重度冷應激對三河牛血液生化指標及相關基因表達的影響
2015-03-22 07:36:40李金龍吳宏軍劉愛榮王雅春
畜牧獸醫學報 2015年8期
關鍵詞:差異
李 瑋,劉 蕤,馬 堯,李金龍,吳宏軍,劉愛榮,俞 英,徐 青,王雅春*
(1.中國農業大學動物科技學院,農業部動物遺傳育種試驗室,畜禽育種國家工程實驗室,北京100193; 2.北京交通大學生命科學與生物工程研究院,北京 100044;3.寧夏大學農學院,銀川 750021; 4.內蒙古謝爾塔拉種牛場,海拉爾 021008;5.內蒙古海拉爾農墾集團,海拉爾 021008)
重度冷應激對三河牛血液生化指標及相關基因表達的影響
李 瑋1,3,劉 蕤1,馬 堯2,李金龍2,吳宏軍4,劉愛榮5,俞 英1,徐 青2,王雅春1*
(1.中國農業大學動物科技學院,農業部動物遺傳育種試驗室,畜禽育種國家工程實驗室,北京100193; 2.北京交通大學生命科學與生物工程研究院,北京 100044;3.寧夏大學農學院,銀川 750021; 4.內蒙古謝爾塔拉種牛場,海拉爾 021008;5.內蒙古海拉爾農墾集團,海拉爾 021008)
本研究旨在檢測與牛冷應激反應相關的生化指標,篩選參與牛冷應激反應的相關基因。以內蒙古三河牛為研究對象,對30頭青年牛進行室外-32 ℃冷應激處理3 h,采集冷應激前后試驗個體血樣,檢測血液生化指標及基因表達量的變化。冷應激3 h后,試驗個體T3、T4及ACTH3個血液生化指標極顯著升高(P<0.01)。mRNA差異顯示方法(DDRT)檢測到基因LIN54和KLF12在冷應激后表達量降低,熒光定量RT-PCR驗證了冷應激后2個基因表達下調(LIN54下調顯著(P<0.05),KLF12下調極顯著(P<0.01))的結果。篩選到的2個基因分別屬于LINC和KLF家族,LINC參與有絲分裂基因的活化,KLF參與細胞增殖和凋亡。T3、T4及ACTH 3項生化指標可以初步作為衡量三河牛冷應激反應強弱的侯選指標在大群體中進行驗證,篩選到的2個基因,可作為參與三河牛冷應激反應的重要侯選基因,進一步研究2個基因在牛冷應激反應中的作用機制,綜合這些生化指標及侯選基因表達的變化,用于評價個體冷應激反應的強度,在三河牛抗冷應激育種中將具有更廣泛的應用價值。
冷應激;三河牛;生化指標;基因表達;mRNA差異顯示
應激(Stress)是指機體對外界或內部的各種異常刺激所產生的非特異性應答反應的總和。溫度應激是應激反應中最常見的一種,包括熱應激和冷應激。冷應激是我國北方高寒地區一種最普遍的應激因素,在這些高寒地區,冬春寒冷季節達5個月之久,寒流常伴有暴風雪天氣。由于低溫而導致的冷應激常引起畜禽機體生理平衡的破壞,導致免疫力下降,表現為生長放緩、生產性能下降、繁殖力降低、發病率和死亡率高等。北方地區是我國畜禽養殖的主要地區,因此冷應激給我國北方地區的畜牧業造成了巨大經濟損失,是制約我國畜牧業高效發展的主要因素之一。
在冷應激狀態下,動物機體啟動一系列非特異性的自身免疫反應來中和應激的影響,這個過程會導致血液中一些生化分子的濃度發生變化。近年來,對小鼠的冷應激相關研究已證明,冷應激可導致動物促腎上腺皮質激素(ACTH) 的分泌增加,從而作用于腎上腺,調節皮質醇的合成和分泌,以使機體增強適應性,保持體內平衡[1]。另一方面,冷應激可導致動物較短時間內血液中促甲狀腺激素(TSH)的分泌增加,從而引起血清中三碘甲腺原氨酸(T3)、甲狀腺素(T4)水平升高,而不同動物冷應激后不同時間內T3和T4的變化不同[2]。血漿皮質酮(CORT)的濃度在動物急性冷應激時分泌增加,使動物可以抵御外界低溫環境的刺激。K.Fukuhara等[3]報道,大鼠在冷應激1 h后血漿皮質酮水平增加,9 h后趨向于正常,24 h時仍高于對照水平,而冷應激5 d時只稍高于對照水平。開產前金定蛋鴨冷應激12 h內,CORT的含量前期上升迅速,后期變化規律不明顯[4]。此外,動物冷應激反應中谷胱甘肽過氧化物酶(GSH-Px)與紅細胞鉀含量的變化也有較多報道[5-7]。
