磷水平對不同基因型甘蔗苗期根系性狀的影響
2015-04-13 04:09:50鄭超李奇偉黃振瑞敖俊華劉可星
熱帶農業科學 2015年2期
鄭超+李奇偉+黃振瑞+敖俊華+劉可星
摘 要 采用盆栽砂培方法研究4個不同磷水平(0、0.01、0.1、1 mol/L)對粵糖93-159(YT93-159)、新臺糖22號(ROC22)和粵糖00-236(YT00-236)甘蔗苗期根系性狀的影響。結果表明:隨著磷水平的提高,甘蔗生物量增加,地上部生長迅速,根冠比顯著降低;低磷脅迫能促進根系生長,在一定磷濃度范圍內,形態指標值隨著磷水平的升高而提高;高磷(1 mmol/L)會抑制YT93-159和YT00-236根系的生長。根系活力和ATPase活性均隨施磷量的增加有上升的趨勢,根系活躍吸收面積始終保持在49%左右。YT93-159在低磷脅迫下,根系生長最發達,不同磷水平引起的根冠比變化值比ROC22、YT00-236小;YT93-159的根重、地上部重、根系活力、ATPase活性等均最大,ROC22次之,YT00-236最小。
關鍵詞 磷素水平 ;甘蔗 ;基因型 ;根系性狀
分類號 S566.1
磷是植物生長發育所必需的大量營養元素,它不僅是植物體的組成成分,也是植物體內能量載體的主要組分和提供者,在植物的光合作用、呼吸作用、糖分代謝、酶促反應和生理生化調節過程中起著至關重要的作用[1]。由于磷容易被土壤固定,且擴散系數很小,因此土壤磷的作物有效性很大程度上取決于作物根系的生物學特性,如根長、根半徑、根系比表面積、根構型及根分泌物等[2-3]。根系是構成植物體的主要部分,也是植物與外界環境進行物質和能量交換的場所,其分布特征反映了植物對環境的利用程度。植物根系可通過一系列的形態、生理變化主動適應低磷脅迫并改善自身的磷素營養狀況。在缺磷逆境中,植物通常為尋求磷源,根系發育增強、伸長,側根的長度和密度都增加,以加強對磷的吸收[4-5]。在玉米、小麥、大豆等作物中已發現植物可在磷脅迫下通過對根毛的分化、側根與不定根的發生、根冠比增加與根軸增長等方式改變自身根系形態和根系構型來提高對低磷環境的適應[6-7]。
以往對甘蔗的研究主要集中在磷對甘蔗葉的營養、生理、產量、品質和土壤養分等方面,而有關施用磷肥對甘蔗根系性狀的影響鮮有報道。鑒于此,本試驗研究不同磷水平對不同基因型甘蔗的根系形態特征、理化特性的影響,以期為甘蔗合理施磷提供理論依據,同時探討磷對甘蔗根系生長發育的影響及其重要性,為磷高效基因型甘蔗的篩選和遺傳改良提供科學依據。
1 材料與方法
1.1 材料
供試甘蔗品種組培苗:新臺糖22號、粵糖00-236和粵糖93-159(以下簡稱ROC22、YT00-236、YT93-159)。
供試基質:粒徑為0.6~2 mm的粗細適宜的河砂,用清水淘洗去除泥土和粉砂,然后用鹽酸洗滌(使試驗過程中不從砂中向培養液中溶解出鹽類且不吸附鹽類),用自來水清洗至中性,最后用去離子水清洗2次。
