甘薯品種的耐鹽程度差別很大[3-4];有關甘薯對鹽堿混合脅迫下的研究和品種篩選目前報道的還較少.鹽堿脅迫是包含了堿性鹽脅迫和中性鹽脅迫的混合脅迫,鹽堿脅迫下植物不僅受Na+的毒害,還會受高pH 和低水勢的脅迫[5].因此本實驗以5個吉林省主栽的甘薯品種為材料,研究了鹽堿脅迫下甘薯的生理指標變化,評價了不同品種的耐鹽堿性,旨在研究鹽堿混合脅迫下甘薯的生理特性變化,以為甘薯的耐鹽堿品種的篩選和鹽堿地栽培提供參考.
甘薯的耐鹽堿性受多種基因影響,是多個因素相互作用而構成的綜合反應體系,由于每一項指標不能一概地反應甘薯的耐鹽堿性,單項的機理研究又存在一定的局限性,也不具有說服力,因此綜合多個指標進行研究分析才可以使實驗結果更加客觀.隸屬函數法是模糊集合的特征函數,利用隸屬函數法對植物的抗逆性進行分析在玉米、馬鈴薯、水稻等作物的抗逆性研究中都有報道.本文通過求出各耐鹽堿性指標在品種中的隸屬函數值,再通過平均值來評價各個品種的耐鹽堿性,對甘薯的耐鹽堿性進行了綜合的分析與評價,以期為耐鹽堿指標的選擇提供一種可行的方法,同時為培育優良的耐鹽堿甘薯品種提供理論依據.
1 材料與方法
1.1 實驗材料
供試材料為吉徐薯1號、吉徐薯2號、吉1114、徐紫薯2號、徐薯28,均由吉林省農業科學院經濟植物研究所惠贈.
1.2 實驗設計
采用沙培方式種植薯苗,選取生長良好、長勢一致的甘薯苗為試材.薯苗扦插后第一周作為緩苗期,早晚各澆1次Hoagland營養液,澆灌量為細沙持水量的2倍;扦插后第二周開始處理,用Hoagland營養液將NaHCO3和Na2CO3按照n(NaHCO3)∶n(Na2CO3)=5∶1的比例配制成鹽濃度為100mmol/L的溶液作為鹽堿處理液,對照組使用Hoagland營養液,處理后每隔2d取樣測量各個指標.
1.3 測定指標及方法
植株鮮干重、葉綠素含量、相對電導率、丙二醛含量、可溶性蛋白含量、可溶性糖含量、SOD 活性根據張治安和張美善報道的方法進行測定[6].
1.4 綜合評價
采用隸屬函數法對各個品種進行耐鹽堿性評價.即
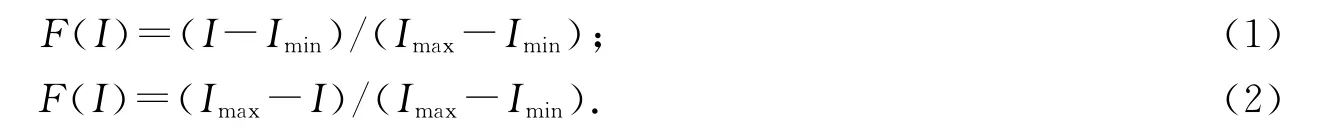
式中:F(I)為隸屬函數值;I 為指標的測定數值;Imax,Imin分別為實驗品種的某一指標的最大值和最小值.當某指標與植物耐鹽堿能力成正相關時采用公式(1),當某指標與植物耐鹽堿能力成負相關時采用公式(2);最后將各個指標的隸屬函數值求和計算平均值.隸屬函數值越大說明植株受到危害的程度越大,即耐鹽堿性越弱.通過比較平均值的大小,即可評價各甘薯品種抗鹽堿性的強弱.
1.5 數據分析
實驗數據用Microsoft Excel 2010(Microsoft)和SPSS 19.0(IBM SPSS)軟件進行分析處理.
