宮頸癌中CSN6對E6AP的調節作用及其臨床意義
2015-03-11 02:15:35王超宋昱郭奇桑劉艷梅高蜀君
中國臨床醫學 2015年3期
關鍵詞:水平
王超 宋昱 郭奇桑 劉艷梅 高蜀君
(復旦大學附屬婦產科醫院婦科,*宮頸疾病診療中心,上海 200011)
?
·論著·
宮頸癌中CSN6對E6AP的調節作用及其臨床意義
王超宋昱*郭奇桑*劉艷梅*高蜀君*
(復旦大學附屬婦產科醫院婦科,*宮頸疾病診療中心,上海200011)
摘要目的:探討宮頸癌中組成型光形態發生因子9信號復合體亞單位6(constitutive photomorphogenesis 9 signalosome subunit 6,CSN6)對E6AP的調節作用。方法: 在癌癥和腫瘤基因圖譜(The Cancer Genome Atlas,TCGA)數據庫中檢索CSN6和E6AP基因,比較分析其相關性。采用質粒轉染、蛋白質印跡(Western blotting)、四甲基偶氮唑藍(methyl thiazolyl tetrazolium,MTT)、流式細胞術等方法檢測CSN6對E6AP表達及細胞生物學的影響。通過在裸鼠皮下接種不同CSN6表達狀態的Hela細胞株建立動物模型,觀察CSN6對腫瘤生長的影響。結果:數據庫資料顯示,CSN6和E6AP在腫瘤中有過表達和突變的現象。CSN6可以上調E6AP蛋白的表達,抑制CSN6的表達可以使E6AP表達下降,細胞增殖能力降低,細胞周期停滯于G0-G1期。動物實驗顯示,敲除CSN6可以顯著抑制腫瘤生長,且腫瘤中E6AP蛋白表達水平下降,p53蛋白表達水平增加。結論:CSN6可以通過上調E6AP蛋白的表達而促進宮頸癌的發生發展。
關鍵詞宮頸癌;人乳頭瘤病毒;感染;組成型光形態發生因子9信號復合體;E6AP
宮頸癌是女性生殖系統三大常見惡性腫瘤之一,其發病率在女性所有惡性腫瘤中位居第四,在15~44歲年齡段的女性的惡性腫瘤中位居第二。2012年的數據統計顯示,全世界每年新增宮頸癌病例數527 624例,死亡265 653例[1]。目前我國宮頸癌發病率高居世界第2位,僅次于智利,且發病呈年輕化趨勢明顯。
宮頸癌的發病與持續性的人乳頭瘤病毒(human papilloma virus,HPV)感染關系密切[2-3]。至少15種HPV亞型被列入高危型HPV(high risk HPV,HR-HPV),可引起生殖道及非生殖道的惡性腫瘤,16和18型HPV在超過70%的宮頸癌中可被檢測出[4-5]。HR-HPV E6和E7蛋白是其編碼的主要癌蛋白。HR-HPV E6參與了多條凋亡通路的調控,其中最為經典的是E6通過E6AP介導的p53泛素化降解。E6AP作為泛素連接酶,可與E6和p53形成三聚體,從而特異性介導p53的泛素化降解,阻斷凋亡信號的傳遞[6]。
組成型光形態發生因子9信號復合體(constitutive photomorphogenesis 9 signalosome,COP9 or CSN)是一種存在于真核細胞中的結構高度保守的多亞基蛋白復合體,與組成型光形態發生(constitutive photomorphogenesis,COP)現象密切相關[7]。CSN與26S蛋白酶體的19S“蓋子”復合物具有同源性;19S“蓋子”被認為具有識別泛素化底物并將其傳送至蛋白水解核心復合物進行降解的功能[8];CSN被認為是連接信號傳導和蛋白降解的分子平臺[9],在調控多聚泛素鏈降解中具有重要的催化作用[10]。大量的研究[11-13]證明:CSN能通過調控細胞的分裂和分化、影響信號傳導、活化細胞轉化及調節DNA損傷應答等多種途徑參與腫瘤的發生及發展;CSN的2個亞單位CSN5和CSN6在多種惡性腫瘤(例如乳腺癌[14]、肺癌[15-16]、甲狀腺癌[17]等)中呈高表達且影響腫瘤的發生及發展。