營養液配制:依據甘蔗對N、P、K的吸收特性對Hoagland 營養液配方進行適當改良。KNO3 252 mg/L,Ca(NO3)2·4H2O 506 mg/L,MgSO4 246 mg/L,ZnSO4·7H2O 0.21 mg/L,CuSO4·5H2O 0.1 mg/L,H3BO3 2.86 mg/L,(NH4)6Mo7O24·4H2O 0.01 mg/L,MnCl2·4H2O 1.81 mg/L。鐵鹽溶液(需單獨配制):FeSO4·7H2O 7.95 g,EDTA·Na 9.3 g,蒸餾水500 mL,稀釋1 000倍。KH2PO4 139 mg/L(單獨配制),分別稀釋1、10、100倍。
1.2 方法
1.2.1 試驗設計
試驗于2009年8月26日在廣東海洋大學興農樓5樓玻璃溫室進行。試驗設置3個甘蔗品種(ROC22、YT00-236、YT93-159),4個磷水平(0、0.01、0.1、1 mol/L,以下簡稱P0、P1、P2、P3),共12個處理,每個處理重復4次,總共96桶。每桶裝砂1.5 kg,砂桶底部留有小孔,浸入盛裝營養液的桶5 cm深。選用20 cm左右、長勢均勻的甘蔗組培苗移植(移苗應在10:00 a.m.前或17:00 p.m.后進行),每桶移栽2株。試驗均按照溫室試驗管理,每天按時補充去離子水至營養液為種植時的水平,防止由于水分減少而導致營養液的養分濃度改變,調節營養液的pH值至6.0,每周更換1~2次營養液。在移栽30~40 d后,收獲并測定各個根系的性狀。
1.2.2 測定項目及方法
(1)植株生物量的測定:采用烘干測重法,首先將地上部與根部分別稱取鮮重,然后在105℃下殺青,而后在60℃下烘干至恒重,再分別稱重。
(2)根系形態的測定方法:將植株根系鮮樣充分清洗整理后,運用數字化掃描儀將根系圖像掃描存入電腦,再利用根系圖像分析軟件對根圖像進行分析,最后獲取根長、根尖數、根體積等根系形態參數。
(3)根系活力的測定:分別將側根根尖的根毛區切斷,稱取不同處理相同部位側根根毛區0.5 g,具體測定方法采用氯化三苯基四氮唑(TTC)法。以μg TTF/(g FW·h)為單位表示活力大小。
(4)根系總吸收面積和活躍吸收面積的測定:用量筒排水法分別量取不同處理相同部位側根根毛區鮮樣1.0 mL,具體測定方法采用甲烯藍法。
(5)根系ATPase活性的測定:參照Chen等[8]的方法。
1.3 數據處理
采用Microsoft Excel 2003和SPSS(V17.0)軟件對數據進行統計分析,利用Duncan法進行多重比較,顯著性水平α=0.05。
2 結果與分析
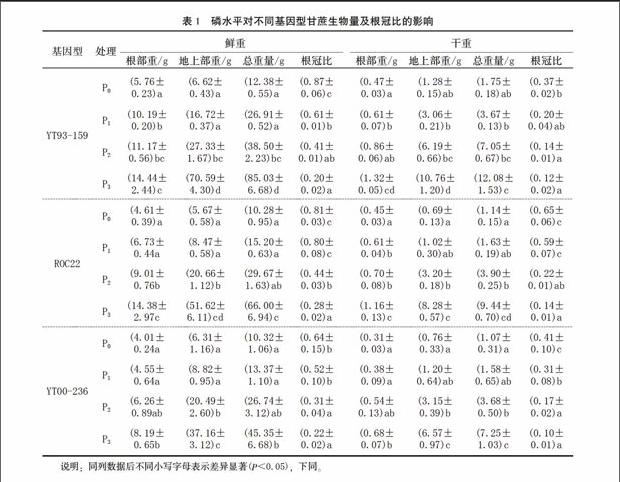
2.1 磷水平對甘蔗生物量及根冠比的影響
由表1可見,施磷使甘蔗生物量明顯增加,根部重和地上部重均與磷濃度的變化呈正相關。隨著磷濃度的增加,生物量呈上升的平滑曲線變化,YT93-159的增加量明顯最高,ROC22次之,YT00-236最低。說明施磷對甘蔗生物量的積累有顯著的促進作用。YT93-159的生物量既與品種遺傳特性有關,也與其對磷的利用效率有關。