冷應激反應是生物機體的一種整體變化,涉及到機體多系統、多層次的協調效應[8-9]。關于動物冷應激反應機制及受冷應激調控的基因有很多報道,大約有幾十種基因參與機體的冷應激反應。其中研究較多的有熱休克蛋白基因(HSPs)、解偶聯蛋白基因(UCPs)、冷誘導結合RNA蛋白基因(CIRP)、金屬硫蛋白基因(MT)。李忠秋等[10]在對豬成纖維細胞進行的冷應激研究中發現,冷應激會誘導HSP90的表達量顯著上調,且隨著溫度的降低而升高。李士澤等[11]、吳永魁等[12]分別對大鼠和仔豬進行冷應激,發現HSP70蛋白表達量在組織及淋巴細胞中均上調。金福厚等[13]從冷處理的BALB/C鼠的睪丸組織中克隆出CIRP的cDNA,表明CIRP在生物體中能夠被低溫誘導,可能防止生物體遭受冷損傷。谷愛梅等[14]通過對NIH3T3細胞轉染MTⅡA基因,檢測到MT表達的上調可提高細胞對低溫的耐受性,且隨其含量的升高耐受性增大。這些基因的表達變化和冷應激強度及冷應激過程密切相關,可作為評價動物冷應激反應強弱的另一種有效的標志物。目前,篩選差異表達基因的技術主要有扣除雜交、抑制消減雜交、mRNA差異顯示等多種[15-16]。 其中,mRNA差異顯示技術[17](Differential display reverse transcription polymerase chain reaction,DDRT-PCR)具有速度快、操作簡便,能夠檢測低豐度mRNA、靈敏度高,花費較低等優點[18-20]。
目前,國內外有關畜禽冷應激的研究較少,而對牛冷應激相關的生化指標和基因的篩選研究的相關報道更少。本研究以內蒙古三河牛為試驗群體,-32 ℃冷應激3 h,檢測試驗個體應激前后血液生化指標及基因的表達變化,篩選三河牛冷應激反應相關的血液生化指標及差異表達基因,初步探討三河牛冷應激反應的分子機理。
1 材料與方法
1.1 試驗動物和樣品采集
隨機選取內蒙古謝爾塔拉牛場三河牛群體中體質健康、體重相近、月齡相近且無親緣關系的30頭青年母牛,飼養于同一牛舍內。采用配對試驗設計,進行冷應激前后三河牛血液生化指標和基因差異表達的試驗。該牛場為磚瓦結構,牛群采用半舍飼飼養方式,飼喂全混合日糧。試驗動物在溫度為5 ℃左右的牛舍內過夜飼養15 h,然后在室外平均氣溫-32 ℃的低溫環境中冷處理3 h。分別采集冷應激前后牛頸靜脈抗凝血10 mL,非抗凝血10 mL。
1.2 血液生化指標檢測
取所有30頭青年母牛的新鮮非抗凝血各10 mL,3 000 r·min-1離心10 min,收集血清檢測GSH-Px、T3、T4、ACTH、CORT的含量,收集紅細胞檢測鉀離子含量。所有生化指標均委托北京華英生物技術研究所采用放射免疫分析法進行測定。
1.3 血液白細胞總RNA提取
取30頭青年母牛的新鮮抗凝血各10 mL, 3 000 r·min-1離心10 min,吸取白膜層細胞,加入1.5 mL RNA固定液中,-80 ℃低溫冷凍保存,用于RNA的提取。采用TRIzol Reagent(Invitrogen)提取總RNA,取200 μL加入RNA固定液的白細胞樣品,加入1 mL TRIzol,充分裂解細胞,提取細胞總RNA,Nanodrop 2000檢測總RNA的含量和純度,2%瓊脂糖檢測總RNA完整性。
1.4 反轉錄和DDRT-PCR
在30頭青年母牛中任選6個個體樣本用于DDRT-PCR試驗,200 ng總RNA,加入2 μL 50 μmol·L-1錨定引物,用RNase-free水調至14 μL,70 ℃保溫10 min后,迅速置于冰上冷卻。加入4 μL 5×M-MLV Buffer,1 μL 10 mmol·L-1dNTP mixture,0.5 μL 40 U·μL-1RNase inhibitor,0.5 μL 200 U·μL-1M-MLV反轉錄酶(TaKaRa),42 ℃保溫1 h,70 ℃處理15 min,終止反應。以cDNA產物為模板,3條錨定引物和24條隨機引物(表3)兩兩組合進行DDRT-PCR 反應。反應體系為20 μL,其中2 μL 10×PCR Buffer,1.5 μL dNTP mixture(2.5 mmol·L-1),0.2 μLTaq聚合酶(5 U·μL-1),錨定引物(10 μmol·L-1)1 μL,隨機引物(10 μmol·L-1)0.5 μL,滅菌去離子水補齊至20 μL。反應條件: 94 ℃ 5 min;94 ℃ 30 s,38 ℃ 60 s,72 ℃ 60 s,25個循環;94 ℃ 30 s,40 ℃ 60 s,72 ℃ 60 s,15個循環;72 ℃ 5 min。3條錨定引物和24條隨機引物的序列詳見表3。
1.5 差異片段的PAGE凝膠電泳,銀染顯示及克隆測序