CSN6可以抑制MDM2的自身泛素化,使其在細胞中穩定表達,持續發揮泛素化降解p53的作用,導致p53對細胞生長、凋亡和DNA修復的調控作用丟失,促進腫瘤形成[18]。有研究[19]證實,在宮頸癌中,MDM2-p53通路因HR-HPV E6的參與而特異性轉換為E6-E6-AP-p53通路。但目前宮頸癌中E6AP的上游調節機制尚不明確,CSN6是否同樣可以通過E6AP促進p53的降解,目前尚無相關報道,CSN6在宮頸癌中的作用也未見報道。因此,本研究探討了CSN6在宮頸癌中的作用以及其對E6AP和p53的影響。
1資料與方法
1.1細胞和主要試劑HPV陽性細胞系(Hela和Caski)、HPV陰性細胞系(C33a)以及正常細胞(HEK-293T)購自美國標準生物品典藏中心(ATCC)。DMEM F12培養基(含10%胎牛血清)購自美國Gibco公司。MG132(C2211)購自美國Sigma公司。抗體來源如下:CSN6、 E6-AP購自瑞士ENZO life Sciences公司;p53、Flag(M2 monoclonal antibody)、Actin購自美國Sigma公司;Myc購自英國Abcam公司。Flag-CSN6和Myc-CSN6見參考文獻[18]。shRNA-CSN6、N-CSN6(1-170 aa)、C-CSN6(160-327 aa)、CSN6(S60A)和CSN6(S60D)突變體的構建見參考文獻[15,20-21]。
1.2shRNA-CSN6穩定細胞株的構建每60 mm細胞培養皿中鋪入4×105個Hela細胞,18 h后進行細胞轉染。重組慢病毒質粒GIPZ12-pshRNA-CSN6和陰性對照質粒轉染采用Lipofectamine TM 2000轉染試劑,用2 μg/mL嘌呤霉素(puromycin)進行篩選,每隔48 h更換一次新鮮培養基,維持puromycin的濃度,持續培養至14 d左右可形成細胞克隆,挑選克隆進行擴大培養,以備后續實驗使用。
1.3蛋白質印跡(Western blotting)和免疫共沉淀法吸凈培養液,用磷酸鹽緩沖液(phospahate buffered saline,PBS)小心漂洗2次,加入預冷的含有蛋白酶/磷酸化酶抑制劑的1×裂解緩沖液[0.5L 1×裂解緩沖液組成成分:7.5 g 1 mol/L Tris(美國Fisher公司),15 mL 5 mol/L NaCl(美國Fisher公司),0.5 mL NP-40(美國USB Corp公司),0.5 mL TritonX-100(美國Sigma公司)和1 mL 0.5 mol/L乙二胺四乙酸(美國Fisher公司)。蛋白酶/磷酸化酶抑制劑組分:5 mmol/L礬酸鈉(NaV),1 mmol/L氟化鈉(NaF),1 μmol/L 二硫蘇糖醇(DTT),0.1 mg/mL胃蛋白酶抑制劑(Pepstatin)A,1 mmol/L苯甲基磺酰氟(phenylmethylsulfonyl fluoride)和1000×全蛋白酶抑制劑混合物(瑞士Roche公司)], 于4 ℃或冰上放置20 min裂解細胞,4 ℃以13 000 r/min離心15 min。將離心后的上清液分為2份:1份加入6×十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)樣品緩沖液,混勻后于100 ℃煮沸10 min,作為總細胞裂解液于-20 ℃保存;另1份用于免疫沉淀,1 mL細胞裂解液加入10 μL Protein A/G瓊脂糖珠,于4 ℃共振1 h,以10 000 r/min離心10 min,取上清備用,將瓊脂糖珠分別與目的抗體、IgG抗體(陰性對照)混合并加入上面的上清液中,混勻器勻速旋轉(4 ℃過夜);將免疫沉淀后的溶液于4 ℃以3000 r/min離心3 min,去上清液,用1×裂解緩沖液洗滌瓊脂糖珠,離心后棄上清液,共洗滌3次。將1×裂解緩沖液與6×SDS樣品緩沖液混合,于100 ℃煮沸10 min,作為免疫共沉淀樣本。最后將總細胞裂解液和免疫共沉淀樣本等量上樣于10%聚丙烯酰胺凝膠并電泳,電泳完畢后將蛋白轉移至聚偏氟乙烯膜,用5%牛血清白蛋白封閉1 h,加入一抗后4 ℃反應過夜。第2天洗膜后加入辣根過氧化物酶標記的二抗,室溫孵育1 h,用電化學發光法檢測。
