SG97A平衡致死系統的構建及其初步應用
2015-03-22 12:44:59耿士忠陳曉娟潘志明焦新安
畜牧獸醫學報 2015年12期
關鍵詞:系統
耿士忠,陳曉娟,潘志明,焦新安
(揚州大學 江蘇省人獸共患病學重點實驗室/江蘇省動物重要疫病與人獸共患病防控協同創新中心,揚州 225009)
SG97A平衡致死系統的構建及其初步應用
耿士忠#,陳曉娟#,潘志明,焦新安*
(揚州大學 江蘇省人獸共患病學重點實驗室/江蘇省動物重要疫病與人獸共患病防控協同創新中心,揚州 225009)
敲除雞傷寒沙門菌(SG)減毒疫苗株97Aasd基因,構建宿主載體平衡致死系統并表達新城疫病毒(NDV)F蛋白,構建二價活菌苗。PCR擴增兩段位于asd基因外側的序列作為等位基因,以氯霉素抗性(CmR)基因替代asd基因,構建自殺質粒pGMB151Δasd::CmR,通過E.coliχ7213與SG97A固相雜交,將質粒轉入SG97A,反向篩選獲得SG97AΔasd::CmR,利用質粒pCP20去除CmR后,DAP的依賴菌株命名為SG97AΔasd。將含asd基因的質粒pYA3334轉入SG97AΔasd中構建宿主載體平衡致死系統,并表達NDV-F蛋白。結果表明,通過等位基因重組方法,成功構建SG97AΔasd及宿主載體平衡致死系統,重組菌能夠表達NDV-F蛋白,并能在雞體內誘導產生特異性抗體。本研究成功構建SG97AΔasd宿主載體平衡致死系統,并表達NDV-F蛋白及其誘導宿主產生抗體,為應用該系統構建二價活菌苗的研究奠定了基礎。
雞傷寒沙門菌;asd基因; 宿主載體平衡致死系統;NDV-F蛋白
雞傷寒(fowl typhoid)是由雞傷寒沙門菌引起的雞的常見多發性傳染病,呈急性或慢性敗血癥狀,以肝、脾等實質器官的病變和下痢為特征,主要發生于成年雞和青年雞,產蛋期母雞對本病最為敏感,蛋雞一旦暴發雞傷寒,其死亡率急劇上升。該菌除了水平傳播外,也能經卵垂直傳遞,帶菌種雞所產種蛋的帶菌率可達20%~30%,不僅孵化率低,并有死雛病雛,還可污染孵化器、出雛器和育雛室,造成水平傳播擴散,使整個養雞周期呈惡性循環,給養雞業帶來的直接和間接損失巨大。減毒雞沙門菌97A是經轉座突變及粗糙變異方法篩選所獲得,研究結果表明該減毒株具有較好的安全性和免疫效力,也能顯著地限制強毒株在雞體內的定居和增殖[1]。
多價疫苗和黏膜免疫是未來免疫預防傳染病的主要方向。利用宿主載體平衡致死系統使沙門菌作為疫苗載體已在醫學與獸醫學領域引起廣泛重視,但目前研究沙門菌載體多為鼠傷寒沙門菌[2-3],其他沙門菌較為少見。作者以減毒雞沙門菌疫苗株97A為親本菌,敲除asd基因后,構建以減毒雞沙門菌疫苗株97AΔasd基因缺失株為宿主和含asd基因的pYA3334質粒為載體的宿主載體平衡致死系統,并利用該系統表達NDV-F蛋白,為構建二價活疫苗的研究奠定基礎。
1 材料與方法
1.1 菌株與質粒
試驗所用的細菌和質粒,見表1。
表1 試驗所用的細菌和質粒
Table 1 Bacteria and plasmids used in this study

材料Material名稱Name用途Use來源Source細菌BacteriaSG97ARecipientOurlabE.coliDH5αHostforpMD18OurlabE.coliχ7213(KmRΔasd)HostforpGMB151OurlabE.coliSpy372(λ?pir)HostforpGMB151Ourlabχ4550(S.typhimurium)STΔasdOurlabE.coliχ6216HostforpGMB151OurlabE.coliχ3730HostforpGMB151tomethylateOurlab質粒PlasmidpUC18(AmpR)CloningvectorOurlabpMD18?TCloningvectorTaKaRapKD3IncludingCmRgeneOurlabpGMB151(AmpRSmR)SuicidevectorOurlabpYA3334(asd+)PlasmidofexpressingDAPOurlabpYA3334?dsRED(asd+)PlasmidofexpressingDAPanddsREDOurlab