取DDRT-PCR產物4 μL,加入2 μL上樣緩沖液,6%非變性聚丙烯酰胺凝膠130 V電泳12~14 h,至溴酚藍離膠底1.5 cm時結束電泳,進行銀染。去離子水中漂洗5 min,10%乙醇固定10 min,1% AgNO3染色10 min,2.5% NaOH洗5 min,甲醛顯色至條帶清晰。記錄分子量為100~1 000 bp,清晰可辨,可以重復擴增的差異顯示片段。按同一擴增片段在試驗個體冷應激前后出現與否進行差異表達片段的篩選。篩選標準:在3個以上個體中變化同時出現,即作為差異表達的候選片段。回收差異表達的候選片段,采用與原PCR相同的引物組合和反應條件,進行二次擴增,克隆質粒酶切鑒定正確后,委托華大基因公司進行測序,用BLAST軟件進行序列分析。
1.6 熒光定量RT-PCR
對通過DDRT-PCR篩選到的候選差異基因,采用熒光定量RT-PCR進一步在30個個體中進行驗證。使用PrimerScript RT regent Kit(TaKaRa)合成cDNA第一鏈,凍存于-40 ℃。以Ensemble上牛的基因組序列為模板,采用Primer 3設計定量PCR引物(表1)。用SYBR Select Master Mix(Applied Biosystems)進行實時定量PCR反應,每個個體設置3次重復,以GAPDH為內參基因,通過Ct法計算該個體目的基因冷應激前后目標基因的相對表達量。
1.7 數據處理
冷應激前后血液中6項生化指標的數據,運用SAS9.1軟件進行配對t檢驗,再利用GLM過程分析各因素對生化指標變化的影響,模型中考慮體重和月齡;熒光定量RT-PCR數據運用Excel進行配對t檢驗。所有數值用“Mean±SD”表示。
2 結 果
2.1 冷應激前后三河牛血清生化指標的變化分析
對30頭三河牛冷應激前后的6項血液生化指標:血清谷胱甘肽過氧化物酶(GSH-Px)、三碘甲腺原氨酸(T3)、甲狀腺素(T4)、促腎上腺皮質激素(ACTH)、皮質酮(CORT)、紅細胞鉀含量進行檢測,統計分析結果見表2。三河牛冷應激3 h后,檢測的6項血液生化指標中,T3、T4和ACTH含量極顯著升高(P<0.01),其余幾項指標未表現出顯著差異,而試驗個體體重和月齡對各生化指標的變化無顯著影響(P>0.05)。
表1 熒光定量RT-PCR引物
Table 1 Primers for Real-time PCR

基因名稱Genename引物序列(5′?3′)PrimersequenceKLF12Forward:GCAAAGCACAAATGGACCCC,Reverse:AGGGCCGTAGATCCAGTTTCLIN54Forward:CTCTGGTCCGGTAATCACGA,Reverse:GAACCCTACCTGCAATGACCGAPDHForward:GGCGCCAAGAGGGTCAT,Reverse:AGGCATTGCTGACAATCTTGAG
表2 三河牛冷應激前后血液中相關生化指標的變化
Table 2 Changes of the blood biochemical parameters of Sanhe cattle before and after cold stress

T3/(ng·mL-1)T4/(ng·mL-1)ACTH/(pg·mL-1)CORT/(ng·mL-1)GSH?PX/(U·mL-1)K+/(mmol·L-1)應激前Beforecoldstress0.84±0.17a38.93±7.29a19.88±4.70a248.69±14.69801.11±50.9419.39±4.28應激后Aftercoldstress1.34±0.32b50.03±8.03b27.74±6.63b248.76±21.5784.21±107.4919.68±4.75
同列中不同肩注字母表示差異極顯著(P<0.01)
Different letters in the same row mean extremely significant difference between the treatments (P<0.01)
2.2 DDRT-PCR結果
本研究利用文獻[21]中報道的3條錨定引物和24條隨機引物配對組合,采用高低嚴謹(先經過25個低嚴謹PCR循環,再采用15個高嚴謹PCR循環)相結合的PCR方法,將大小為100~1 000 bp,清晰可見,相同的差異在3個以上個體中重現的片段記錄為差異片段,結果共發現21條冷應激前后表達量有差異的片段,表3提供了獲得21個差異片段的引物組合、大小等詳細信息,部分差異顯示結果見圖1 A。
