Cd2+對狗牙根、假儉草種子萌發及幼苗生長的影響
2015-04-15 08:01:02岑畫夢彭玲莉楊雪吳亞嬌廖源林李西陳其兵蔡仕珍
草業學報 2015年5期
岑畫夢,彭玲莉,楊雪,吳亞嬌,廖源林,李西,陳其兵,蔡仕珍
(四川農業大學風景園林學院,四川 成都 611730)
?
Cd2+對狗牙根、假儉草種子萌發及幼苗生長的影響
岑畫夢,彭玲莉,楊雪,吳亞嬌,廖源林,李西,陳其兵,蔡仕珍*
(四川農業大學風景園林學院,四川 成都 611730)
探討了Cd2+濃度對狗牙根與假儉草種子萌發和幼苗生長的影響。結果表明,兩草種種子的發芽勢、發芽率、發芽指數、活力指數、根長以及幼苗長度均隨Cd2+濃度的升高而降低;葉片葉綠素含量隨Cd2+濃度的升高略有降低;葉片丙二醛(MDA)含量隨Cd2+濃度的升高呈先降后升趨勢;狗牙根過氧化氫酶(CAT)活性隨Cd2+濃度升高呈先升高后降低趨勢, 在5.0 mg/L時顯著高于CK, 假儉草各處理濃度的CAT活性均低于CK;兩草種的過氧化物酶(POD)活性隨Cd2+濃度的升高呈先升高后降低趨勢, 并在10.0 mg/L時達到峰值;Cd2+濃度在5.0,10.0,20.0,40.0 mg/L時, 狗牙根和假儉草多酚氧化酶(PPO)活性均高于CK, 兩草種PPO活性隨Cd2+濃度的升高都呈先升高后降低趨勢。兩草種對Cd2+均有一定的耐受能力, 狗牙根耐受能力大于假儉草, 二者均可以應用于Cd2+輕度污染土壤。
Cd2+;狗牙根;假儉草;種子萌發;幼苗生長
隨著中國工業迅速發展,含有重金屬的工業污水大量排放,化肥、農藥和污泥等的廣泛使用,使土壤存在不同程度的重金屬污染。包括Cd、汞、鉛、鉻、砷等,其中重金屬污染以遷移性強、毒性大被列為五毒之首。Cd在植物體內較易富集和沉淀,并隨生物體轉移,一旦進入食物鏈后將會對人類健康造成威脅。根據國家《土壤環境質量標準》(GB15618-1995),對于不同 pH 的土壤,國家二級標準規定Cd含量0.3~1.0 mg/kg為輕度污染,大于國標二級則達到污染級。環境保護部與國土資源部《全國土壤污染狀況調查公報》顯示,我國鎘、汞、砷、銅、鉛、鉻、鋅、鎳8種重金屬為主的無機物超標點位占全部超標點位的82.8%,其中鎘污染位居8種無機污染物之首,達7%。