冬小麥品種‘蘭天23號’苗期抗條銹性遺傳分析
2015-03-23 07:24:08黃苗苗李亞凱賈秋珍孫振宇王曉明王萬軍曹世勤金社林
植物保護 2015年5期
關鍵詞:分析
黃苗苗,李亞凱,黃 瑾,賈秋珍,孫振宇,張 勃,王曉明,王萬軍,曹世勤*,金社林*
(1.甘肅農業大學草業學院,蘭州 730070;2.甘肅省農業科學院植物保護研究所,蘭州 730070;3.甘肅省天水市農業科學研究所甘谷試驗站,甘谷 741200)
小麥條銹病是中國及世界小麥生產上的最主要病害之一,種植抗病品種是防治該病最經濟有效且有利于環境保護的措施[1-4]。由于小麥條銹病菌生理小種變異快,小麥抗病遺傳基礎脆弱,致使品種抗病性喪失,引致病害流行危害。自1993年以來,隨著以條銹菌CYR31和CYR32為代表的Hy和水源致病類群的不斷分化和優勢小種毒性頻率的不斷上升,致使生產上大面積推廣種植的對條銹病的抗性持續達20余年的‘繁6’及其衍生系小麥品種抗病性喪失[5],導致2002年小麥條銹病在全國范圍內大流行,造成當年產量損失超過13億kg[2]。從2010年開始,中國農業科學院植物保護研究所和四川、甘肅省農業科學院植物保護研究所先后在四川及甘肅省監測到條銹菌新毒性菌系‘貴農22’致病類群(V26)[6-8],該致病類群中的代表菌系G22-9 和G22-14 不僅具有CYR32 和CYR33的致病特點,而且還對國內重要抗源材料‘92R’、‘貴農21’、‘貴農22’和‘Moro’具有很強的致病性,引致與這些材料有親緣關系的品種(系)及抗源材料先后在生產上喪失抗病性。研究發現,實現抗性基因多樣化是克服品種過快喪失其抗性有效可行的辦法[9]。甘肅隴南麥區是我國小麥條銹病的常發易變區和新小種策源地[1],不斷加強該區的重要生產品種及抗源材料的抗性遺傳基礎研究,同時進行抗病新資源的挖掘、評價和利用,將會增加抗病基因多樣性,對持續控制小麥條銹病發生流行具有積極的推動作用。目前國內外相關學者利用經典遺傳學方法,在品種抗條銹性遺傳分析方面開展了諸多研究,明確了供試材料的抗條銹基因數量及遺傳方式[10-14],為這些材料的更好利用打下了良好的基礎。
‘蘭天23號’是甘肅省農業科學院小麥研究所通過有性雜交選育而成的冬小麥新品種。該品種具有優異的抗條銹性,同時兼具良好的豐產性和生態適應性,目前已成為甘肅省主要的冬小麥生產品種。在其抗條銹性遺傳研究方面,國內尚未開展相關研究。基于此,作者開展了該品種苗期抗條銹性遺傳分析,旨在明確該品種抗條銹性遺傳基礎,為其被更好地利用提供依據。
1 材料與方法
1.1 供試材料