將21條差異表達片段經過二次PCR擴增、回收、克隆后,最后得到11個大小正確的陽性克隆質粒,對11個克隆測序結果進行同源性比對,共發現2條序列分別與牛的2個基因同源,引物組合H-T11A/AP5所得差異片段位于牛基因組6號染色體LIN54基因的第一個外顯子(ENSBTAE00000113827)區,而H-T11A/AP12所得差異片段位于牛基因組12號染色體KLF12基因的第一個外顯子(ENSBTAE00000440535)區。Blast比對結果表明,這2個基因在人、小鼠、牛及其他一些物種之間具有較高的保守性(表4)。
2.3 差異基因的熒光定量PCR驗證結果
采用SYBR Green熒光標記的方法,以GAPDH作為內參基因,對2個差異表達基因LIN54和KLF12進行驗證。在mRNA差異顯示的銀染膠圖譜中(圖1A),與冷應激前相比,2個基因冷應激后表達下調。從基因的定量PCR結果可以看出(圖1B),2個差異顯示基因的相對表達量冷應激后顯著下調,這與mRNA差異顯示的銀染圖譜結果一致,其中KLF12極顯著下調(P<0.01),LIN54顯著下調(P<0.05)。
3 討 論
當機體遇到冷環境的刺激時,機體內T3和T4的含量迅速升高,T3的升高可增強Na+/K+-ATPase的活性,促進Na+、K+的跨膜轉運,引起紅細胞內K+含量的改變,并導致體內ATP消耗的增加,最終導致機體產熱增加。冷應激會激活下丘腦-垂體-甲狀腺軸(HTP),導致促甲狀腺激素釋放激素的分泌增加,從而引起血液中甲狀腺素(T4)、三碘甲腺原氨酸(T3)含量的升高。在本研究中,三河牛冷應激前后血清中T3、T4和ACTH的變化顯著,這與已有的研究報道一致,提示這3項生化指標可以作為衡量三河牛冷應激反應強弱的輔助指標。本研究沒有發現其他指標的顯著變化,可能是由于冷處理時間較短,或者三河牛對低溫有較強的耐受性[22-24]等原因造成的。因為不同種屬動物對冷應激的反應不盡相同,甚至同一種屬對不同時間和不同強度的冷應激所做出的反應也不同。對小鼠和豬進行不同程度的急性冷應激后,血液中ACTH、T3、T4的含量顯著升高,但延長冷應激時間或停止冷應激后,T3和T4的含量都會有下降的趨勢[25-26]。因此,利用血液中生化指標的變化來衡量三河牛冷應激反應的強弱,還需要更為詳盡的研究,而利用分子生物學手段,篩選與三河牛冷應激相關的分子標記,再綜合生化指標變化的數據,可能會更準確地反應個體對冷應激的反應,在三河牛抗冷應激育種中具有更廣泛的應用價值。
表3 DDRT-PCR引物及冷應激前后差異表達片段
Table 3 Primers for DDRT-PCR and fragments expressed differentially before and after cold stress
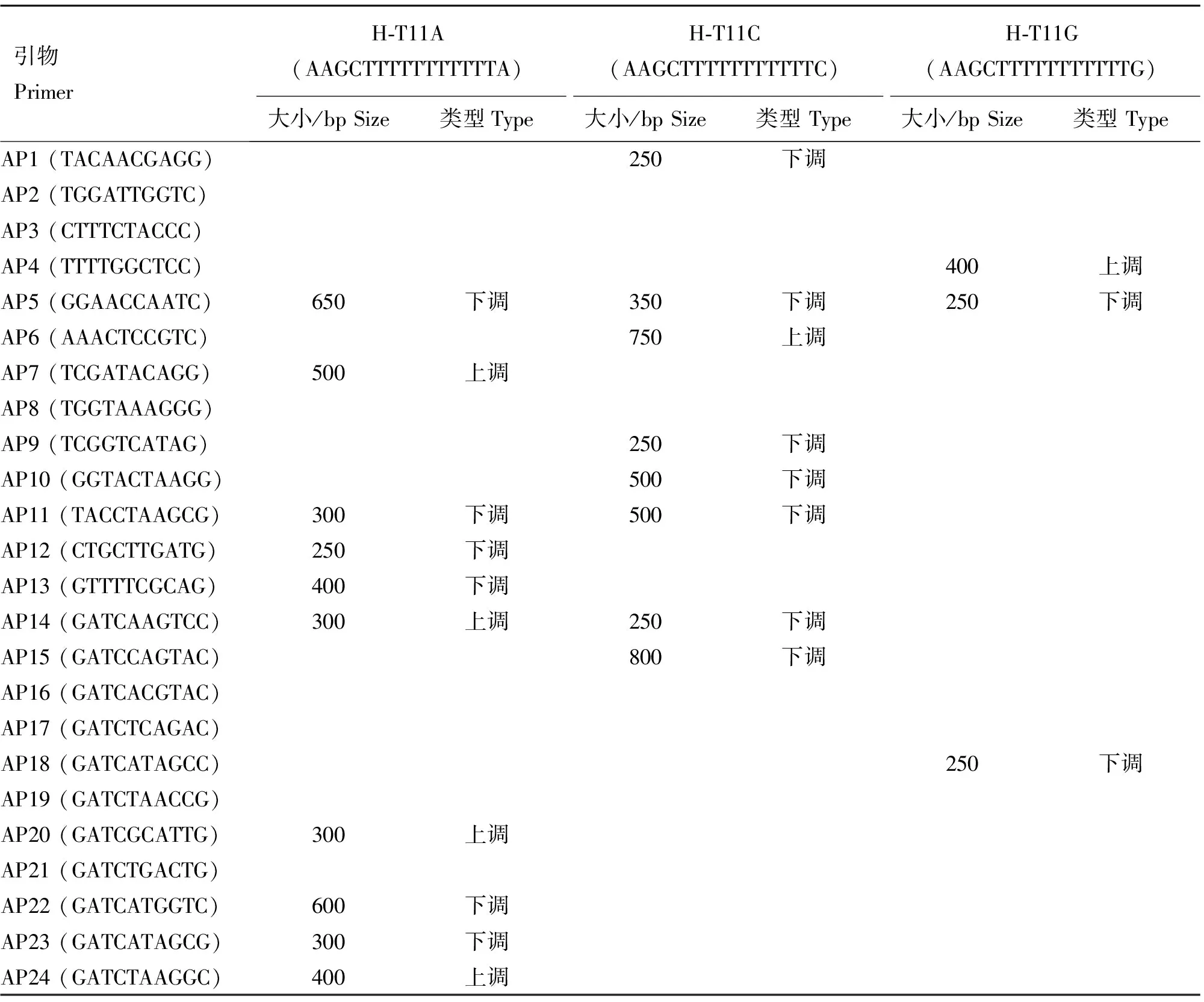
引物PrimerH?T11A(AAGCTTTTTTTTTTTA)H?T11C(AAGCTTTTTTTTTTTC)H?T11G(AAGCTTTTTTTTTTTG)大小/bpSize類型Type大小/bpSize類型Type大小/bpSize類型TypeAP1(TACAACGAGG)250下調AP2(TGGATTGGTC)AP3(CTTTCTACCC)AP4(TTTTGGCTCC)400上調AP5(GGAACCAATC)650下調350下調250下調AP6(AAACTCCGTC)750上調AP7(TCGATACAGG)500上調AP8(TGGTAAAGGG)AP9(TCGGTCATAG)250下調AP10(GGTACTAAGG)500下調AP11(TACCTAAGCG)300下調500下調AP12(CTGCTTGATG)250下調AP13(GTTTTCGCAG)400下調AP14(GATCAAGTCC)300上調250下調AP15(GATCCAGTAC)800下調AP16(GATCACGTAC)AP17(GATCTCAGAC)AP18(GATCATAGCC)250下調AP19(GATCTAACCG)AP20(GATCGCATTG)300上調AP21(GATCTGACTG)AP22(GATCATGGTC)600下調AP23(GATCATAGCG)300下調AP24(GATCTAAGGC)400上調
表4 差異表達片段Blast分析結果
Table 4 Blast analysis of differentially expressed fragments