1.4細胞增殖測定采用四甲基偶氮唑藍(methyl thiazolyl tetrazolium,MTT)法檢測細胞生長速率。按照每孔3×103個細胞的密度接種到96孔板,在37 ℃、CO2體積分數為5%的培養箱中培養。培養第1、2、3天后,分別將20 μ LMTT加入各孔中并繼續培養2 h,然后每孔加入二甲基亞砜100 μL,在570 nm波長處檢測。
1.5流式細胞術檢測細胞周期抑制CSN6表達48 h后,消化并收集細胞,調整細胞濃度為1×106/L,加入490 μL工作濃度的結合緩沖液,振蕩混勻細胞,同時加入5 μL Annexin V-FITC和5 μL碘化吡啶(propidium iodide,PI),室溫下避光染色20 min后用流式細胞儀進行雙參數分析,測定subG1率。用于檢測細胞周期的標本用70%乙醇固定后再用流式細胞儀檢測。
1.6動物模型選取4~5周齡雌性BALB/c裸鼠10只,來自德克薩斯MD安德森腫瘤中心動物飼養中心。將10只裸鼠隨機分為實驗組和對照組,各5只。實驗組裸鼠皮下接種shRNA-CSN6穩定細胞株(1×106個細胞重懸于0.1 mL PBS),對照組接種與實驗組等量的shRNA-VECTOR穩定細胞株。自接種第5天起,每周測量腫瘤體積3次,直至裸鼠接受安樂死。
2結果
2.1在臨床宮頸癌患者中CSN6及E6AP的mRNA表達情況及與主要分子蛋白的相關性在http://www.cbioportal.org網站的癌癥和腫瘤基因圖譜(The Cancer Genome Atlas,TCGA)數據庫里檢索CSN6和E6AP基因,結果發現,CSN6在大多數腫瘤中呈過表達(紅色),見圖1;而E6AP多以突變為主,見圖2。在宮頸鱗狀上皮癌和頸管內腺癌(cervical squamous cell carcinoma and endocervical adenocarcinoma,CESC)中發現,CSN6(基因名稱為COPS6)和E6AP(基因名稱為UBE3A)主要表現為mRNA水平的擴增,COPS6在宮頸癌中無突變,UBE3A有3%的突變,二者在宮頸鱗狀細胞癌和CESC中的表達呈負相關(pearson相關系數為-0.327,P<0.05),見圖3。從TCGA上下載所有CESC患者的CSN6和E6AP的mRNA水平信息,共253例患者,采用Prism 6.0t-test檢驗COPS6低水平組和高水平組的UBE3A表達差異,COPS6低水平組的UBE3A表達水平明顯高于COPS6高水平組(P<0.01),見圖4。同時下載所有CESC患者的反時相蛋白的陣列(reverse-phase protein lysate array,RPPA)數據,共155例患者,根據患者的序列號合并患者的COPS6、UBE3A及RPPA數據,運用R語言分別計算CSN6和E6AP與192種蛋白分子的相關性,COPS6與smad1、smad3、smad4、src-pY416、src-pY527、src、STAT3、STAT5、Stathmin蛋白分子水平呈負相關(P<0.05,見圖5),而UBE3A與192種蛋白分子沒有明顯相關性。

圖1 TCGA數據庫中CSN6 基因在腫瘤中的表達情況

圖2 TCGA數據庫中E6AP基因在腫瘤中的表達情況

圖3 TCGA數據庫中CSN6與E6AP基因表達的相關性

圖4 TCGA數據庫中253例CESC患者的

圖5 TCGA數據庫中CSN6 mRNA水平與RPPA中9種蛋白分子表達的相關性
2.2CSN6對E6AP表達的調節