2010年5月上旬在甘肅省農業科學院植物保護研究所甘谷試驗站,以感病材料‘銘賢169’為母本,以‘蘭天23號’為父本進行雜交,6月下旬收獲F0及親本材料;2010年10月中旬分別種植各親本及部分F0代種子。2011年5月上旬,在F1代材料自交的同時,以F1代材料為母本,以‘銘賢169’為父本繼續進行雜交,分別獲得‘蘭天23號’和‘銘賢169’雜交組合后代F2和BC1代材料。上述各世代材料及親本‘銘賢169’、條銹菌單孢菌系CYR32、CYR33和G22-9均來自甘肅省農業科學院植物保護研究所小麥病害課題組。‘蘭天23號’來自甘肅省農業科學院小麥研究所。
1.2 試驗方法
苗期抗性鑒定和遺傳分析工作于2014年4-5月在甘肅省農業科學院植物保護研究所蘭州溫室進行。親本‘蘭天23號’、‘銘賢169’及雜交F1代、BC1代種子各播種1盆,F2代種子播種6~10盆,共播種3套。在小麥生長的1葉1心期,采用抖孢子粉法[10]分別接種條銹菌單孢菌系CYR32、CYR33和G22-9。接種后的幼苗置于10℃黑暗條件下保濕24h,之后置于常規溫室生長18d,待感病對照品種‘銘賢169’充分發病后,逐株調查記載各組合供試材料的反應型[10-11,14]。反應型記載采用0、0;、1、2、3、4共6級標準進行[1]。為保證遺傳分析試驗的準確性,在采用0~4級標準的基礎上,再加入1+、2-和2+3個級別[15-16],以進行輔助分析。對實測值的抗感比率與期望的比率進行χ2適合性檢驗[14,16]。
2 結果與分析
2.1 對CYR33抗條銹性遺傳分析
用CYR33的單孢菌系分別對‘蘭天23號’/‘銘賢169’組合的親本及F1代、BC1代和F2代材料進行抗病性鑒定和統計分析。結果(表1)顯示,‘蘭天23號’對CYR33 表現免疫,反應型為0 型;‘銘賢169’表現高度感病,反應型為4 型,F1代植株均表現抗病,反應型為0~0;型,F2代群體表現抗感分離。根據雙親、F1、F2、BC1代反應型級別及各級反應型數目,將0~2+型植株劃為抗病類型,3~4型劃為感病類型。198株F2代群體中,表現抗病的有144株,表現感病的有54株,經χ2檢驗,符合由1對顯性基因控制的3R∶1S的理論比例(χ2{3∶1}=0.520 8<=3.84,P=0.25~050)。12株BC1代植株中,抗病株和感病株各6株,符合由1對顯性基因控制的1R∶1S的理論比例(χ2{1∶1}=0.083 3,P=0.10~0.25),與F2代分析結果相一致。表明‘蘭天23號’對CYR33的抗病性由1對顯性基因控制。
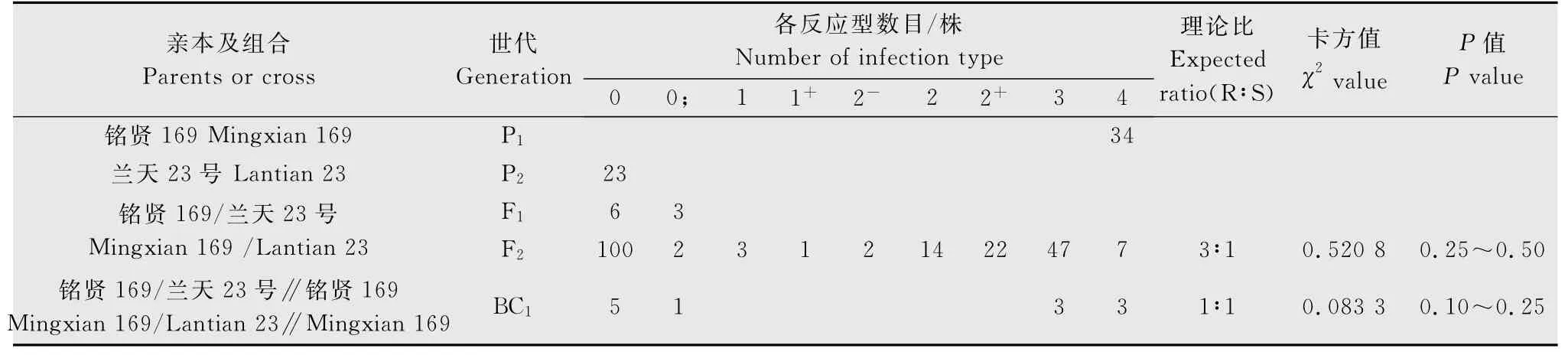