2 結果與分析
2.1 鹽堿脅迫對不同基因型甘薯植株葉綠素含量的影響
鹽堿脅迫對不同基因型甘薯品種植株葉綠素含量的影響結果見表1.由表1可見,不同基因型的甘薯在鹽堿脅迫下葉綠素的含量均出現下降現象,但各品種間存在差異.與對照相比,徐紫薯2號的葉綠素含量下降的最小,吉徐薯1號和吉徐薯2號下降較大,徐薯28和吉1114居中.鹽堿脅迫下,植物吸收水分困難,葉片發黃,植株葉綠素含量有所降低,這也影響了植物的光合作用,進而使植物生長緩慢.
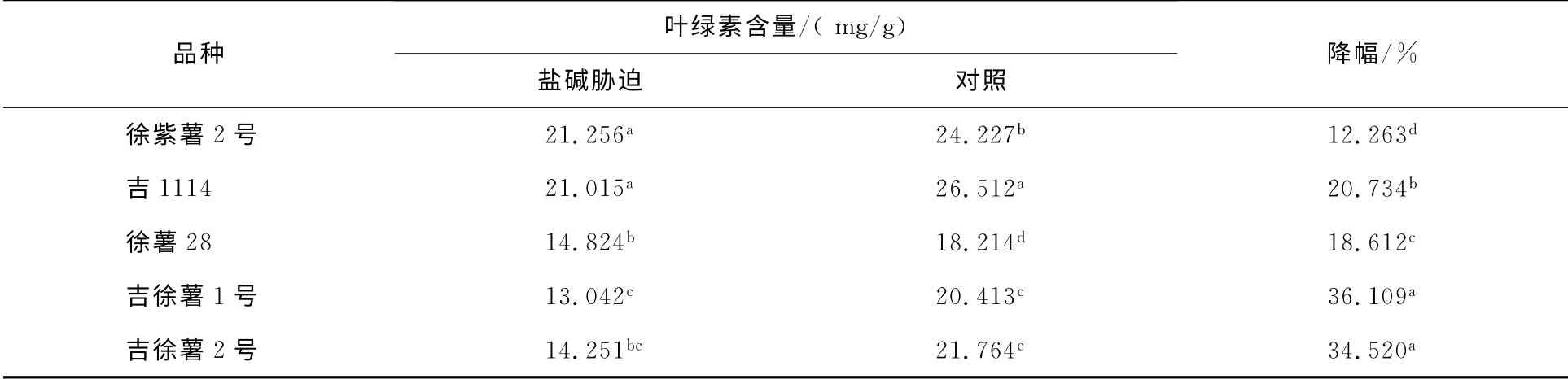
表1 鹽堿脅迫對不同基因型甘薯幼苗葉綠素含量的影響
2.2 鹽堿脅迫對不同基因型甘薯相對電導率的影響
鹽堿脅迫對不同基因型甘薯品種相對電導率的影響結果見表2.由表2可見,鹽堿脅迫下,5個甘薯品種的相對電導率均出現增大現象,其中吉徐薯2號增大幅度最大,比對照增大了76.52%;吉徐薯1號增大了63.16%;吉1114增大幅度最小,增大了19.06%;徐紫薯2號和徐薯28的相對電導率增大幅度居中,且差異性不顯著.植株相對電導率的變化可以反映出細胞受傷害的程度,5個甘薯品種中,吉徐薯2號的相對電導率最高,說明細胞受傷害的程度最為嚴重.
2.3 鹽堿脅迫對不同基因型甘薯丙二醛含量的影響
鹽堿脅迫對5個甘薯品種丙二醛含量的影響結果見表3.由表3可見,在鹽堿處理下,不同基因型甘薯的丙二醛含量均出現增大現象,其中吉1114增加最少,且與其他品種間差異顯著;吉徐薯1號和吉徐薯2號增加較大,兩品種間差異不顯著;徐紫薯2號的丙二醛含量大于徐薯28的,且達到了差異顯著的水平.植株體內丙二醛含量的變化可以反映植株受危害的程度,鹽堿脅迫下吉1114丙二醛含量最低,吉徐薯1號最高,說明吉1114受到的危害最小,而吉徐薯1號最大.