根冠比的變化被認為是植物對低磷的耐受機制之一,是植物對低磷脅迫適應性反應和耐低磷能力的標志[6]。中國對小麥、油菜、玉米、水稻等的研究結果也表明,缺磷時,不同作物品種間的地上部分干重變化很大,磷高效品種的根冠比遠大于磷低效品種[6,9-10]。從P0處理到P1處理,YT93-159、ROC22、 YT00-236的鮮重根冠比依次下降26%、1%、12%,干重根冠比依次下降17%、6%、10%。初步說明,YT93-159在低磷環境下仍能保持正常的生長速度,屬于耐低磷品種。從P0處理到P3處理,YT93-159、ROC22、YT00-236的鮮重根冠比依次下降67%、53%、42%,干重根冠比依次下降25%、51%、31%。隨著施磷量的增加,各處理品種的根冠比呈下降趨勢。這說明光合產物在分配方向和強度上隨磷素水平的改變而變化,高磷供應使光合產物分配到地上部的比例增大,以促進莖葉的生長發育,提高作物產量;缺磷或低磷脅迫使光合產物分配到地下部的比例增大,以促使根系發達,擴大與土壤的接觸面積,從而提高根際養分有效性。而YT93-159的生長速度比較快。在低磷環境下,YT93-159的生物量最大,干重根冠比最小。由此可見,YT93-159對磷素反應敏感,能有效利用土壤中的磷素增加根重。
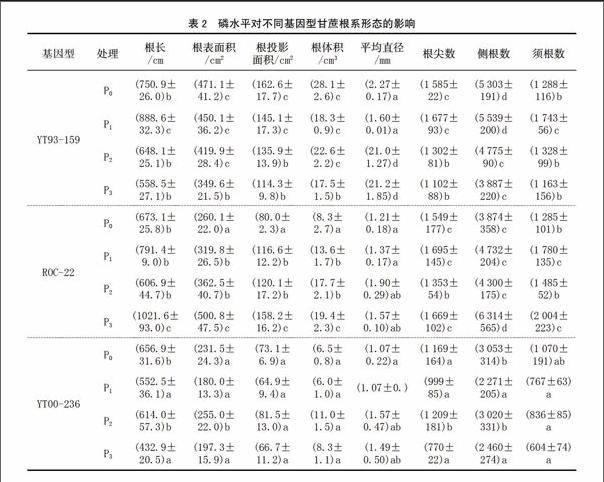
2.2 磷水平對甘蔗根系形態的影響
由表2可知,YT93-159在低磷條件下,根系生長得更好;ROC22和 YT00-236則是隨著磷水平的提高,根系生長得更好。但超過一定的磷濃度,根系的生長則受到抑制,尤其在YT93-159上表現的更加明顯。總之,施磷可促進根系的生長。
植物一般僅能吸收距根表面1~4 mm根際土壤中的磷[11]。植物受低磷脅迫時,根系發育加快,以縮短磷離子擴散到植物根的距離和擴大根系的吸收面積。植物適應低磷脅迫的一個重要的反應就是改變根系的生長及構型[12]。本試驗中,在P0和P1處理下,各基因型品種的根系各項形態指標值明顯增加,尤其YT93-159在各處理中表現得更加明顯。在P1處理下,YT93-159除根尖數和須根數僅次于ROC22外,其余數值均比相同磷水平的ROC22和 YT00-236高,并且在YT93-159的所有磷水平處理中排在第1位,可見,低磷脅迫下,YT93-159的根系生長反而更加好,說明YT93-159可能是耐低磷品種,低磷脅迫使其根系發達,以吸收更多的養分來維持正常的生長。因此,YT93-159耐低磷能力最強,ROC22次之,YT00-236最低。因為根直徑與磷的吸收效率呈反比,在根重相同情況下,根越細,磷的吸收效率越高[13];而其他根系性狀如根長、側根數、須根數等均與磷吸收效率呈正比,表明低磷促進YT93-159根系的生長,高磷則抑制根系的生長。
2.3 磷水平對甘蔗根系活力的影響
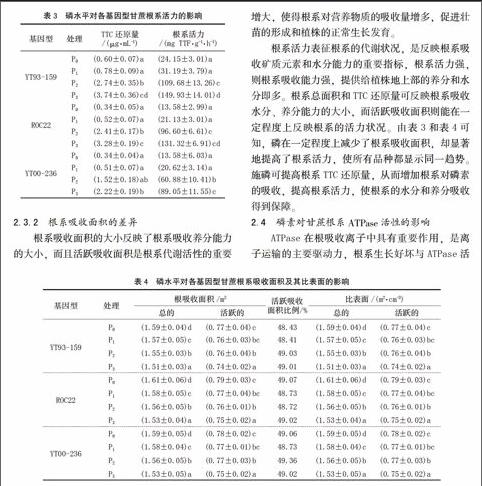
2.3.1 根系TTC還原量的差異