2.2.1CSN6可與E6AP相互結合前述分析表明,CSN6和E6AP存在相關性。免疫共沉淀結果發現,宮頸癌Hela細胞株內的CSN6蛋白可以和E6AP蛋白結合,見圖6。抑制CSN6的表達后,E6AP的表達減少,見圖7。

(IP:免疫共沉淀;IB:免疫印跡;WCL:全細胞裂解液)
2.2.2CSN6對E6AP的調節依賴HPV的感染為進一步了解CSN6與E6AP的相關性,用myc-CSN6質粒轉染了4株宮頸癌細胞系:Hela和Caski
(合并HR-HPV感染),C33A(不合并HR-HPV感染,伴有p53突變)和人胚胎源腎上皮細胞株HEK-293T。結果顯示,HPV陽性細胞中,隨著外源性CSN6表達增加,E6AP表達水平呈劑量依賴性遞增;而在HPV陰性細胞中則無此現象;見圖8。

圖7 CSN6敲除對E6AP蛋白表達的影響
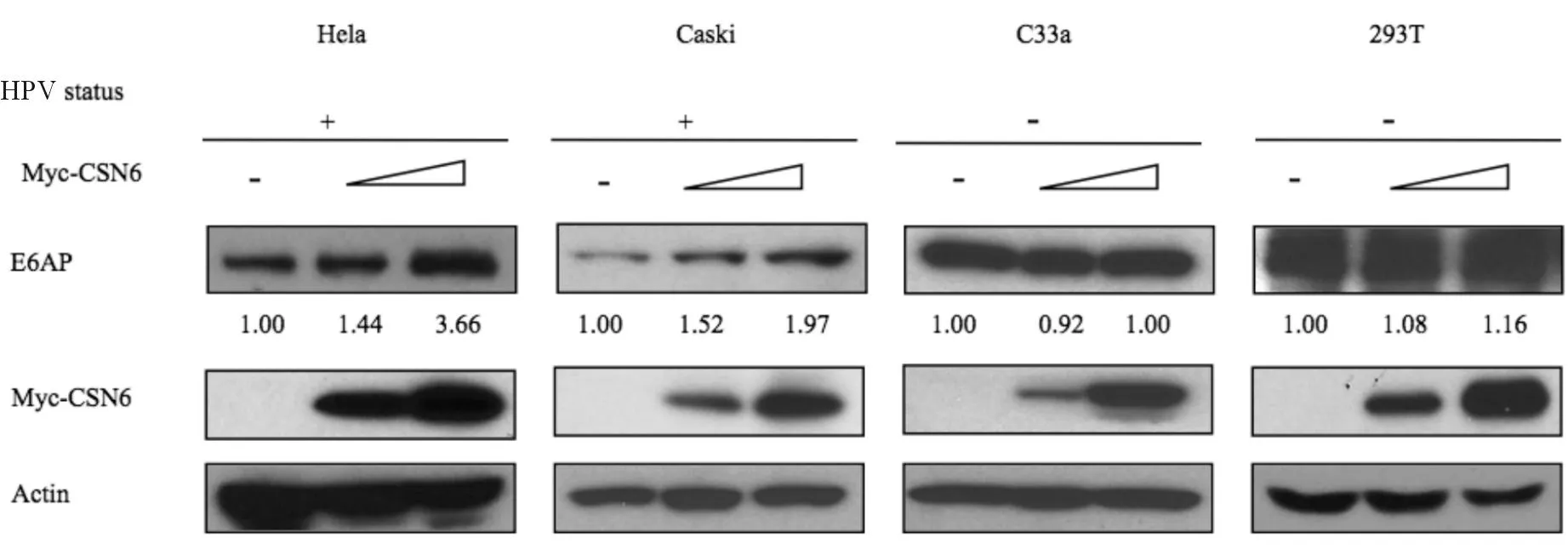
圖8 4種細胞系中過表達CSN6對E6AP蛋白表達的影響
2.3抑制CSN6后細胞增殖能力下降、凋亡增加MTT結果顯示,轉染shRNA-CSN6的Hela細胞的增殖能力明顯低于對照組,見圖9。流式細胞術檢測結果顯示,CSN6的敲除增加了細胞的凋亡率并促進細胞G0-G1期的停滯,見圖10a~10c。
2.4下調CSN6的表達可抑制腫瘤生長為進一步證實CSN6在宮頸癌中的作用,分別用感染CSN6 shRNA的Hela細胞和轉染對照空載體的Hela細胞接種裸鼠,觀察腫瘤的生長情況。結果顯示,CSN6敲除組腫瘤生長受到顯著抑制。Western blotting證實CSN6的敲除不僅降低E6AP蛋白表達水平,同時增加了p53蛋白表達水平,見圖11a、11b。

圖9 抑制CSN6對細胞增殖能力的影響(MTT)

a:抑制CSN6可促進細胞凋亡并促進細胞G0-G1期停滯(左下象限代表存活細胞,左上象限代表壞死細胞,右下象限代表早期凋亡細胞,右上象限代表晚期凋亡細胞);b:為a的統計柱狀圖;c:抑制CSN6促進細胞G0-G1期停滯
圖10流式細胞術檢測結果

a:宮頸癌移植成瘤實驗;b:Western blotting結果顯示,抑制CSN6可減少E6AP蛋白的表達水平,增加p53蛋白的表達水平,(#1~ #5分別為實驗組和實驗對照組和編號1~5 裸鼠)
圖11下調CSN6抑制腫瘤生長
3討論
目前認為,HPV的E6、E7蛋白是與宮頸癌關系最密切的2個癌蛋白,對E6、E7蛋白作用機制的研究有望為宮頸癌的靶向治療提供一定的理論依據。既往研究[22]證實,E6、E7蛋白均可以阻斷凋亡通路,實現細胞的永生化,而且E6的抗凋亡作用被認為與p53/PUMA/Bax通路密不可分。E6AP介導了E6對p53的泛素化降解過程,預示著對E6AP的調節可能會成為宮頸癌靶向治療的目標。本研究發現,與26S蛋白酶體的19S“蓋子”復合物具有同源性的CSN6可以正向調節HPV陽性細胞的E6AP水平,而對HPV陰性細胞沒有影響。CSN6表達水平越高,E6AP在細胞中越穩定,降低CSN6表達水平可增加E6AP蛋白的水解,因此,體外細胞實驗結果提示CSN6在蛋白水平上正向調節E6AP。這似乎與臨床患者中的mRNA水平分析結果不相吻合,253例宮頸癌患者的COPS6和UBE3A mRNA表達水平呈負相關。可能原因有:(1)細胞實驗結果顯示HPV陽性情況下CSN6正向調節E6AP,但253例患者中絕大多數未提供HPV是否感染的信息,僅少數注明HPV16或18型感染。因此,體外細胞實驗得出的結論并不適用于這253例患者;(2)體外實驗從蛋白水平進行研究,而臨床數據來自mRNA水平,由于體內mRNA和蛋白表達調控的復雜性,二者不存在可比性。