南方土壤污染重于北方,西南、中南地區土壤重金屬超標范圍較大,且鎘的含量在全國范圍內普遍增加,在西南地區和沿海地區增幅超過50%,在華北、東北和西部地區增加10%~40%。Cd污染狀況越來越嚴峻,并成為當前環境治理中突出的問題之一。土壤在受到Cd污染后自我修復周期可能需要兩百年左右,因而嘗試植物修復可能是處理Cd污染比較有效途徑之一[1]。
Cd對植物有較強的毒害作用[2],可以使植物的光合作用受到抑制[3],具體表現為:葉綠體受到破壞,光合酶的活性和葉綠素的合成下降,葉綠素總量下降,致使光合速率降低;抗氧化酶保護系統活性下降,進而誘發植物體內活性氧爆發,引起膜脂過氧化[4-5],而活性氧的非酶抗氧化系統同樣受到抑制[6],植株生物量和養分分配產生變化[7], 從而影響植物的生長[8],甚至導致植株死亡。種子萌發是植物生活史的關鍵環節,也是對外界抵抗力最弱的階段。Cd2+脅迫下,植物種子發芽率降低[9],根系發育受阻[10-11],植株生長受到抑制,但由于部分植物生長快、適應性強,對Cd具有較強的吸收能力和耐性[3]。
草坪草在大地綠化美化中兼具觀賞價值和改善生態環境雙重作用。不可避免地需要種植在Cd污染的土壤中。假儉草(Eremochloaophiuroides)和狗牙根(Cynodondactylon)均為常用的暖季型草坪草,分布較廣,根深,耐旱,耐貧瘠,耐踐踏,侵占性和再生能力強,成坪快,覆蓋率高,是我國南方建植各類草坪及公路護坡、護埂、護堤的理想綠化地被植物[12-13]。也是工業廢棄地、鎘污染灌溉區等各類污染地修復的備選材料。但尚缺乏其對重金屬耐受能力的研究。種子萌發和幼苗生長是植物生活史的開始和生命周期中生命力最脆弱的階段,對不利環境因子的耐受能力最小。如果種子萌發和幼苗階段能夠在Cd污染地生存, 那么成苗植株在污染地的生存就不存問題了。研究Cd對兩種草坪草種子萌發和幼苗生長的影響、評估兩草種對Cd脅迫的耐受能力和Cd污染土壤的修復能力,對其用于Cd污染地的植被修復具有重要生產實踐意義和科學價值。本試驗以假儉草和狗牙根為研究對象,采用培養皿培養種子的方法,研究不同的Cd2+濃度對兩種草坪草種子萌發及幼苗生長過程中生理生化的影響,進而分析并比較兩種草坪草對Cd2+的耐受能力,最終為草坪草對Cd2+耐受性的研究提供理論數據和科學依據,并為處于土壤鎘污染脅迫下綠化草種的選擇提供一定的理論參考。