表2 鹽堿脅迫對不同基因型甘薯幼苗細胞膜透性的影響

表3 鹽堿脅迫對不同基因型甘薯幼苗丙二醛含量的影響
2.4 鹽堿脅迫對不同基因型甘薯植株鮮重、干重的影響
鹽堿脅迫對5個甘薯品種鮮重、干重的影響結果如圖1所示.由圖1可見,鹽堿脅迫抑制了5個甘薯品種的生長,但對每個品種的抑制程度不同,其中吉徐薯1號的植株鮮重比對照減少60.16%;徐薯28的植株鮮重減小幅度最小,為31.81%;吉徐薯2號的植株干重減小幅度最大為79.81%;吉1114的植株干重減小幅度最小,為44.04%.從甘薯植株鮮、干重的變化可以看出,吉1114的耐鹽堿性比較強,而吉徐薯1號和吉徐薯2號較弱.
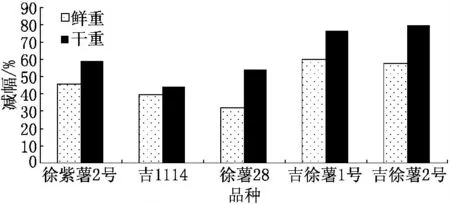
圖1 鹽堿脅迫對不同品種甘薯 幼苗鮮重、干重的影響
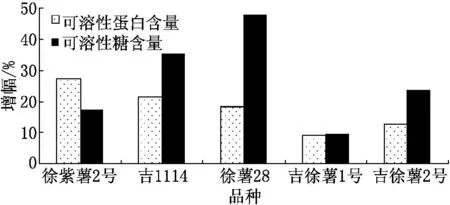
圖2 鹽堿脅迫對不同甘薯品種可溶性蛋白、 可溶性糖含量的影響
2.5 鹽堿脅迫對不同基因型甘薯可溶性蛋白、可溶性糖含量的影響
鹽堿脅迫對不同基因型甘薯植株體內可溶性蛋白、可溶性糖含量的影響結果見圖2.由圖2可見,鹽堿脅迫下甘薯植株體內的可溶性蛋白、可溶性糖含量均有所增加.與對照相比,5個甘薯品種可溶性蛋白含量的增幅在9.22%~27.34%的范圍內,吉徐薯1號增加幅度最小,徐紫薯2號增加幅度最大;可溶性糖含量增加幅度差異較大,徐薯28增加幅度最多達到47.87%,吉徐薯1號最小為9.42%.
2.6 鹽堿脅迫對不同基因型甘薯保護性酶類的影響
鹽堿脅迫下甘薯植株體內的SOD 活性(鮮樣)呈先升高后降低的趨勢,5個甘薯品種的變化基本趨勢相同.如圖3所示,在第3次取樣時達到峰值,吉1114的峰值最高,且與其他品種間均達到差異顯著的水平,其次為徐紫薯2號和徐薯28號,但差異并不顯著;從第4次取樣開始各品種的SOD 活性開始下降;最后一次取樣時,吉1114的SOD 活性最高,吉徐薯2號的SOD 活性最小.