p53是迄今為止發現的與腫瘤相關性最強的基因之一,在細胞周期調控、抑制細胞生長、誘導腫瘤細胞凋亡等方面有重要作用。既往研究證實,CSN6可以通過多種途徑調節p53。例如有關CSN6-MDM2軸,研究[16,18]發現,人乳腺癌組織中CSN6與MDM2的表達呈正相關,CSN6與MDM2直接作用,作用的位點為MDM2的294-384aa,而MDM2自我泛素化的位點為賴氨酸364(位于MDM2與CSN6相互作用的區域內)。CSN6通過抑制MDM2的泛素化,增強MDM2的穩定性,導致p53的降解。此外,還有CSN6-14-3-3ó-Akt軸。14-3-3ó蛋白作為一種潛在的腫瘤抑制因子,是p53的下游靶點,可以負性調節Akt[23-24]。 研究[21]發現,CSN6能夠通過穩定COP1而介導14-3-3ó蛋白的泛素化降解,從而減少14-3-3ó蛋白對p53穩定性的正向作用,該功能與MDM2無關。此外,14-3-3ó蛋白的減少可激活Akt蛋白,激活的Akt通過磷酸化MDM2的絲氨酸166和186位點使其磷酸化,從而經MDM2途徑增強p53的泛素化和降解,促進細胞增殖。Xue等[15]還發現,Akt激酶可以磷酸化CSN6的絲氨酸60位點而使CSN6磷酸化,以減少泛素介導的CSN6降解,從而增加CSN6的穩態表達,進一步導致p53降解。本研究發現,除了以上途徑之外,CSN6還可以通過E6AP起到調節p53的作用。E6AP是泛素連接酶(ubiquitin-ligase enzyme,E3)的HECT結構域家族成員。HPV E6蛋白與E6AP結合后將活化的泛素轉移到p53蛋白上,最終p53蛋白在26S蛋白酶體的作用下沿泛素化降解途徑降解。CSN有8個亞單位,CSN1、2、3、4、7及8的C端均具有高度保守的PCI域(COP9蛋白酶體啟動因子),該結構有助于CSN各亞基間的相互結合以共同構建CSN復合體;而CSN5和CSN6的N端均具有高度保守的MPN域(Mpr1p-Pad1p的N端),該結構域對于CSN發揮促進腫瘤細胞增殖的作用非常重要[25-28]。 早期關于擬南芥的遺傳學研究[26]表明,CSN6的MPN結構域對于構建CSN必不可少。本研究結果顯示,在HPV陽性細胞系中,CSN6的過表達增加了E6AP蛋白的表達水平;而且shCSN6組p53蛋白表達水平較對照組明顯增加,這也對應了動物實驗中shCSN6組裸鼠的腫瘤生長明顯較對照組緩慢的現象。
本研究發現,在宮頸癌中CSN6對E6AP有正向調節作用,從而降低p53的穩定性,這與宮頸癌的發生發展密切相關。有關CSN6的作用機制值得進一步研究,例如前面提到的CSN6與smad、STAT等的相關性。
參考文獻
[1]Parkin DM.The global health burden of infection-associated cancers in the year 2002[J].Int J Cancer,2006,118(12):3030-3044.
[2]zur Hausen H.Papillomaviruses and cancer:from basic studies to clinical application[J].Nat Rev Cancer,2002,2(5):342-350.
[3]Bosch FX,de Sanjosé S.Chapter 1:Human papillomavirus and cervical cancer--burden and assessment of causality[J].J Natl Cancer Inst Monogr,2003,(31):3-13.
[5]Bernard HU,Burk RD,Chen Z,et al.Classification of papillomaviruses(PVs) based on 189 PV types and proposal of taxonomic amendments[J].Virology,2010,401(1):70-79.