1 材料與方法
1.1 材料處理及方法
試驗于2014年5月在四川農業大學成都校區園林植物實驗室進行,供試材料來源于四川金種燎原種業科技有限公司。試驗前精選優質飽滿的假儉草、狗牙根種子,將種子分別用紗布包裹,0.1%的KMnO4溶液中浸泡20 min消毒后取出,蒸餾水洗去表面殘留的KMnO4溶液,逐粒均勻放入內鋪雙層定量濾紙、直徑為90 mm的培養皿中,每皿均勻放入100粒種子。CdCl2·2.5H2O(分析純)溶于蒸餾水中。水中氯化鎘對一般植物生長危害的臨界質量濃度為1.0 mg/L,而一般業內公認的標準,超積累植物體內的鎘含量要達到一般植物的100倍以上,考慮到供試材料為生命周期中抗逆性最脆弱的種子萌發和幼苗生長初期階段,將CdCl2·2.5H2O濃度設置到一般植物臨界危害質量濃度的100 倍(100 mg/L)以下,配制Cd2+濃度分別為5.0,10.0,20.0,40.0,80.0 mg/L的處理液。吸管將處理液加入培養皿中,至濾紙飽和,后將培養皿放入光強4000 lx、12 h光照/12 h黑暗、相對濕度90%左右、25℃人工氣候箱中恒溫培養。以蒸餾水為對照,每處理重復3次,共培養36皿。試驗期間保持濾紙濕潤,每36 h更換1次等體積的處理液,觀察種子萌發和幼苗生長情況,每天記錄種子萌發數,直到沒有種子萌發為止。種子萌發期間統計發芽率(17 d內的發芽率)、發芽勢、發芽指數和活力指數。第17 天時測定幼苗芽長、根長、葉片葉綠素含量、丙二醛以及抗氧化酶活性。
1.2 測定指標和方法
1.2.1 種子發芽與幼苗生長情況 依據《國際種子檢驗規程》(2012版)進行萌發試驗。每天8:00-9:00觀察種子發芽情況,并記錄種子發芽數,以有根有芽、胚根長到0.2~0.3 cm為發芽標準,計算相應時間的發芽勢(狗牙根10 d內的發芽率、假儉草12 d內的發芽率)、17 d內的發芽率;第17天用游標卡尺測量芽長(根頸至葉尖的長度)和根長,重復3次,隨機取10 株,計算每處理30株平均值。