差異片段Number同源基因Homologousgene片段大小/bpSize序列號AccessionNo.基因功能Genefunction表達模式Type1Proteinlin?54homolog(LIN54)510AC_000163.1有絲分裂相關基因的活化下調2Krueppel?likefactor12(KLF12)375AC_000169.1細胞增殖和凋亡下調
通過DDRT法篩選到2個差異表達基因,KLF12屬于Kruppel樣轉錄因子(Kruppel-like factors,KLF)家族。KLF家族是高度保守的轉錄因子家族,在真核細胞的基因轉錄調控過程中起著重要作用,參與調節細胞增殖、凋亡等多種生命活動。目前,KLF12與腫瘤相關的研究比較多,而與應激相關的研究未見報道。但體內和體外試驗證明,同一家族的KLF11基因能使氧化應激基因SOD2和CAT1的表達下調,增加了細胞對氧化應激的敏感性[27]。而冷應激可以誘發機體的氧化應激,導致體內自由基增多,對機體造成損傷。也有研究報道稱KLF11在小鼠肝臟的糖代謝過程中起著重要作用[28],而糖代謝又是為機體提供能量的主要途徑,與冷應激后機體維持體溫恒定密切相關,但具體的機理仍待進一步的研究。而同家族的另一成員KLF4,能夠與HSPs基因啟動子區結合可直接調控HSPs的基礎表達,在小鼠熱休克反應中表達上調[29],提示KLF12可能參與動物的應激反應。而LIN54屬于LINC家族(人源的LINC家族也稱DREAM),LINC家族是在哺乳動物細胞中發現的一種新的E2F-口袋蛋白復合體,在靜止期細胞和有絲分裂基因的活化中起重要作用[30-32]。LIN54是LINC的一個重要的家族成員,在細胞周期進程中起著重要的作用,LIN54富含半胱氨酸的CXC結構域是連接cdc2啟動子的新型DNA結合區域,在cdc2啟動子處發現了LIN54的兩個結合位點,其中一個和細胞周期轉錄起始位點的同源區域重疊。在冷應激環境中,細胞活性降低,因為LIN54與細胞周期進程高度相關,推測其在維持細胞穩態中起重要作用。

A.兩個冷應激前后差異表達片段銀染結果:BC.冷應激前;AC.冷應激后。B.兩個冷應激前后差異表達片段的熒光定量PCR結果:**代表差異極顯著(P<0.01);*代表差異顯著(P<0.05)A.Silver staining map of 2 fragments differentially displayed before and after cold stress:BC.Before cold stress;AC.After cold stress.B.RT-qPCR results of 2 differentially expressed before and after cold stress:** means very significant difference(P<0.01);* means significant difference(P<0.05)圖1 兩個冷應激前后差異表達片段Fig.1 Two fragments differentially expressed before and after cold stress
本研究所用樣品來自內蒙古海拉爾地區的三河牛,牛本身作為一種大型的經濟動物,在試驗處理和樣本的采集過程中存在許多局限性。在對三河牛冷應激研究的過程中,發現試驗個體的體重、月齡、個體血樣采集中的反應等都可能對試驗結果造成影響,而且無法采集參與冷應激調控體系中的重要組織樣本,不能比較參與組織與血液應對冷應激反應的基因表達及各生化指標的差異,不能連續追蹤冷應激不同時間內基因表達及各生化指標的變化趨勢等問題,使得對牛等大型動物的冷應激研究存在許多不確定性,為了克服這些問題,本課題組下一步準備先采用模式動物(小鼠或大鼠)和細胞模型,精確控制所有試驗條件,檢測不同時間及不同強度的冷應激條件下基因的表達差異及生化指標的變化,篩選與冷應激相關的生化指標和基因,然后再對牛群中對應的冷應激相關基因進行分析和驗證。盡管存在諸多問題,本研究首次從分子水平及生化指標2個方面同時進行牛的冷應激研究,后期的工作將擴大群體進一步驗證及分析篩選到的差異表達基因及檢測到的生化指標,尋找能夠評價牛冷應激反應強度的標志物,研究牛冷應激反應的分子遺傳機制。