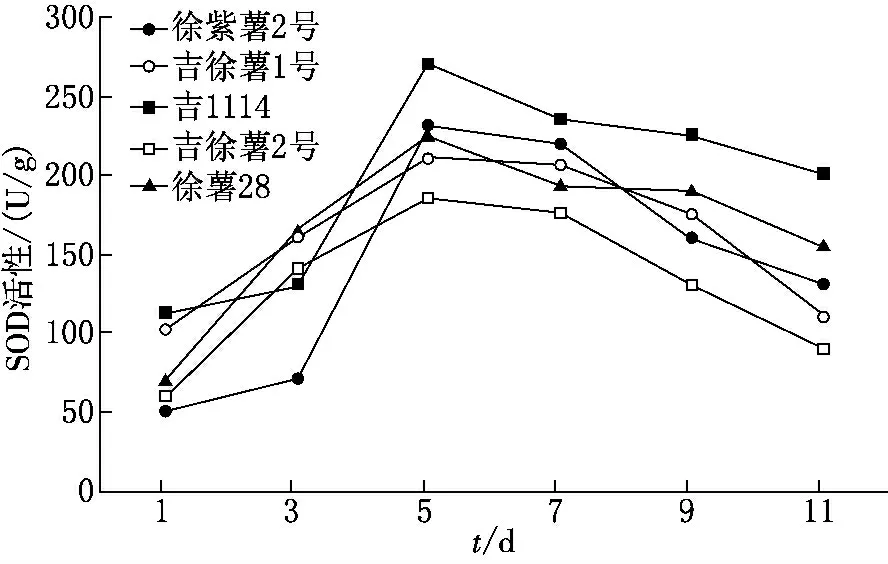
圖3 鹽堿脅迫對不同甘薯品種SOD活性的影響
2.7 不同基因型甘薯的耐鹽堿性評價
以7個指標的變化幅度為依據,利用隸屬函數法分析5個甘薯品種的耐鹽堿能力,結果見圖4.如圖4所示,不同基因型甘薯品種的耐鹽堿能力存在較大差異.
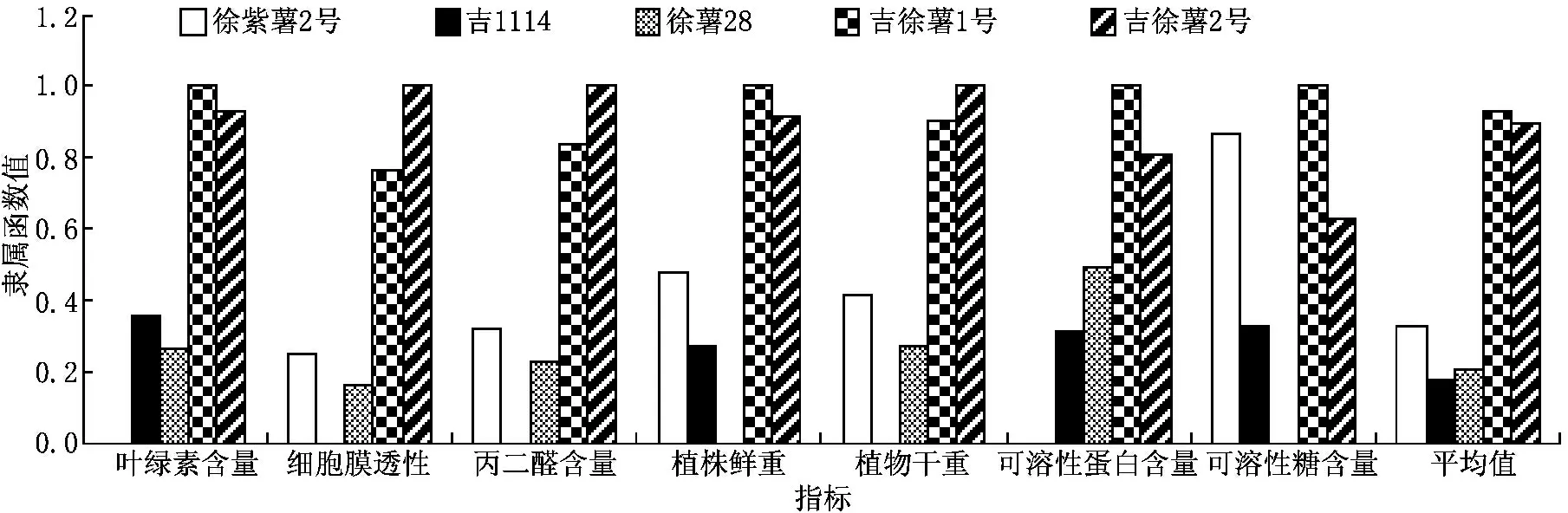
圖4 5個甘薯品種耐鹽堿能力綜合評價
3 討論及結論
研究表明,鹽堿脅迫引發了甘薯植株體內一系列的生理響應機制,對甘薯具有抑制生長的效應,降低了植株的鮮重、干重、葉綠素含量等指標,且不同品種間存在著明顯的差異.本文以丙二醛含量、植株鮮干重、葉綠素含量、細胞膜透性等7個指標的變化為依據,運用隸屬函數法分析了5個甘薯品種的耐鹽堿能力,根據綜合指數的排序,其耐鹽堿能力強弱的綜合排序為:吉1114>徐薯28>徐紫薯2號>吉徐薯2號>吉徐薯1號.