發芽勢=[10 d(狗牙根)和12 d(假儉草)發芽種子數/供試種子總數]×100%
發芽率=(17 d發芽種子數/供試種子總數)×100%
發芽指數(GI)=∑(Gt/Dt)
式中,Gt為在t日內發芽的種子數,Dt為發芽天數。
活力指數(VI)=S×GI
式中,S為發芽t時間時胚根及胚軸的總長度。
1.2.2 幼苗生長和生理指標測定 第17 天每處理取0.5 g鮮葉,浸提法[14]提取和測定葉綠素含量,硫代巴比妥酸法[14]測定丙二醛(MDA)含量,采用愈創木酚法[15]測定過氧化物酶(POD)活性,紫外分光光度法[15]測定過氧化氫酶(CAT)活性和多酚氧化酶(PPO)活性。
1.3 數據處理
用Excel 2007和SPSS 20.0軟件完成數據統計分析。
2 結果與分析
2.1 Cd2+對兩草種種子發芽的影響
2.1.1 Cd2+對兩草種種子發芽勢和發芽率的影響 發芽勢是評價種子的發芽速度和發芽整齊度的重要指標。圖1所示,隨Cd2+濃度升高,狗牙根和假儉草發芽勢曲線呈平緩變化到快速下降趨勢,即Cd2+濃度小于10.0 mg/L,曲線較為平緩,濃度高于10.0 mg/L,曲線快速下降,且濃度越高,下降趨勢越明顯。其中,Cd2+濃度在5.0和10.0 mg/L時,狗牙根與CK發芽勢差異不明顯,而假儉草較CK分別上升了11.4%和4.5%;當Cd2+濃度為40.0 mg/L時,狗牙根的發芽勢較CK降了16.2%,假儉草較CK下降了18.4%(低于CK的50%);當Cd2+濃度為80.0 mg/L時,狗牙根的發芽勢較CK降了72.2%,僅為CK的18.2%,假儉草發芽率為0。說明低濃度(≤10.0 mg/L)Cd2+對狗牙根發芽勢的影響不明顯,高濃度(≥20.0 mg/L)Cd2+則明顯抑制,而對假儉草則表現低促高抑,且Cd2+對假儉草的影響大于狗牙根。
隨Cd2+濃度升高,狗牙根和假儉草種子發芽率均呈降低趨勢。其中Cd2+濃度低于20.0 mg/L, 狗牙根種子發芽率略低于CK, 而Cd2+濃度高于20.0 mg/L則大幅度下降, 80.0 mg/L時發芽率較CK低72.2%;假儉草種子發芽率在Cd2+濃度高于5.0 mg/L時就開始大幅度下降,當Cd2+濃度為40.0 mg/L時,發芽率較CK下降了90.0%以上(圖2)。表明Cd2+對狗牙根種子發芽率的抑制作用小于假儉草種子。