植物根系是活躍的吸收器官和合成器官,根的生長狀況和活力水平直接影響地上部生長、營養狀況及產量水平。根系活力大小一定程度上反映了作物吸收養分能力的強弱,一般情況下,根系活力越高,吸收養分的能力越強。根系TTC還原量通常作為衡量根系活力大小的有效指標。由表3可知,各基因型的TTC還原量均隨著磷水平的提高而增加,根系活力也呈上升趨勢,并且在P3達到最大值,說明磷可提高根系活力。此外,YT93-159在各個磷水平中根系活力最大,尤其在低磷條件下表現得更加明顯。ROC22和 YT00-236在P0與P1處理時,差異并不明顯,而在P2與P3處理時,ROC22的TTC還原量與根系活力明顯高于YT00-236。
2.3.2 根系吸收面積的差異
根系吸收面積的大小反映了根系吸收養分能力的大小,而且活躍吸收面積是根系代謝活性的重要參數。由表4可知,各基因型甘蔗根系吸收面積隨磷濃度的升高有一定的下降趨勢,且呈顯著性差異。不同基因型甘蔗在相同磷水平處理下根系吸收面積無顯著性差異。以所有處理的根體積為1 cm3計算,從P0處理到P3處理,YT93-159、ROC22、YT00-236的根系總吸收面積依次降低為5.04%、4.79%、3.33%,根系活躍吸收面積依次降低為3.84%、4.73%、3.25%。這表明無磷或低磷供應能在一定程度上提高根系的吸收面積,此結果與前述低磷對根系性狀的影響一致。由于根系吸收面積的增大,使得根系對營養物質的吸收量增多,促進壯苗的形成和植株的正常生長發育。
根系活力表征根系的代謝狀況,是反映根系吸收礦質元素和水分能力的重要指標,根系活力強,則根系吸收能力強,提供給植株地上部的養分和水分即多。根系總面積和TTC還原量可反映根系吸收水分、養分能力的大小,而活躍吸收面積則能在一定程度上反映根系的活力狀況。由表3和表4可知,磷在一定程度上減少了根系吸收面積,卻顯著地提高了根系活力,使所有品種都顯示同一趨勢。施磷可提高根系TTC還原量,從而增加根系對磷素的吸收,提高根系活力,使根系的水分和養分吸收得到保障。
2.4 磷素對甘蔗根系ATPase活性的影響
ATPase在根吸收離子中具有重要作用,是離子運輸的主要驅動力,根系生長好壞與ATPase活性高低密切相關[14],ATPase活性的增強是根系代謝旺盛的重要標志。本試驗結果表明,磷對根系ATPase活性產生顯著的影響。由表5可見,從P0到P3,YT93-159、ROC22、 YT00-236的ATPase活性依次提高了4.11、3.12、3.47倍。P0與P1之間ATPase活性差異并不顯著,到P2時出現顯著差異,P3時出現極顯著差異。所有的根系ATPase活性均表現出隨供磷水平的提高而增強的趨勢,說明供磷能提高根系的吸收能力和能量代謝的水平,磷對ATPase活性具有重要的促進作用。此外,在不同磷水平處理下,YT93-159的ATPase活性最高,ROC22次之,YT00-236最低。因此,YT93-159根系對養分的吸收較強,具有較強的生理活動能力,根系新陳代謝也較為旺盛。
3 討論與結論