有研究表明,鹽堿脅迫提高了植株葉綠素酶的活性,進而加快了葉綠素的降解,導致葉綠素含量減少[7];同時降低了植株碳的同化量,從而降低了植株的光合效率,抑制了植株的生長[8].大量實驗證明,生物量可以綜合體現出植物的耐鹽堿性,是評價植物耐鹽堿性的可信指標[9-10],在番茄(Lycopersicon esculentum Mill.)、大豆(Glycine max)、水稻(Oryza glaberrima)、小麥(Triticum aestivum Linn.)等植物的研究中已有報道[11-18],結果與本文中甘薯的干重、鮮重、葉綠素含量均有所降低的結論一致.
相對電導率是反映植物細胞膜系統穩定性的重要指標,在逆境條件下,膜蛋白會因細胞膜受到破壞使細胞質外滲,進而導致相對電導率加大,增幅越大傷害程度就越大[19],反映較為直接.本文中,吉1114的相對電導率增幅最小,反映出其具有一定的耐鹽堿能力,這與最后耐鹽堿性綜合評價的結果一致.
鹽堿脅迫下,高Na+的存在使植株吸水困難,植物細胞膜系統也受到一定的破壞而發生失水,植株就會產生一些生理響應機制,合成新的可溶性蛋白、可溶性糖以加強滲透調節作用,增強植株的抗鹽堿性.本實驗中,5個甘薯品種的可溶性蛋白、可溶性糖含量在受到鹽堿脅迫后都有所增加,反映出甘薯具有一定的耐鹽堿性,但也因品種的不同存在差異.
植株在逆境時,會積累較多的活性氧,丙二醛是膜脂過氧化的終產物之一,其含量高低可以作為細胞受脅迫程度的指標之一,它的主要傷害是導致膜脂過氧化,損傷生物膜結構;與此同時植物體內保護性酶活性會應激提高以保護植物細胞免受活性氧的損傷[20-21].本研究中,5個甘薯品種的丙二醛含量均升高,但程度不同,說明不同品種受到傷害的程度不同.甘薯體內活性氧清除系統相關酶活性明顯受到鹽堿脅迫的影響,SOD 活性應激升高,之后逐漸降低,這說明甘薯植株自身有著比較強的生理適應性,在一定程度上,能有效清除體內產生的活性氧,最大程度減少甘薯所受到的傷害,且品種間存在著差異.
本研究采用隸屬函數法對5個甘薯品種的7項指標進行了綜合耐鹽堿性評價,評價消除了個別指標帶來的片面性,較客觀地評價了不同基因型甘薯的耐鹽堿性.但是此次研究尚未考慮每項指標的貢獻率問題,有待在今后的進一步研究中結合聚類分析法、隸屬函數法、主成分分析法、灰色關聯度法以及數據標準化法等[22-23]進行論證,使抗逆性的研究結果更加明確、客觀.