2.1.2 Cd2+對兩草種種子發芽指數和活力指數的影響 發芽指數可以衡量植物的發芽能力及活力。活力指數是種子發芽速率和生長量的綜合反映,是反映種子活力的更好指標。由圖3和圖4可知, 隨Cd2+濃度升高,狗牙根與假儉草種子發芽指數和活力指數均呈下降趨勢。其中, 狗牙根種子的發芽指數在Cd2+濃度低于20.0 mg/L時差異較小,但高于20.0 mg/L時顯著降低,當Cd2+濃度為80.0 mg/L時,發芽指數比CK降低了56.7%,活力指數降低了89.5%;而假儉草種子的發芽指數和活力指數在Cd2+濃度高于5.0 mg/L時就明顯下降,當Cd2+濃度為80.0 mg/L時發芽指數比CK降低了88.8%,活力指數降低了99.6%。
2.1.3 Cd2+對兩草種根長、芽長的影響 隨Cd2+濃度的升高,狗牙根和假儉草根長和芽長均顯著降低。狗牙根根長減少幅度明顯大于假儉草。當Cd2+濃度為5.0 mg/L時,狗牙根根長較CK下降了67.4%,芽長與CK差異顯著,當濃度為80.0 mg/L時的根長較CK下降了99.1%,芽長為0 cm;而假儉草根長在Cd2+濃度為5.0 mg/L時較CK下降了28.6%,芽長較CK降低了15.0%,當濃度大于20.0 mg/L時,根長受到明顯抑制,小于0.5 cm,芽長低于4.0 cm,在80.0 mg/L時根長為0,芽長僅1.1 cm,較CK降低了87.7%(圖5~8)。說明狗牙根和假儉草根長和芽長均受到明顯抑制,對根的抑制作用大于芽。
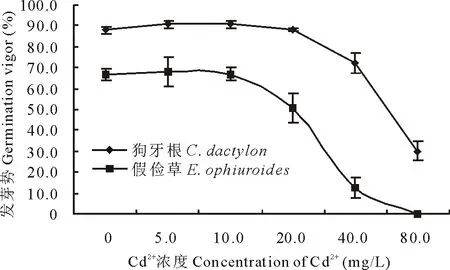
圖1 不同濃度Cd2+對兩草種種子發芽勢的影響Fig.1 Effects of different concentration of Cd2+ on the germination vigor of two turfgrass species
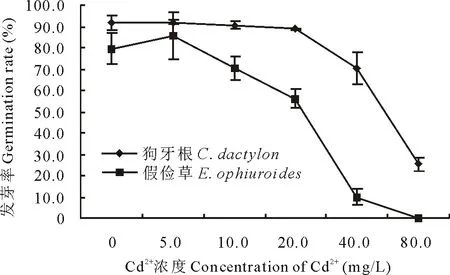
圖2 不同濃度Cd2+對兩草種發芽率的影響Fig.2 Effects of different concentration of Cd2+ on the germination rate of two turfgrass species