1.2 主要試劑
用于分子克隆的 Primer STAR HS聚合酶、限制性內切酶、連接酶、產物純化試劑盒和膠回收試劑盒均購自TaKaRa公司,羊抗雞HRP-IgG和羊抗鼠HRP-IgG購自Sigma公司,GST-F蛋白及其小鼠抗血清自備。
1.3 引物設計與合成
根據GenBank中已發表的沙門菌基因組序列,以軟件Primer5.0設計出特異性的引物,如表2所示。引物由南京金思瑞生物有限公司合成。
表2 PCR擴增所用的引物序列
Table 2 The primer sets for PCR in this study

目的基因Geneamplified引物名稱Primers引物序列(5′?3′)Primesequences擴增片段長度/bpAmpliconsize備注Noteasd12asdp1asdp2TTGGATCCCCGTTGAATGATGATGACCGTTCTCGAGTGCGTTAGGAAGGGAATC1959BamHⅠXhoⅠasd34asdp3asdp4TTCTCGAGGTAGCTTAATCCCGCGGGTATTGGATCCGAGCGTTCATTGTCATCGAC2079XhoⅠBamHⅠasd56asdp5asdp6TTGCTTTCCAACTGCTGAGCTCCTATCTGCGTCGTCCTAC1796(wt)1360(Δasd+CmR)CmRCmFCmRACTCGAGGTGTAGGCTGGAGCTGCTTCACTCGAGATGGGAATTAGCCATGGTCC1032XhoⅠXhoⅠHtohtoFhtoRACTGGCGTTATCCCTTTCTCTGCTGATGTTGTCCTGCCCCTGGTAAGAGA495SalmonellaNDV?FFp1Fp2AACCATGGGAAATGCTGCCAACATCCTCAAGGATCCATCCAAATCGGTCTCTAC594NcoⅠBamHⅠ
1.4 雞傷寒沙門菌減毒株SG97AΔasd缺失株的構建
根據本實驗室所建立的沙門菌基因敲除方法[4-5],通過擴增asd基因上下游1 959和2 079 bp兩個片段作為同源臂,以便等位基因交換。以CmR替代asd基因,構建重組自殺質粒,固相雜交法轉移重組自殺質粒到雞傷寒沙門菌減毒株97A中獲得單同源臂交叉互換重組菌后,運用10%蔗糖和無NaCl培養基及氯霉素篩選DAP生長依賴細菌,即可以獲得asd基因缺失株,命名為SG97AΔasd。
1.5 宿主載體平衡致死系統的構建
將含有asd基因的pYA3334質粒及含有紅色熒光蛋白基因(dsRED)的pYA3334-dsRED質粒轉化到χ4550和SG97AΔasd中,構建宿主載體平衡致死系統,觀察重組菌對DAP的生長依賴性及外源紅色熒光蛋白基因表達。
1.6 宿主載體平衡致死系統表達NDV-F基因
1.6.1 構建表達NDV-F基因宿主載體平衡致死系統 PCR擴增含NDV-F基因,克隆測序后,亞克隆到pYA3334(asd+)質粒限制性內切酶識別位點NcoⅠ和BamHⅠ間,形成重組質粒pYA3334-F。經宿主菌E.coliχ6212及中間宿主E.coliχ3730(甲基化重組質粒)后,再轉化至終末宿主減毒雞傷寒沙門菌SG97AΔasd缺失株及χ4550,形成重組菌SG97A(pYA3334-F)和χ4550(pYA3334-F)。1.6.2 重組質粒pYA3334-F在SG97AΔasd和χ4550中的表達 分別取OD600 nm=1.0的重組沙門菌χ4550(pYA3334-F)、SG97A(pYA3334-F)及空載體菌χ4550(pYA33340)和SG97AΔasd(pYA3334)各1 mL,12 000 r·min-1離心2 min,棄上清,沉淀用PBS洗滌3次后,加50 μL的ddH2O重懸,加等體積的2 ×上樣緩沖液,煮沸5 min,各取20 μL進行SDS-PAGE電泳;并以χ4550表達的F蛋白割膠免疫法制備小鼠抗血清作Western blotting分析。