[1] 孫 燕,柳鋒霖,宋耿青,等.急性和慢性束縛應激對大鼠內臟敏感性和神經內分泌的影響[J].中華消化雜志,2006,26(1):38-41. SUN Y,LIU F L,SONG G Q,et al.Effects of acute and chronic restraint stress on visceral sensitivity and neuroendocrine response in rats[J].ChineseJournalofDigestion,2006,26(1):38-41.(in Chinese)
[2] 李士澤,楊玉英,楊煥民,等.急性冷暴露和冷習服對大鼠和雛雞某些內分泌活動影響的比較生理學研究[J].中國應用生理學雜志,2008,24(1):23-24. LI S Z,YANG Y Y,YANG H M,et al.Comparative physiology research of acute cold exposure and cold acclimation to the certain endocrine activity in rats and chicks[J].TheChineseJournalofAppliedPhysiology,2008,24(1):23-24.(in Chinese)
[3] FUKUHARA K,KVETNANSKY R,CIZZA G,et al.Interrelations between sympathoadrenal system and hypothalamo-pituitary-adrenocortical/thyroid systems in rats exposed to cold stress[J].JNeuroendocrinol,1996,8(7):533-541.
[4] 王建鑫,王 安.急性冷應激對金定鴨開產前內分泌活動的影響[J].東北農業大學學報, 2005,36(4):480-485. WANG J X,WANG A.Study on the effect of acute cold stress on incretion activity in pre-egged Jin-ding ducks in cage[J].JournalofNortheastAgriculturalUniversity,2005,36(4):480-485.(in Chinese)
[5] PAJOVIC S B,PEJIC S,STOJILJKOVIC V,et al.Alterations in hippocampal antioxidant enzyme activities and sympatho-adrenomedullary system of rats in response to different stress models[J].PhysiolRes,2006,55:453-460.
[7] 曹靖寶,于宏敏.紅細胞鉀作為中國荷斯坦牛耐寒性選擇遺傳標記物的可靠性研究[J].中國奶牛,1997 (6):26-28. CAO J B,YU H M.Reliability research about erythrocyte potassium as the selected genetic markers of Chinese Holstein cow cold resistance[J].ChinaDairyCattle,1997(6):26-28.(in Chinese)
[8] WANG J W,XU S W.Effects of cold stress on the messenger ribonucleic acid levels of corticotrophin-releasing hormone and thyrotropin-releasing hormone in hypothalami of broilers[J].PoultSci,2008,87(5):973-978.
[9] WANG J T,LI S,LI J L,et al.Effects of cold stress on the messenger ribonucleic acid levels of peroxisome proliferator-activated receptor-γ in spleen,thymus,and bursa of fabricius of chickens[J].PoultSci,2009,88(12):2549-2554.