圖3 不同濃度Cd2+對兩草種種子發芽指數的影響Fig.3 Effects of different concentration of Cd2+ on the germination index of two turfgrass species
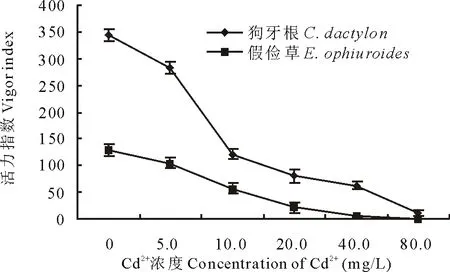
圖4 不同濃度Cd2+對兩草種種子活力指數的影響Fig.4 Effects of different concentration of Cd2+ on the vigor index of two turfgrass species
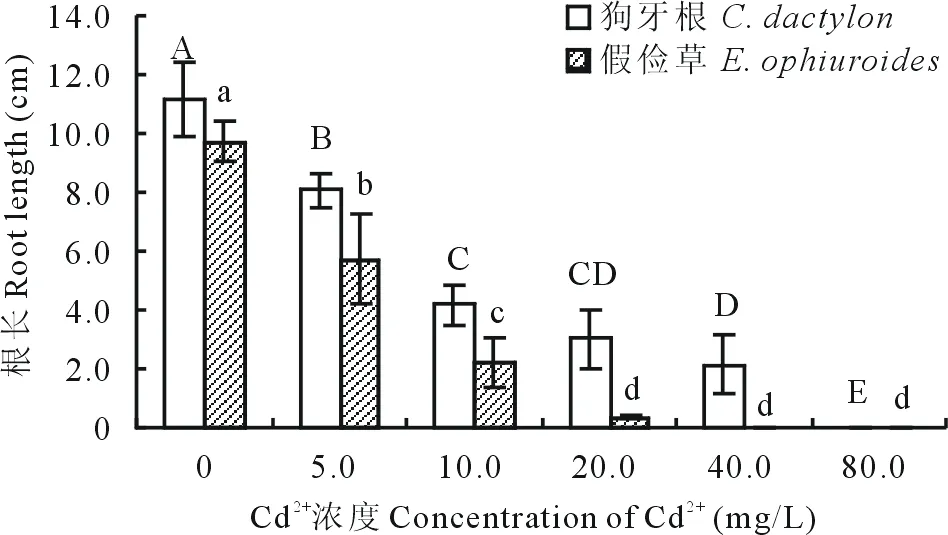
圖5 不同濃度Cd2+對兩草種根長的影響Fig.5 Effects of different concentration of Cd2+ on the root length of two turfgrass species
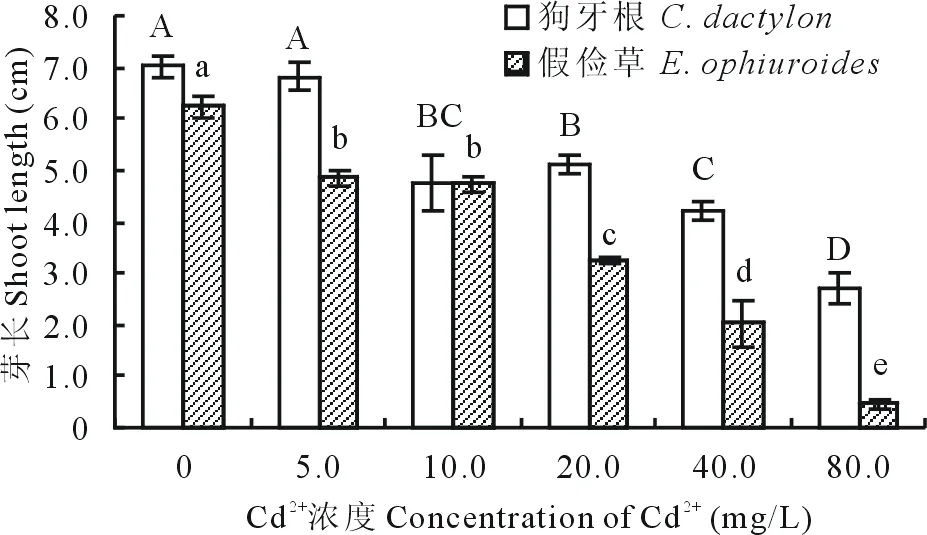
圖6 不同濃度Cd2+對兩草種芽長的影響Fig.6 Effects of different concentration of Cd2+ on the shoot length of two turfgrass species
不同字母表示差異顯著(P<0.05),下同。The different small letters mean the significant difference atP<0.05, the same below.
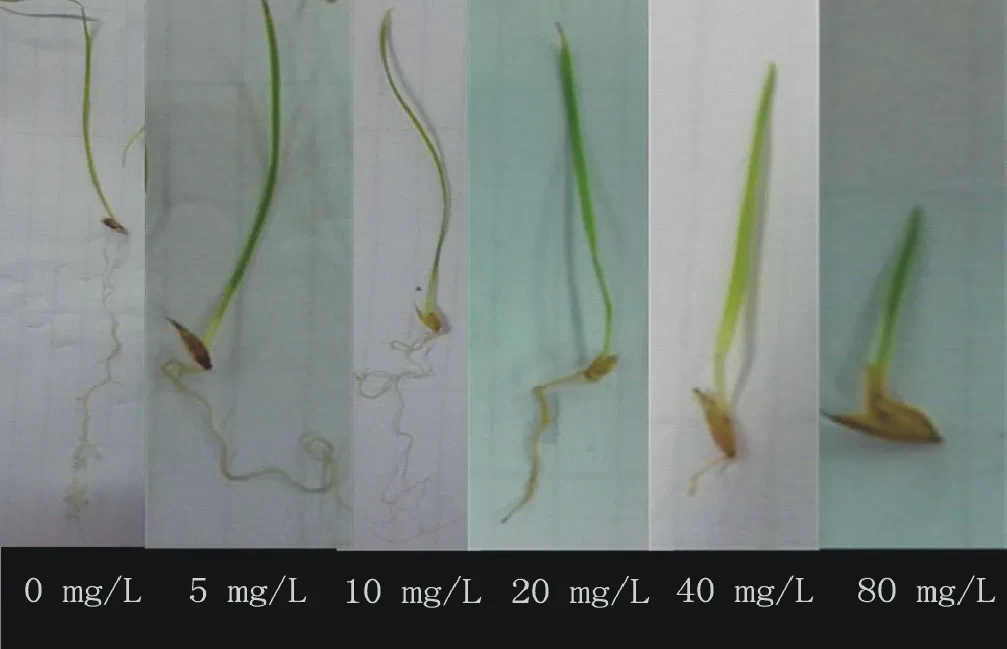
圖7 不同濃度Cd2+對假儉草幼苗生長的影響Fig.7 Effects of different concentration of Cd2+ on the seedling growth of E.ophiuroides
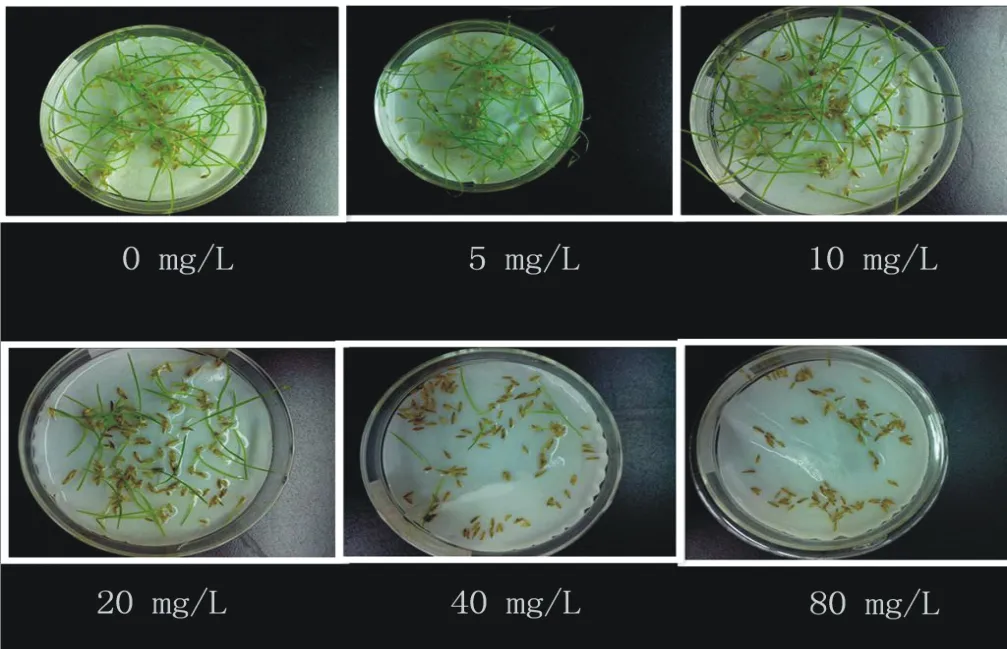