根系是作物吸收水分和養分的重要器官,也是較先感受并傳導養分脅迫信號的器官,尤其對磷的虧缺反應敏感。磷脅迫條件下,作物地上部的光合產物向根部轉移量相對增加,使根冠比增大[15]。楊瑞吉等[16]的研究結果發現,不同基因型小麥開花期的根干重均隨施磷量的增加而增加,磷脅迫下根數量增加,根系變長,根半徑減小,以適應低磷逆境。在本研究中,甘蔗也會通過改變其根構型與各種根系性狀等來適應低磷脅迫,各基因型甘蔗在P0和P1(0.01 mol/L)處理下,根長、根表面積、根投影、根體積、根尖數、側根數、根尖數均明顯增加,根平均直徑減小,地上部生長受抑制,根冠比顯著提高。隨著供磷水平的提高,只有ROC22的各根系指標值相應地增大,YT00-236和YT93-159的根系生長卻受到不同程度的抑制。這與王建霄等[17]的研究結果一致,即隨著磷濃度的增加,橡膠樹幼苗1~4級根的平均長度、數量和總長度隨著供磷濃度的增加,呈先上升后下降的趨勢。
高磷供應下(1 mmol/L),各基因型甘蔗的干物質積累都達到最大值,積累量大小為YT93-159>ROC22>YT00-236。這與曹愛琴等[18]研究結果不同基因型菜豆在高磷處理條件下其生物量和吸磷量均高于低磷處理的觀點相類似。此外,高磷在一定程度上抑制根系的生長,從而使光合產物向地上部運輸,降低了根冠比。同時,TTC還原量明顯增大,根系活力和ATPase活性明顯增強。
劉靈等[19]發現,在低磷處理下供試大豆基因型間生物量和產量具有極顯著的基因型差異,且根形態、構型與磷效率密切相關。另有研究為表明,植株根冠比、根長的增加被認為是植物適應低磷脅迫的特征,在低磷條件下,根冠比、根長增加顯著的物種或基因型具有較好的低磷適應性[20]。與正常供磷相比,低磷處理條件下山核桃根長、根表面積和根體積顯著降低[21]。本試驗結果表明,YT93-159在低磷脅迫下,根平均直徑最小,而根長、根尖數、側根數、根表面積等都達到最大值,并且根系生長明顯發達,由此推斷,YT93-159與ROC22、YT00-236相比,苗期相對耐低磷。由于在各個磷水平處理下,YT93-159的生物量、根長、根體積、平均直徑、根系活力和ATPase活性等均高于ROC22與 YT00-236,YT93-159的磷利用效率最高,ROC22次之,YT00-236最低。筆者還發現,在各個磷水平下,植物根系的活躍吸收面積無顯著差異,始終保持在49%左右,這說明一定范圍的根系活躍吸收面積是甘蔗生長的必須保障。
綜上所述,低磷脅迫在一定程度上促進了甘蔗根系的生長,適當供磷,促進地上部生長,降低根冠比,有利于生物量的積累,增強根系活力,使得根系代謝增強,因此磷對甘蔗的生長至關重要。目前,磷在土壤中的難溶性和難移動性決定了圍繞提高根際磷的溶解性、擴展根系的吸收范圍的研究始終是工作的核心。磷效率的廣泛變異性給人們以啟示,培育磷高效的作物品種具有巨大的潛力[22]。
參考文獻
[1] Abelson P H. A potential phosphate crisis[J]. Science, 1999, 283(11): 2 015-2 021.