1.6.3 重組菌免疫接種雛雞 以重組菌SG97AΔasd(pYA3334-F) 免疫接種10日齡雛雞,3組,10羽·組-1,1×109CFU·羽-1,肌肉注射免疫3次,中間間隔2周。各免疫組免疫后第12 天采血,制備血清,-20 ℃保存。同時設SG97AΔasd(pYA3334)陰性對照組和PBS空白對照組。
1.6.4 抗體效價的測定 GST-F蛋白經純化后包被ELISA板,20 μg·孔-1,4 ℃包被過夜,PBST洗滌3次;加入含10% BSA的封閉液,200 μL·孔-1,4 ℃封閉過夜后,PBST洗滌5次;加入各免疫組血清,同時設立陰性血清對照和PBS空白對照,37 ℃作用3 h后,PBST洗滌3次;加入HRP標記的羊抗雞IgG抗體(工作濃度1∶10 000),37 ℃作用1 h后,PBST洗滌4次;加入底物OPD,37 ℃避光顯色15 min,用2 mol·L-1H2SO4終止后測OD492 nm值。
2 結 果
2.1 雞傷寒沙門菌減毒株SG97AΔasd的構建
asdp1/p2擴增出約2 kb基因片段asd12,asdp3/p4擴增出約2 kb基因片段asd34,asdp5/p6能擴增出SG97AΔasd約1.4 kb基因片段,而野生菌(wt)則擴增出1.84 kb基因片段;CmF/CmR能擴增出1.0 kbCmR基因,親本株SG97A則不能被擴增(圖1)。結果顯示通過多對引物PCR鑒定,擴增產物都與預期的大小一致,表明雞傷寒沙門菌減毒株SG97AΔasd被成功構建,且通過特異性引物htoF/htoR PCR證實該菌為雞傷寒沙門菌(圖2)。

M.DL2000 DNA相對分子質量標準;1,3,5,7.SG97AΔasd;2,4,6,8.SG97A;1,2.asdP1/P2 PCR;3,4.asdP3/P4 PCR;5,6. asdP5/P6 PCR;7,8.CmF/CmR PCRM.DL2000 DNA marker;1,3,5,7.SG97AΔasd;2,4,6,8.SG97A;1,2.asdP1/P2 PCR;3,4.asdP3/P4 PCR;5,6.asdP5/P6 PCR;7,8.CmF/CmR PCR圖1 SG97AΔasd缺失株的PCR鑒定Fig.1 PCR identification of SG97AΔasd

M. DL2000 DNA相對分子質量標準; 1. 野生菌; 2、3. SG97AΔasd; 4. ddH2O對照M. DL2000 DNA marker; 1. wt; 2, 3. SG97AΔasd; 4. ddH2O control圖2 SG97A沙門菌屬特異性鑒定Fig.2 PCR identification of Salmonella
2.2 宿主載體平衡致死系統的構建
根據asd基因表達產物特性,SG97AΔasd對二氨基庚二酸(DAP)生長依賴,圖3中顯示SG97AΔasd在含DAP培養基上能生長(A),而在無DAP培養基上則不能生長(B)。同樣,轉化了質粒pYA3334的SG97AΔasd在無DAP的培養基上則能生長,而沒有轉化質粒pYA3334的SG97AΔasd則不能在無DAP的培養基上生長。兩個結果都說明SG97AΔasd對DAP具有生長依賴性。通過將能夠表達紅色熒光蛋白的pYA3334-dsRED質粒轉化入SG97AΔasd,互補細菌中能夠表達外源基因(A),顯現紅色熒光;而僅轉化pYA3334的質粒則無熒光出現(B)(圖4)。

圖3 SG97AΔasd在LB+DAP平板(A)和LB平板(B)上的生長情況Fig.3 The growth of SG97A Δasd on LB with DAP (A) or without DAP (B)
2.3 宿主載體平衡致死系統的生長特性及其穩定性
如圖5所示,轉化pYA3334質粒的SG97AΔasd缺失株形成載體宿主互補的平衡致死系統后,互補菌生長速度與SG97AΔasd缺失株相當,與野生菌的生長速度有顯著的差別。通過FACS測定轉化了pYA3334-dsRED質粒的SG97AΔasd能觀察紅色熒光蛋白的表達,第100代載體宿主互補菌與第50代和第1代的細菌差異不大(圖6),說明載體宿主之間能夠形成的平衡致死系統有良好的穩定性。