圖8 不同濃度Cd2+對狗牙根發芽的影響Fig.8 Effects of different concentration of Cd2+ on the germination of C.dactylon
2.2 Cd2+對兩草種幼苗部分生理指標的影響
2.2.1 Cd2+對兩草種幼苗葉綠素含量的影響 隨Cd2+濃度增加,狗牙根葉綠素a、葉綠素a+b含量呈降低趨勢,均在Cd2+濃度為80.0 mg/L時降至最低,分別較CK下降了77.9%和51.1%;葉綠素b含量略呈先升后降低趨勢,在Cd2+濃度為5.0 mg/L時達最高,比CK增加了27.5%,在各Cd2+濃度為80.0 mg/L時最低,較CK降低了17.6%(圖9)。假儉草的葉綠素a、葉綠素b和葉綠素a+b含量均隨Cd2+濃度增加呈降低趨勢,均在Cd2+濃度為80.0 mg/L時降至最低,分別較CK下降了34.7%,59.2%和41.4%。表明Cd2+能使狗牙根和假儉草葉片葉綠素含量降低,且隨著Cd2+濃度的加大影響加劇。
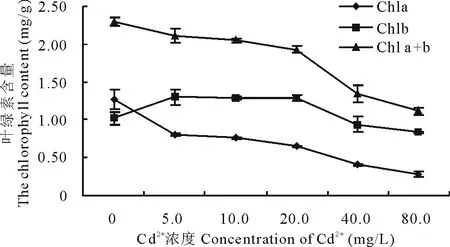
圖9 Cd2+對狗牙根葉綠素含量的影響Fig.9 Effects of Cd2+ on the chlorophyll content of C. dactylon
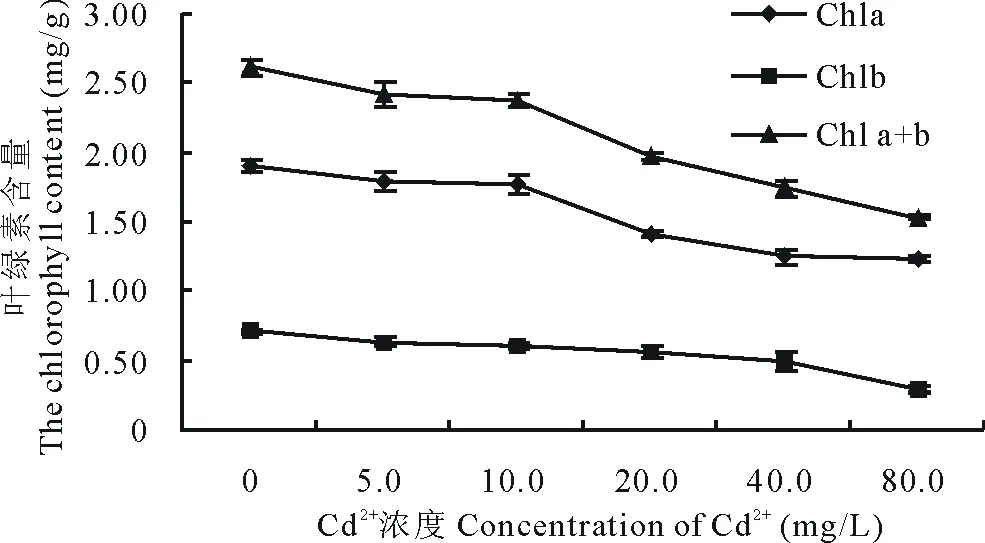
圖10 Cd2+對假儉草葉綠素含量的影響Fig.10 Effects of Cd2+ on the chlorophyll content of E. ophiuroides
2.2.2 Cd2+對兩草種幼苗MDA含量的影響 丙二醛(MDA)是衡量植物逆境脅迫下膜脂過氧化程度的指標,其在機體內的積累會對細胞產生毒害作用[2]。隨Cd2+處理濃度的升高,狗牙根和假儉草葉中的MDA含量均呈先下降后上升的變化趨勢,當Cd2+濃度為20.0 mg/L時,MDA含量最低,分別比CK下降了44.1%和23.4%,當Cd2+濃度達到80.0 mg/L時,MDA含量最高,分別比CK上升了45.1%和20.3%(圖11)。