表1 ‘蘭天23號’與‘銘賢169’組合各世代對CYR33苗期抗條銹性表現Table 1 The resistance of wheat varieties‘Lantian 23’,‘Mingxian 169’and their progenies to Puccinia striiformis f.sp.tritici physiological race CYR33at seedling stage
2.2 對CYR32抗條銹性遺傳分析
用CYR32的單孢菌系分別對‘銘賢169’/‘蘭天23’組合的親本及F1、BC1和F2代材料進行抗病性鑒定和統計分析。結果(表2)顯示,‘蘭天23號’對CYR32表現免疫,反應型為0 型;‘銘賢169’表現高度感病,反應型為4型;F1代植株均表現抗病,反應型為0~0;型;F2代群體表現抗感分離。根據雙親、F1、F2、BC1代反應型級別及各級反應型數目[14],將0~2+型植株劃為抗病類型,3~4型劃為感病類型。在84株F2代群體中,表現抗病的有62株,表現感病的有22株,經χ2檢驗,符合由1對顯性基因控制的3R∶1S 的理論比例(χ2{3∶1}=0.111 1<=3.84,P=0.10~0.25)。11 株BC1代植株,5株表現抗病,6株表現感病,符合由1對顯性基因控制的1R∶1S 的理論比例(χ2{1∶1}=0.181 8,P=0.25~0.50),與F2代分析結果相一致。表明‘蘭天23號’對CYR32的抗病性由1對顯性基因控制。
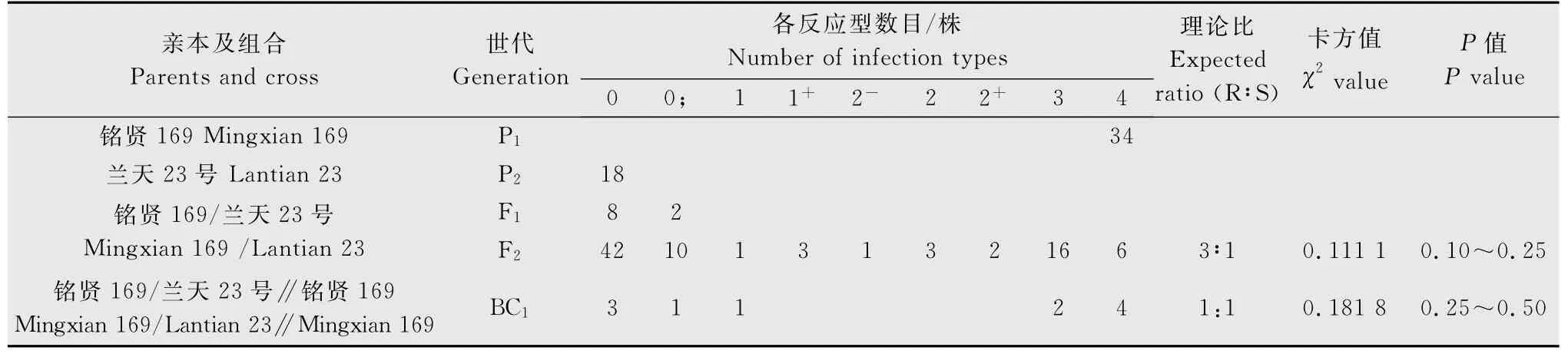
表2 ‘蘭天23號’與‘銘賢169’組合各世代對CYR32苗期抗條銹性表現Table 2 The resistance of wheat varieties‘Lantian 23’,‘Mingxian 169’and their progenies to Puccinia striiformis f.sp.tritici physiological race CYR32at seedling stage
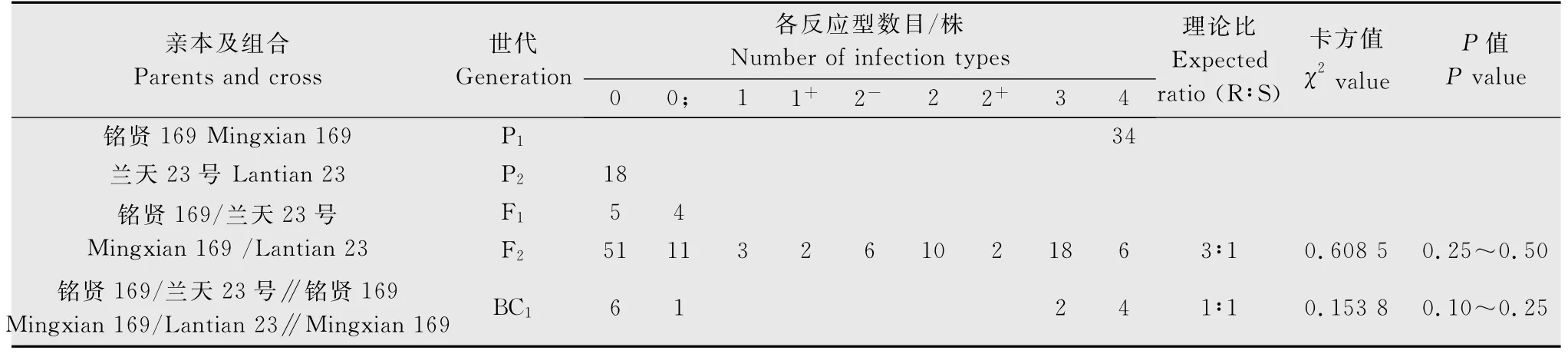
表3 ‘蘭天23號’與‘銘賢169’組合各世代對G22-9苗期抗條銹性表現Table 3 The resistance of wheat varieties‘Lantian23’,‘Mingxian169’and their progenies to Puccinia striiformis f.sp.tritici physiological race G22-9at seedling stage