[2] Gardner W K, Barber D A. The acquisition of phosphorus by Lupinus albus L. The probable mechanism by which phosphorus movement in the soil root interface is enhanced[J]. Plant and Soil, 1983, 70(1): 107-124.
[3] Ae N, Arihara J, Okada K, et al. Phosphorus uptake by pigeon pea and its role in cropping systems of the Indian Subcontinent[J]. Science(Washington D C), 1990, 248(3): 477-480.
[4] Burton H R, Kasperbauer M J. Changes in chemical composition of tobacco laminar during senescence and curing 1, plastid pigments[J]. Agriculture and Food Chemistry, 1985, 33(5): 879-883.
[5] 秦 松,劉大翠,劉 靜,等. 土壤肥力對煙葉化學成分及品質的影響[J]. 土壤通報,2007,38(5):901-905.
[6] 馬樣慶,梁 霞. 植物高效利用磷機制的研究進展[J]. 應用生態學報,2004,15(4):712-716.
[7] 何 蓉,程雪斌,胡秋芬,等. 云南12種豆科灌木飼料中主要磷形態分析[J]. 云南大學學報:自然科學版,2003,25(6):528-530.
[8] Chen J C, Fu W H. Determine for activity of membrane cooped ATPase in leaf cell[J]. Chinese Journal of Cell Biology, 1983, 5(3): 21-24.
[9] 孫海國,張福鎖. 缺磷脅迫下的小麥根系形態特征研究[J]. 應用生態學報,2002,13(3):295-299.
[10] 郭玉春,林文雄,石秋梅,等. 低磷脅迫下不同磷效率水稻苗期根系的生理適應性研究[J]. 應用生態學報,2003,14(1):61-65.
[11] 李慶逵. 現代磷肥的研究進展[J]. 土壤學進展,1986,2(1):1-7.
[12] Lynch J P. Root architecture and plant productivity[J]. Plant Physiology, 1985, 109(1): 7-13.
[13] 王 毅. 植物耐低磷脅迫遺傳學研究策略[J]. 熱帶農業科學,2004,24(4):34-40.
[14] 朱 林,張春蘭,沈其榮. 施用稻草等有機物料對連作黃瓜根系活力、硝酸還原酶、ATP酶活力的影響[J]. 中國農學報,2002,18(1):17-19.
[15] 劉國順,肖慶禮,王艷麗. 不同供磷能力的土壤施磷對烤煙根體積和根冠比以及根系傷流組分的影響[J]. 中國煙草學報,2009,15(2):28-32.
[16] 楊瑞吉,張小紅,王鶴齡,等. 不同基因型春小麥對磷脅迫適應性研究[J]. 西北植物學報,2005,25(11):2 314-2 318.
[17] 王建霄,羅 微,茶正早,等. 水培條件下不同磷水平對橡膠樹幼苗根系生長發育的影響[J]. 海南大學學報,2009,27(3):265-269
[18] 曹愛琴,廖 紅,嚴小龍. 低磷土壤條件下菜豆根構型的適應性變化與磷效率[J]. 土壤學報,2002,39(2):276-281.
[19] 劉 靈,廖 紅,王秀榮,等. 不同根構型大豆對低磷的適應性變化及其與磷效率的關系[J]. 中國農業科學,2008,41(4):1 089-1 099.
[20] 曹一平. 高等植物的礦質營養[M]. 北京:北京農業大學出版社,1988:137-144.
[21] 李永夫,金松恒,葉正錢,等. 低磷脅迫對山核桃幼苗根系形態和生理特征的影響[J]. 浙江林學院學報,2010,27(2):239-245.
[22] 樊明壽,張福鎖. 植物磷吸收效率的生理基礎[J]. 生命科學,2001,13(3):129-131.