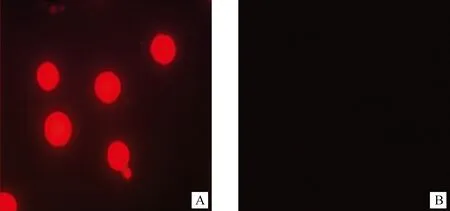
A. SG97AΔasd (pYA3334-dsRED);B. SG97AΔasd (pYA3334)圖4 SG97AΔasd互補細菌表達紅色熒光Fig.4 Red fluorescence expressed by SG97AΔasd(pYA3334-dsRED)

圖5 SG97AΔasd缺失株生長曲線Fig.5 The growth curves of SG97A and SG97AΔasd

圖6 FACS測定不同代次SG97A Δasd (pYA3334-dsRED)中dsRED的穩定性Fig.6 The stability of SG97A Δasd (pYA3334-dsRED) by FACS analysis
2.4 利用SG97AΔasd宿主載體平衡致死系統表達新城疫病毒F基因
2.4.1 表達質粒重組菌的構建 PCR擴增新城疫病毒(NDV)F基因,克隆測序正確后,亞克隆入pYA3334載體限制性內切酶識別位點NcoⅠ和BamHⅠ間,構建宿主平衡致死系統表達質粒pYA3334-F,經感受態E.coliχ6212擴增,及中間宿主E.coliχ3730完成重組質粒的甲基化過程后,重組質粒pYA3334-F轉化χ4550和SG97AΔasd,分別命名為χ4550(pYA3334-F)和SG97AΔasd(pYA3334-F)。
2.4.2 重組菌χ4550(pYA3334-F)和SG97AΔasd(pYA3334-F)的Western blotting鑒定 新城疫病毒F蛋白能夠在重組菌χ4550(pYA3334-F)中正確表達,約20 kd(圖7)。但是在重組菌SG97AΔasd(pYA3334-F)表達產物約40 ku,為理論值的2倍(圖8A),Western blotting分析顯示該表達產物能與小鼠F蛋白抗血清特異性反應(圖8B)。
2.4.3 重組沙門菌SG97AΔasd(pYA3334-F)誘導的F蛋白抗體測定結果 SG97AΔasd(pYA3334-F)以1×109CFU·羽-1的劑量肌注免疫雛雞,SG97AΔasd(pYA3334)作為陰性對照,分別在二免及三免后第12天采血,制備血清。間接ELISA結果顯示在雞二免后血清內檢測到抗NDV-F蛋白的抗體,平均抗體效價為1∶200;三免后平均抗體效價為1∶400(圖9)。

M.蛋白質相對分子質量標準; 1.χ4550 (pYA3334-F); 2. χ4550 (pYA3334)M.Protein marker;1.χ4550 (pYA3334-F);2.χ4550 (pYA3334)圖7 χ4550 (pYA3334-F)的SDS-PAGE鑒定Fig.7 SDS-PAGE identification of χ4550 (pYA3334-F)

M.蛋白質相對分子質量標準;1.SG97AΔasd (pYA3334-F);2.SG97AΔasd (pYA3334)M.Protein marker;1.SG97AΔasd (pYA3334-F);2.SG97AΔasd (pYA3334)圖8 SG97AΔasd (pYA3334-F)的SDS-PAGE(A)及Western blotting(B)鑒定Fig.8 SDS-PAGE(A) and Western blotting(B) identification of SG97AΔasd (pYA3334-F)

圖9 間接ELISA檢測雞血清中抗NDV-F抗體滴度Fig.9 The titers of anti-NDV-F antibody in chicken sera by indirect ELISA
3 討 論
多價苗由于免疫后同時獲得對多種病原的免疫保護,減少對動物的應激,省時省力而備受青睞。1988年,宿主載體平衡致死系統首次被報道應用于重組減毒沙門菌疫苗(recombinant attaunatedSalmonellavaccines,RASVs)。此系統能夠確保編碼外源基因的重組質粒在重組菌免疫宿主后的穩定性,避免抗生素耐藥基因的使用[6]。以減毒沙門菌作為載體構建雙價或多價活疫苗,是目前研制新型疫苗的重要途徑之一。所構建的重組減毒沙門菌可經口服免疫、鼻內免疫[7-8]和直腸免疫[9]等多種途徑免疫,在機體內表達特異抗原[10]誘導獲得體液免疫、細胞免疫及黏膜免疫[11-12],提供對細菌[13]、寄生蟲[14]和病毒[15]感染的保護作用。