[1]肖娟.水質對鹽堿地水鹽耦合運移和作物生長影響的研究[D].北京:中國農業大學,2004.
[2]趙可夫.植物抗鹽生理[M].北京:中國科學技術出版社,1993:3-4,136-138.
[3]孫曉波,謝一芝,馬鴻翔.甘薯幼苗對海水脅迫的生理生化響應[J].江蘇農業學報,2008,24(5):600-606.
[4]柯玉琴,潘廷國,NaCl脅迫對甘薯苗期生長、IAA 代謝的影響及其與耐鹽性的關系[J].應用生態學報,2002,13(10):1303-1306.
[5]EI S H M A,SHADDAD M A K.Comparative effect of sodium carbonate,sodium sulphate,and sodium chloride on the growth and related metabolic activities of pea plants[J].J Plant Nutrition,1996,19(5):717-728.
[6]張治安,張美善,蔚榮海,等.植物生理生化實驗指導[M].北京:中國農業科學技術出版社,2004.
[7]高永生,王鎖民,龔海軍.鹽脅迫下植物離子轉運的分子生物學研究[J].草業學報,2003,12(5):18-25.
[8]GRIEVE C M,LESCH S M,MAAS E V,et al.Leaf and spikelet primordia initiation in salt-stressed wheat[J].Crop Science,1993,33:1286-1292.
[9]VICENTE O,BOSCAIU M,NARANJO M A,et al.Responses to salt stress in the halophyte Plantago crassifolia(Plantaginaceae)[J].Journal of Arid Environments,2004,58(4):463-481.
[10]LEVITT J.Response of plants to environmental stress[M].New York:Academic Press,1980:365-434.
[11]羅慶云,於丙軍,劉友良.大豆苗期耐鹽性鑒定指標的檢驗[J].大豆科學,2001,20(3):177-182.
[12]劉祖祺,張石城.植物抗性生理學[M].北京:中國農業出版社,1994:278.
[13]李倩中,蘇家樂,劉曉宏,等.4 種槭屬植物耐鹽性差異的研究[J].江蘇農業科學,2009(6):227-228.
[14]趙鎖勞,竇延玲.小麥耐鹽性鑒定指標及其分析評價[J].西北農業大學學報,1998,26(6):80-85.
[15]鐘小仙,鄒 軼,張建麗,等.海鹽脅迫對海濱雀稗植株形態與生長的影響[J].江蘇農業科學,2009(6):235-236.
[16]祁棟靈,韓龍植,張三元.水稻耐鹽/堿性鑒定評價方法[J].植物遺傳資源學報,2005,6(2):226-231.
[17]湯日圣,黃益洪,唐現洪,等.生物源脫落酸對鹽脅迫下辣椒苗生長和某些生理指標的影響[J].江蘇農業學報,2009,25(4):856-860.
[18]WALTER L.植物生理生態學[M].翟志席,郭玉海,馬永澤,譯.北京:中國農業大學出版社,1997:305.
[19]肖雯,李恢先,蒲陸梅.幾種鹽生植物抗鹽生理指標的研究[J].西北植物學報,2000,20(5):818-825.
[20]?TAJNER D,POPOVI□M,?TAJNER M.Herbicide induced oxidative stress in lettuce,beans,pea seeds and leaves[J].Biologia Plantarum,2003,47:575-579.
[21]呂金印,郭 濤.水分脅迫對不同品種甜高粱幼苗保護酶活性等生理特性的影響[J].干旱地區農業研究,2010,28(4):89-93.
[22]孟慶立,關周博,馮佰利,等.谷子抗旱相關性狀的主成分與模糊聚類分析[J].中國農業科學,2009(8):2667-2675.
[23]張衛星,趙致,柏光曉,等.不同基因型玉米自交系的抗旱性研究與評價[J].玉米科學,2007,15(5):6-11.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