[10] 李忠秋,劉春龍,馬 紅,等.冷應激對東北野豬成纖維細胞HSP90 mRNA 轉錄水平的影響[J].畜牧獸醫學報,2012,43(12):1978-1983. LI Z Q,LIU C L,MA H,et al.Influence of cold stress on transcription ofHSP90 in northeast wild boar fibroblasts[J].ActaVeterinariaetZootechnicaSinica,2012,43(12):1978-1983.(in Chinese)
[11] 李士澤,任寶波,楊煥民,等.不同強度冷應激對大鼠肌肉、脾臟和肝臟中 HSP70 表達的影響[J].應用與環境生物學報,2006,12(2):235-238. LI S Z,REN B B,YANG H M,et al.The effects of different intensity of cold stress on the expression of HSP70 in rat muscle,spleen and liver[J].ChineseJournalofApplied&EnvironmentalBiology,2006,12(2):235-238.(in Chinese)
[12] 吳永魁,計 紅,胡仲明,等.不同強度冷應激對仔豬應激激素和 HSP70 的影響[A].中國畜牧獸醫學會動物傳染病學分會第三屆豬病防控學術研討會[C],2008:871-872. WU Y K,JI H,HU Z M,et al.The effects of different intensity of cold stress on stress hormones and HSP70 in piglet[A].China Animal Husbandry and Veterinary Association Animal Epidemiology Branch of the Third Conference on Swine Disease Prevention and Control[C],2008:871-872.(in Chinese)
[13] 金福厚,龐 巖,李士澤,等.BALB/C 鼠睪丸組織中冷誘導 RNA 結合蛋白的 cDNA 克隆與序列分析[J].應用與環境生物學報,2009,15(1):87-90. JIN F H,PANG Y,LI S Z,et al.CDNA cloning and sequence analysis of cold-induced RNA binding protein in BALB/C mice testicular tissue[J].ChineseJournalofApplied&EnvironmentalBiology,2009,15(1):87-90.(in Chinese)
[14] 谷愛梅,董兆申,鄧欣珠.金屬硫蛋白與細胞耐寒力的關系[J].中華勞動衛生職業病雜志,1999,17(5):272-275. GU A M,DONG Z S,DENG X Z.Relationship between metallothionein and cold tolerance of cells[J].ChineseJournalofIndustrialHygieneandOccupationalDiseases,1999,17(5):272-275.(in Chinese)
[15] 李文雍,陳清軒.mRNA 差異顯示技術研究進展[J].農業生物技術學報,2004,12(5):597-601. LI W Y,CHEN Q X.Progress in the technique of mRNA differential display[J].JournalofAgriculturalBiotechnology,2004,12(5):597-601.(in Chinese)
[16] 安 健,汪 明,王黎霞,等.mRNA 差異顯示 PCR 的研究進展[J].北京農學院學報,2005,20(2):64-68. AN J,WANG M,WANG L X,et al.mRNA differential display and its application on drug-resistance of coccidia[J].JournalofBeijingAgriculturalCollege,2005,20(2):64-68.(in Chinese)
[17] HAAG E,RAMAN V.Effects of primer choice and source ofTaqDNA polymerase on the banding patterns of differential display RT-PCR[J].Biotechniques,1994,17(2):226-228.
[18] LIANG P,PARDEE A B.Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction[J].Science,1992,257(5072):967-971.
[19] STEINAU M,RAJEEVAN M S.RNA profiling in peripheral blood cells by fluorescent differential display PCR[M].MethodsMolBiol,2009:211-222.
[20] SHEN M M.Identification of differentially expressed genes in mouse development using differential display andinsituhybridization[J].Methods,2001,24(1):15-27.
[21] BAUER D,MUULLER H,REICH J,et al.Identification of differentially expressed mRNA species by an improved display technique (DDRT-PCR)[J].NucleicAcidsRes,1993,21(18):4272-4280.
[22] 盧振峰,劉莉莉,趙國麗,等.三河牛DGAT1 基因 K232A 位點與產奶性狀的關聯分析[J].中國奶牛,2014(3):13-16. LU Z F,LIU L L,ZHAO G L,et al.Association of K232A in DGAT1 gene with milk production traits in Sanhe cattle[J].ChinaDairyCattle,2014(3):13-16.(in Chinese)
[23] 馬秋萌,秦春華,吳宏軍,等.三河牛目標性狀邊際效益研究[J].中國農業科學,2014,47(7):1409-1416. MA Q M,QIN C H,WU H J,et al.Study on marginal profits of traits in the breeding goal in Sanhe cattle[J].ScientiaAgriculturaSinica,2014,47(7):1409-1416.(in Chinese)
[24] 吳宏軍,馬孝林,劉愛榮,等.內蒙古三河牛培育歷程及進展[J].中國牛業科學,2012,38(4):48-52. WU H J,MA X L,LIU A R,et al.Breeding history and current improvements of Sanhe cattle in Inner Mongolia[J].ChinaCattleScience,2012,38(4):48-52.(in Chinese)
[25] 吳永魁.仔豬冷應激反應中激素、HSP70 及其 mRNA 的動態分析[D].長春:吉林大學,2006. WU Y K.Dynamic analysis of hormones,HSP70 and HSP70 mRNA during piglets cold stress[D].Changchun:Jilin University,2006.(in Chinese)
[26] SASAKI F,WU P,ROUGEAU D,et al.Cytochemical studies of responses of corticotropes and thyrotropes to cold and novel environment stress[J].Endocrinology,1990,127(1):285-297.