2.3 對G22-9抗條銹性遺傳分析
用G22-9的單孢菌系分別對‘銘賢169’/‘蘭天23’組合的親本及F1代、BC1代和F2代材料進行抗病性鑒定和統計分析。結果(表3)顯示,‘蘭天23號’對G22-9表現免疫,反應型為0型;‘銘賢169’表現高度感病,反應型為4型,F1代植株均表現抗病,反應型為0~0;型,F2代群體表現抗感分離。根據雙親、F1、F2、BC1代反應型級別及各級反應型數目,將0~2+型植株劃為抗病類型,3~4型劃為感病類型。在109株F2代群體中,表現抗病的有85株,表現感病的有24株,經χ2檢驗,符合由1對顯性基因控制的3R∶1S 的理論比例(χ2{3∶1}=0.608 5<=3.84,P=0.50~0.75)。13株BC1代植株中,7株表現抗病,6株表現感病,也符合由1對顯性基因控制的1R∶1S的理論比例(χ2{1∶1}=0.153 8,P=0.10~0.25),與F2代分析結果相一致。表明‘蘭天23號’對G22-9的抗病性由1對顯性基因控制。
3 討論
Biffen研究證明小麥抗條銹性不僅是可以遺傳的,而且符合孟德爾遺傳規律[17]。Loegering和Brow der在Flor基因對基因學說為原理的基礎上,建立了通用的基因推導原則,并在禾谷類作物抗病基因分析方面得到廣泛應用[18]。曹世勤等通過該方法分析發現,‘蘭天23號’含有未知抗條銹病基因[19]。本研究表明,‘蘭天23號’對我國當前條銹菌主要流行小種CYR33、CYR32及新菌系G22-9的抗病性均由1對顯性抗性基因控制。分析其系譜,其親本材料為‘87-121’和‘SXAF4-7’。甘肅省農業科學院植物保護研究所自1996年開始,連續多年在甘肅隴南的汪川良種場和甘谷試驗站,對‘87-121’和‘SXAF4-7’的鑒定及監測結果發現,父本‘87-121’在苗期和成株期對接種的CYR33、CYR32和自然誘發的條銹菌均表現感病,平均病情3~4/40/100(反應型/嚴重度/普遍率),母本‘SXAF4-7’表現抗病,反應型0~0;型,屬免疫、近免疫類型(未發表資料)。故此推斷,‘蘭天23號’所含的抗條銹基因來源于母本‘SXAF4-7’。但對CYR33、CYR32和G22-9均為顯性抗性的基因是同一基因還是未知新基因,則有待于進一步研究。
徐建龍等[20]認為小種的選擇是推斷品種抗病基因來源的關鍵。在本試驗中鑒定條銹菌系選擇甘肅省及中國出現頻率高、毒力強的CYR33、CYR32及新毒性菌系G22-9作為供試菌系進行抗條銹基因遺傳分析,其研究結果對抗病親本選擇有重要指導意義。
[1]李振岐,曾士邁.中國小麥銹病[M].北京:中國農業出版社,2002.