[6]Scheffner M,Huibregtse JM,Vierstra RD,et al.The HPV-16 E6 and E6-AP complex functions as a ubiquitin-protein ligase in the ubiquitination of p53[J].Cell,1993,75(3):495-505.
[7]Wei N,Chamovitz DA,Deng XW.Arabidopsis COP9 is a component of a novel signaling complex mediating light control of development[J].Cell,1994,78(1):117-124.
[8]Li L,Deng XW.The COP9 signalosome:an alternative lid for the 26S proteasome?[J].Trends Cell Biol,2003,13(10): 507-509.
[9]Wei N,Deng XW.The COP9 signalosome[J].Annu Rev Cell Dev Biol,2003,19:261-286.
[10]Lee MH,Zhao R, Phan L,et al.Roles of COP9 signalosome in cancer[J].Cell Cycle,2011,10(18): 3057-3066.
[11]Duan W,Gao L,Wu X,et al.Differential response between the p53 ubiquitin-protein ligases Pirh2 and MdM2 following DNA damage in human cancer cells[J].Exp Cell Res,2006,312(17):3370-3378.
[12]Hetfeld BK,Peth A,Sun XM,et al.The COP9 signalosome-mediated deneddylation is stimulated by caspases during apoptosis[J].Apoptosis,2008,13(2):187-195.
[13]Richardson KS,Zundel W.The emerging role of the COP9 signalosome in cancer[J].Mol Cancer Res,2005,3(12): 645-653.
[14]Wang W,Tang M,Zhang L,et al.Clinical implications of CSN6 protein expression and correlation with mutant-type P53 protein in breast cancer[J].Jpn J Clin Oncol,2013,43(12):1170-1176.
[15]Xue Y,Chen J,Choi HH,et al.HER2-Akt signaling in regulating COP9 signalsome subunit 6 and p53[J].Cell Cycle,2012,11(22):4181-4190.
[16]Iyer SV, Iwakuma T. A novel link between the HER2-Akt and MDM2-p53 pathways via CSN6[J].Cell Cycle,2012,11(22): 4112.
[17]Birol M,Echalier A.Structure and function of MPN(Mpr1/Pad1 N-terminal) domain-containing proteins[J].Curr Protein Pept Sci,2014,15(5):504-517.