[27] FERNANDEZ-ZAPICO M E,MLADEK A,ELLENRIEDER V,et al.An mSin3A interaction domain links the transcriptional activity ofKLF11 with its role in growth regulation[J].EMBOJ,2003,22(18):4748-4758.
[28] 章華兵.KLF11作為一個關鍵的轉錄因子調節小鼠肝臟脂代謝的分子機制研究[D].北京:北京協和醫學院,2012. ZHANG H B.Mouse KLF11 is a key mediator of hepatic lipid metabolism[D].Beijing:Peking Union Medical College,2012.(in Chinese)
[29] 劉 瑛.KLF4:一個熱休克反應和熱休克蛋白表達的新調控介質[D].長沙:中南大學,2006. LIU Y.KLF4,a new regulate mediator of heat shock response and heat shock protein[D].Changsha:Central South University,2006.(in Chinese)
[30] LITOVCHICK L,SADASIVAM S,FLORENS L,et al.Evolutionarily conserved multisubunit RBL2/p130 and E2F4 protein complex represses human cell cycle-dependent genes in quiescence[J].MolCell,2007,26(4):539-551.
[31] SCHMIT F,KORENJAK M,MANNEFELD M,et al.LINC,a human complex that is related to pRB-containing complexes in invertebrates regulates the expression ofG2/Mgenes[J].CellCycle,2007,6(15):1903-1913.
[32] PILKINTON M,SANDOVAL R,COLAMONICI O R.Mammalian Mip/LIN-9 interacts with either the p107,p130/E2F4 repressor complex or B-Myb in a cell cycle-phase-dependent context distinct from theDrosophiladREAM complex[J].Oncogene,2007,26(54):7535-7543.
(編輯 郭云雁)
Effects of Severe Cold Stress on Blood Biochemical Parameters and Related Gene Expression in Sanhe Cattle
LI Wei1,3,LIU Rui1,MA Yao2,LI Jin-long2,WU Hong-jun4,LIU Ai-rong5, YU Ying1,XU Qing2,WANG Ya-chun1*
(1.KeyLaboratoryofAgriculturalAnimalGeneticsandBreeding,NationalEngineeringLaboratoryofAnimalBreeding,CollegeofAnimalScienceandTechnology,ChinaAgriculturalUniversity,Beijing100193,China;2.InstituteofBiologicalScienceandBioengineering,BeijingJiaotongUniversity,Beijing100044,China;3.SchoolofAgriculture,NingxiaUniversity,Yinchuan750021,China;4.InnerMongoliaXieertalaCattleBreedingFarm,Hailaer021008,China; 5.InnerMongoliaHailaerAgriculturalReclamationGroup,Hailaer021008,China)
The aim of this study was to detect the biochemical indexes and relevant genes related to cold stress in Sanhe cattle.Thirty heifer Sanhe cattle in Inner Mongolia were kept at -32 ℃ for 3 h,and then blood samples were collected before and after cold stress separately to detect the biochemical indexes and gene expression changes.After 3 h cold stress,T3,T4 and ACTH in blood significantly increased (P<0.01).The expressions ofLIN54 andKLF12 decreased in silver staining map produced by mRNA differential display method (DDRT),decrease of the 2 genes expression were further verified at the level ofP<0.05 forLIN54 andP<0.01 forKLF12 by fluorescence quantitative RT-PCR.The 2 genes belonged to LINC and KLF families,respectively,LINC involved in mitotic gene activation,KLF involved in cell proliferation and apoptosis.T3,T4 and ACTH may be used as the candidate indexes to measure the strength of cold stress in large population,LIN54 andKLF12 may be the important candidate genes for further study on genetic mechanism of cold stress in cattle.The combination of these biochemical indexes and expression change of candidate genes would be used to assess the change of individual under cold stress,which will be more valuable in breeding for cold resistance ability in Sanhe cattle.
cold stress;Sanhe cattle;biochemical index;gene expression;mRNA differential display
10.11843/j.issn.0366-6964.2015.08.024
2014-11-07
國家自然科學基金(31172191);“十二五”科技支撐計劃(2011BAD28B02);現代農業產業技術體系建設專項資金(CARS-37);長江學者和創新團隊發展計劃(IRT1191)
李 瑋(1990-),女,河北邢臺人,碩士生,主要從事動物分子遺傳學研究,E-mail:lw9056@163.com;劉 蕤(1988-),女,四川宜賓人,碩士,主要從事動物分子數量遺傳學研究,E-mail:kkrabbit-2007@163.com。李 瑋和劉 蕤為共同第一作者
*通信作者:王雅春,教授,E-mail:wangyachun@cau.edu.cn
S823.2
A
0366-6964(2015)08-1463-08
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