當前減毒鼠傷寒沙門菌χ4550(Δasd)是宿主載體平衡致死系統研究較多的工程菌,有學者應用攜帶能編碼乙肝病毒核心抗原質粒的χ4550菌株經胃腸道免疫小鼠后,誘導出了較長久的黏膜保護性抗體[16-17],表明減毒鼠傷寒沙門菌χ4550作為口服疫苗載體具有應用價值。以其他血清型沙門菌為疫苗載體的研究也在研究嘗試中,如豬霍亂沙門菌[18]、雞白痢沙門菌[2]、遲鈍愛德華菌[19]。本研究則利用已有的減毒雞傷寒沙門菌97A,通過缺失其asd基因成功構建宿主載體平衡致死系統,并表達外源蛋白,誘導動物機體產生免疫。
在研究中,作者發現pYA3334-F 質粒在χ4550內表達F蛋白與理論值一樣,但在SG97AΔasd中體外表達蛋白質大小是理論值的2倍,Western blotting分析顯示其能夠與鼠源F蛋白抗血清特異反應,SG97A(pYA3334-F)以1×109CFU·羽-1的劑量肌注免疫雛雞,陽性血清二免平均抗F抗體效價為1∶200,三免平均抗F抗體效價為1∶400,該結果說明pYA3334-F在減毒雞傷寒沙門菌SG97AΔasd中表達了外源F蛋白,但2倍于理論值大小的原因不詳,有待進一步研究。
4 結 論
成功構建SG97AΔasd宿主載體平衡致死系統,能夠表達NDV-F蛋白并誘導宿主產生抗體,為應用該系統構建沙門菌二價活菌苗的研究奠定了基礎。
[1] 劉海俠,王寶安,潘志明,等.減毒雞沙門氏菌疫苗株97A安全性評價[J].中國獸醫雜志,2007,43(11):72-73. LIU H X,WANG B A,PAN Z M,et al.Evaluation on safety to Chicken of attenuatedSalmonellavaccine strain 97A[J].ChineseJournalofVeterinaryMedicine,2007,43(11):72-73.(in Chinese)
[2] 陳松彪,李 靜,尚 珂,等.雞白痢沙門菌C79-13 ΔcrpΔasd平衡致死系統的構建及其生物學特性的研究[J].中國免疫學雜志,2014,30(8):1083-1087,1092. CHEN S B,LI J,SHANG K,et al.Construction ofSalmonellaPullorum C79-13 ΔcrpΔasd mutant balanced-lethal host-vector system and its biological features[J].ChineseJournalofImmunology,2014,30(8):1083-1087,1092.(in Chinese)
[3] 徐引弟,郭愛珍,劉維紅,等.豬霍亂沙門菌C500 株ΔcrpΔasd缺失株平衡致死載體系統的構建及鑒定[J].生物工程學報,2006,22(3):366-372. XU Y D,GUO A Z,LIU W H,et al.Construction and characterization ofΔcrpΔasdMutant host-vector balanced lethal system ofSalmonellacholeraesuisC500 strain[J].ChineseJournalofBiotechnology,2006,22(3):366-372.(in Chinese)
[4] GENG S Z,JIAO X A,PAN Z M,et al.An improved method to knock out theasdgene ofSalmonellaentericaserovar Pullorum[J].JBiomedBiotechnol,2009,2009:646380.doi:10.1155/2009/646380.