[18]Zhao R,Yeung SC,Chen J,et al.Subunit 6 of the COP9 signalosome promotes tumorigenesis in mice through stabilization of MDM2 and is upregulated in human cancers[J].J Clin Invest,2011,121(3):851-865.
[19]Hengstermann A,Linares LK,Ciechanover A,et al.Complete switch from Mdm2 to human papillomavirus E6-mediated degradation of p53 in cervical cancer cells[J].Proc Natl Acad Sci U S A,2001,98(3):1218-1223.
[20]Chen B,Zhao R,Su CH,et al.CDK inhibitor p57(Kip2) is negatively regulated by COP9 signalosome subunit 6[J].Cell Cycle,2012,11(24): 4633-4641.
[21]Choi HH,Gully C,Su CH,et al.COP9 signalosome subunit 6 stabilizes COP1,which functions as an E3 ubiquitin ligase for 14-3-3σ[J].Oncogene,2011,30(48):4791-4801.
[22]Vogt M,Butz K,Dymalla S,et al.Inhibition of Bax activity is crucial for the antiapoptotic function of the human papillomavirus E6 oncoprotein[J].Oncogene,2006,25(29):4009-4015.
[23]Lee MH,Lozano G.Regulation of the p53-MDM2 pathway by 14-3-3 sigma and other proteins[J].Semin Cancer Biol,2006,16(3):225-234.
[24]Yang H,Wen YY,Zhao R,et al.DNA damage-induced protein 14-3-3 sigma inhibits protein kinase B/Akt activation and suppresses Akt-activated cancer[J].Cancer Res,2006,66(6):3096-3105.
[25]Sharon M,Mao H,Boeri Erba E,et al.Symmetrical modularity of the COP9 signalosome complex suggests its multifunctionality[J].Structure,2009,17(1):31-40.
[26]Kotiguda GG,Weinberg D,Dessau M,et al.The organization of a CSN5-containing subcomplex of the COP9 signalosome[J].J Biol Chem,2012,287(50):42031-42041.
[27]Gusmaroli G,Figueroa P,Serino G,et al.Role of the MPN subunits in COP9 signalosome assembly and activity,and their regulatory interaction with Arabidopsis Cullin3-based E3 ligases[J].Plant Cell,2007,19(2):564-581.
[28]Zhang H,Gao ZQ,Wang WJ,et al.The crystal structure of the MPN domain from the COP9 signalosome subunit CSN6[J].FEBS Lett,2012,586(8):1147-1153.
Regulatory Effect of Constitutive Photomorphogenesis 9 Signalosome Subunit 6 on E6AP in Cervical Cancer and Its Clinical Significance
WANGChaoSONGYu*GUOQisang*LIUYanmei*GAOShujun*DepartmentofGynecology,*MedicalCenterofDiagnosisandTreatmentforCervicalDiseases,ObstetricsandGynecologyHospitalofFudanUniversity,Shanghai200011,China
AbstractObjective:To explore the role of constitutive photomorphogenesis 9 signalosome subunit 6(CSN6) in the regulation of E6AP in cervical cancer. Methods:CSN6 and E6AP genes were retrieved from The Cancer Genome Atlas(TCGA), and the correlation between them was analyzed. The effect of CSN6 on expression of E6AP and cytobiology was determined with transfection of plasmids and Western blotting,methyl thiazolyl tetrazolium(MTT) and flow cytometry(FCM).Nude mice were subcutaneously injected with Hela cell strains with different expression levels of CSN6, so as to set up the animal model. And the effect of CSN6 on growth of tumors was observed. Results: The database showed the overexpression and the mutation of CSN6 and E6AP in cancer. CSN6 could up-regulate the expression of E6AP. The inhibition on CSN6 could decrease the expression level of E6AP and the proliferation of cells, and induce the G0-G1 arrest.Animal expreiment showed that knock-out of CSN6 significantly inhibited the tumor growth, decreasing E6AP level and increasing p53 level. Conclusions: CSN6 could contribute to the occurrence and progression of cervical cancer by up-regulating the expression of E6AP.
Key WordsCervical cancer;Human papilloma virus;Infection;Constitutive photomorphogenesis 9 signalosome;E6AP
通訊作者高蜀君,E-mail:shujun99@163.com
基金項目:上海市科學技術委員會自然科學基金資助項目(編號:12ZR1403800)
中圖分類號R 737.33
文獻標識碼A
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