[2]Wan A M,Zhao Z H,Chen X M,et al.Wheat stripe rust epidemic and virulence ofPucciniastriiformisf.sp.triticiin China in 2002[J].Plant Disease,2003,88(8):896-904.
[3]Mcintosh R A,Barinan H S,Park R F,et al.Aspects of wheat rust research in Australia[J].Euphytica,2001,119(1/2):115-120.
[4]Chen X M,Mary M,Euqene A M,et al.Wheat stripe rust epidemics and races ofPucciniastriiformisf.sp.triticiin the United States in 2000[J].Plant Disease,2002,86(1):39-46.
[5]吳立人,牛永春.我國小麥條銹病持續控制的策略[J].中國農業科學,2000,33(5):1-7.
[6]Liu T G,Peng Y L,Chen W Q,et al.First detection of virulence inPucciniastriiformisf.sp.triticiin China to resistance genesYr24(=Yr26)present in wheat cultivar Chuanmai 42[J].Plant Disease,2010,94:1163.
[7]劉太國,王保通,賈秋珍,等.2010-2011年度我國小麥條銹菌生理專化研究[J].麥類作物學報,2012,32(3):574-578.
[8]賈秋珍,黃瑾,曹世勤,等.感染我國重要小麥抗源材料貴農22的條銹菌新菌系的發現及致病性初步分析[J].甘肅農業科技,2012(1):3-5.
[9]Sharp E L,Volin R B.Additive genes in wheat conditioning resistance to stripe rust[J].Phytopathology,1970,60(7):1146-1147.
[10]曹世勤,駱惠生,黃瑾,等.冬小麥品種隴鑒9821抗條銹遺傳分析[J].植物病理學報,2012,42(3):274-280.
[11]姚強,賀苗苗,閻佳會,等.春小麥品種青春39的抗條銹性遺傳分析[J].麥類作物學報,2014,34(1):39-42.
[12]李洋,袁喜麗,姚強,等.歐洲小麥品種Mega抗條銹病基因的遺傳分析及分子標記[J].植物病理學報,2010,40(1):51-56.
[13]姚占軍,徐世昌,萬安民,等.3個小麥條銹菌鑒別寄主的抗性遺傳分析[J].植物遺傳資源學報,2006,7(1):39-43.
[14]仝淑玫,藺瑞明,何月秋,等.小麥抗源Holdfast和Flinor抗條銹病主效、微效基因的遺傳分析[J].中國農業科學,2006,39(11):2243-2249.
[15]徐世昌,張敬原,趙文生,等.小麥京核891-1抗條銹病主效、微效基因的遺傳分析[J].中國農業科學,2001,34(3):272-276.
[16]何家泌.植物抗病遺傳學[M].北京:中國農業出版社,1994.
[17]Biffen R H.Mendelps laws of inheritance and wheat breeding[J].Journal of Agricultura1Science,1905,1:4-46.
[18]Wellings C R,McIntosh R A.Pucciniastriiformisf.sp.triticiin Australasia:pathogenic changes during the first 10years[J].Plant Pathology,1990,39(2):316-325.
[19]曹世勤,張勃,李明菊,等.甘肅省50個主要小麥品種(系)苗期抗條銹基因推導及成株期抗病性分析[J].作物學報,2011,37(8):1360-1371.
[20]徐建龍,林貽滋,奚永安.城特232抗瘟性基因分析研究[J].植物保護學報,1994,21(1):63-66.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