[5] 耿士忠,劉男男,焦新安,等.雞白痢沙門菌S06004ΔspiC突變株的構建與鑒定[J].中國獸醫科學,2014,44(4):379-386. GENG S Z,LIU N N,JIAO X A.et al.Development and identification ofspiCgene-deletedSalmonellapullorummutant strainΔspiC[J].ChineseVeterinaryScience,2014,44(4):379-386.(in Chinese)
[6] WANG S F,KONG Q K,CURTISS R III.New technologies in developing recombinant attenuatedSalmonellavaccine vectors[J].MicrobPathog,2013,58:17-28.
[7] GALEN J E,GOMEZ-DUARTE O G,LOSONSKY G A,et al.A murine model of intranasal immunization to assess the immunogenicity of attenuatedSalmonellatyphilive vector vaccines in stimulating serum antibody responses to expressed foreign antigens[J].Vaccine,1997,15(6-7):700-708.
[8] VECINO W H,MORIN P M,AGHA R,et al.Mucosal DNA vaccination with highly attenuatedShigellais superior to attenuatedSalmonellaand comparable to intramuscular DNA vaccination for T cells against HIV[J].ImmunolLett,2002,82(3):197-204.
[9] KARPENKO L I,NEKRASOVA N A,IGNAT’EV G M,et al.Immune response in oral and rectal immunization by the attenuated strain ofSalmonellacarrying the HIV DNA-vaccine[J].VoprVirusol,2003,48(4):16-20.
[10] PASETTI M F,LEVINE M M,SZTEIN M B,et al.Animal models paving the way for clinical trials of attenuatedSalmonellaentericaserovar typhi live oral vaccines and live vectors[J].Vaccine,2003,21(5-6):401-418.
[11] CURTISS R Ⅲ,ZHANG X,WANDA S Y,et al.Induction of host immune responses usingSalmonella-vectored vaccines//Brogden K A,Minion F C,Cornick N(ed).Virulence mechanisms of bacterial pathogens[M].4th ed.ASM Press,Washington,DC.2007:297-313.
[12] 劉 婷,焦新安,潘志明,等.表達破傷風毒素C片段的重組鼠傷寒沙門菌的構建與鑒定[J].中國獸醫科技,2005,35(11):883-887. LIU T,JIAO X A,PAN Z M,et al.Construction and identification of recombinantSalmonellatyphimuriumstrain expressing tetanus toxin fragment C[J].ChineseJournalofVeterinaryScienceandTechnology,2005,35(11):883-887.(in Chinese)
[13] SCHRIEFER A,MALTEZ J R,SILVA N,et al.Expression of a pilin subunit BfpA of the bundle-forming pilus of enteropathogenicEscherichiacoliin anaroAliveSalmonellavaccine strain[J].Vaccine,1999,17(7-8):770-778.
[14] QU D,WANG S,CAI W,et al.Protective effect of a DNA vaccine delivered in attenuatedSalmonellatyphimuriumagainstToxoplasmagondiiinfection in mice[J].Vaccine,2008,26(35):4541-4548.
[15] WANG J,MICHEL V,LECLERC C,et al.Immunogenicity of viral B-cell epitopes inserted into two surface loops of theEscherichiacoliK12 LamB protein and expressed in an attenuatedaroAstrain ofSalmonellatyphimurium[J].Vaccine,1999,17(1):1-12.
[16] HOPKINS S,KRAEHENBUHL J P,SCH?DEL F,et al.A recombinantSalmonellatyphimuriumvaccine induces local immunity by four different routes of immunization[J].InfectImmun,1995,63(9):3279-3286.
[17] BACHTIAR E W,SHENG K C,FIFIS T,et al.Delivery of a heterologous antigen by a registeredSalmonellavaccine(STM1)[J].FEMSMicrobiolLett,2003,227(2):211-217.
[18] 徐引弟.豬霍亂沙門氏菌C500株crp-、asd-缺失株平衡致死系統的構建及應用[D].武漢:華中農業大學,2006. XU Y D.Construction and application ofcrp-,asd-deleted mutants host-vector balanced lethal system ofSalmonellacholeraesuisC500 strain[D].Wuhan:Huazhong Agricultural University,2006.(in Chinese)
[19] 閆一劍.遲鈍愛德華氏菌平衡致死系統的構建及其在新型疫苗開發中的應用[D].上海:華東理工大學,2013. YAN Y J.Construction of anasdbalanced-lethal system inEdwardsiellatardaand its potential application in multivalent vaccine development[D].Shanghai:East China University of Science and Technology,2013.(in Chinese)
(編輯 白永平)
Construction and Preliminary Application of Balanced Lethal System Based on AttenuatedSalmonellaSG97A
GENG Shi-zhong#,CHEN Xiao-juan#,PAN Zhi-ming,JIAO Xin-an*
(JiangsuKeyLaboratoryofZoonoses,JiangsuCo-InnovationCenterforPreventionandControlofImportantAnimalInfectiousDiseasesandZoonoses,YangzhouUniversity,Yangzhou225009,China)
The aim of this study was to deleteasdgene ofSalmonellagallinarum(SG) attenuated vaccine strain 97A and construct bivalent live vaccine based on host-vector balanced lethal system to express F protein of Newcastle disease virus(NDV).Two DNA fragments flankingasdgene as allele were amplificated by PCR andasdgene was replaced by chloramphenicol resistance(CmR) gene to construct the suicide plasmid pGMB151Δasd::CmR.After conjugation,the recombiant plasmid was transformed into SG97A fromE.coliχ7213,SG97AΔasd::CmRwas reversely screened out under the selectable pressure of chloramphenicol,DAP-dependent strain was named SG97AΔasd.Vector-host balanced lethal system was construct by pYA3334 containingasdgene being transferred into SG97AΔasdand to expressed F protein of NDV.SG97AΔasdwas successfully constructed by allelic exchanges,its growth was DAP-dependent.Vector-host balanced lethal system was constructed by pYA3334 being transferred to SG97AΔasd and could express F protein of NDV and also induce specific antibodies in infected chickens.Vector-host balanced lethal system was constructed successfully based on attenuatedSalmonellaSG97AΔasdin this study and could expressFgene of NDV as foreign gene to induce special antibodies in animal model,it laid a foundation for bivalent live vaccine based on attenuatedSalmonella.
Salmonellagallinarum(SG);asdgene;vector-host balanced lethal system;NDV-F protein
10.11843/j.issn.0366-6964.2015.12.016
2015-04-13
公益性行業(農業)項目(201403054);“863”項目(2011AA10A212);中國博士后基金(2014M551670);江蘇省重點實驗室開放課題(0273896034819);江蘇省博士后基金(1302067C);揚州大學創新培育基金(2013CXJ070)
耿士忠(1972-),男,江蘇高郵人,副教授,博士,主要從事沙門菌致病機制研究及疫苗開發,Tel:0514-87971136,E-mail:gszzsg115@163.com。耿士忠、陳曉娟對本文貢獻相同
*通信作者:焦新安,教授,E-mail:jiao@yzu.edu.cn
S852.612
A
0366-6964(2015)12-2243-08
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