2.2.3 Cd2+對兩草種幼苗CAT、POD、PPO活性的影響 CAT、POD是植物抗氧化保護酶類,在清除逆境脅迫下植物體內產生的大量活性氧方面發揮著很大作用[5]。由圖12和圖13可知,隨Cd2+處理濃度的升高,狗牙根葉片中CAT活性和POD活性均呈先升高后降低的變化趨勢,且CAT峰值出現在5.0 mg/L Cd2+時(是CK的2.3倍),POD峰值出現在10.0 mg/L Cd2+,CAT活性在Cd2+濃度高于5.0 mg/L急劇下降、而POD活性在Cd2+濃度高于20.0 mg/L急劇下降;假儉草葉片中CAT活性呈先降低后升高趨勢,谷值出現在10.0 mg/L Cd2+(較CK下降了1.1倍),而POD活性均呈先升高后急劇降低的變化趨勢,峰值出現在10.0 mg/L Cd2+。表明低濃度Cd脅迫刺激了CAT、POD活性升高,有利于活性氧的清除。
多酚氧化酶(PPO)是植物呼吸作用中重要的功能酶,其活性直接影響呼吸作用。圖14顯示,狗牙根和假儉草葉片中的PPO活性隨Cd2+濃度的升高呈先升高后降低的趨勢,其中狗牙根在5.0 mg/L Cd2+時最高,是CK的1.4倍,假儉草在20.0 mg/L時最高,約為CK的2倍,且狗牙根PPO活性在Cd2+濃度≤40.0 mg/L時均高于CK, 而假儉草各處理下PPO活性均高于CK。表明除狗牙根在80.0 mg/L Cd2+脅迫外,兩個草種的呼吸作用都沒有受到明顯抑制。
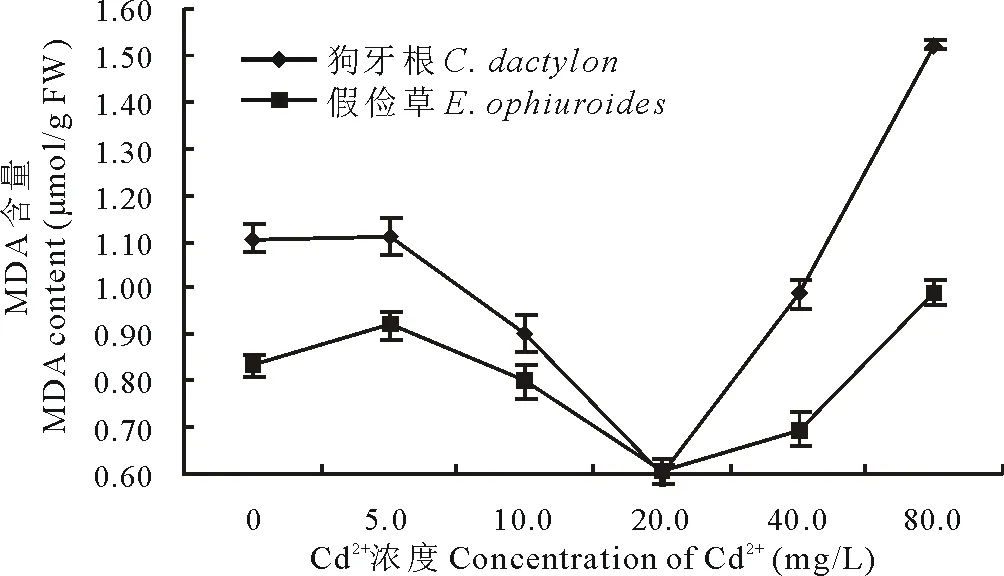
圖11 Cd2+對兩草種MDA含量的影響Fig.11 Effects of Cd2+ on the MDA content of two turfgrass species
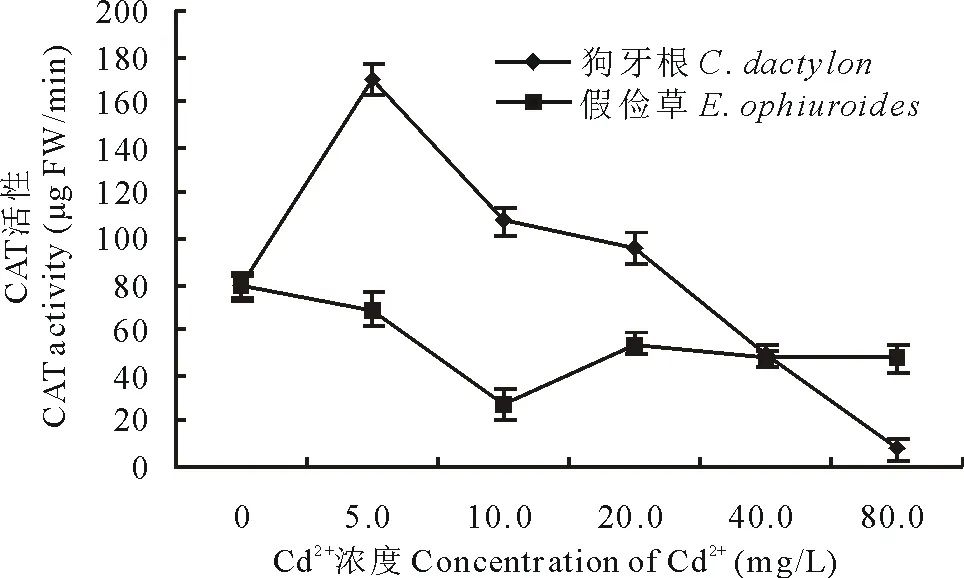
圖12 Cd2+對兩草種CAT活性的影響Fig.12 Effects of Cd2+ on the CAT activity of two turfgrass species
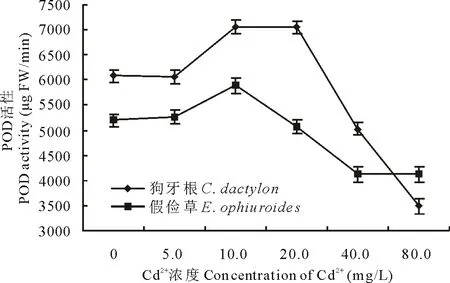
圖13 Cd2+對兩草種POD活性的影響Fig.13 Effects of Cd2+ on the POD activity of two turfgrass species
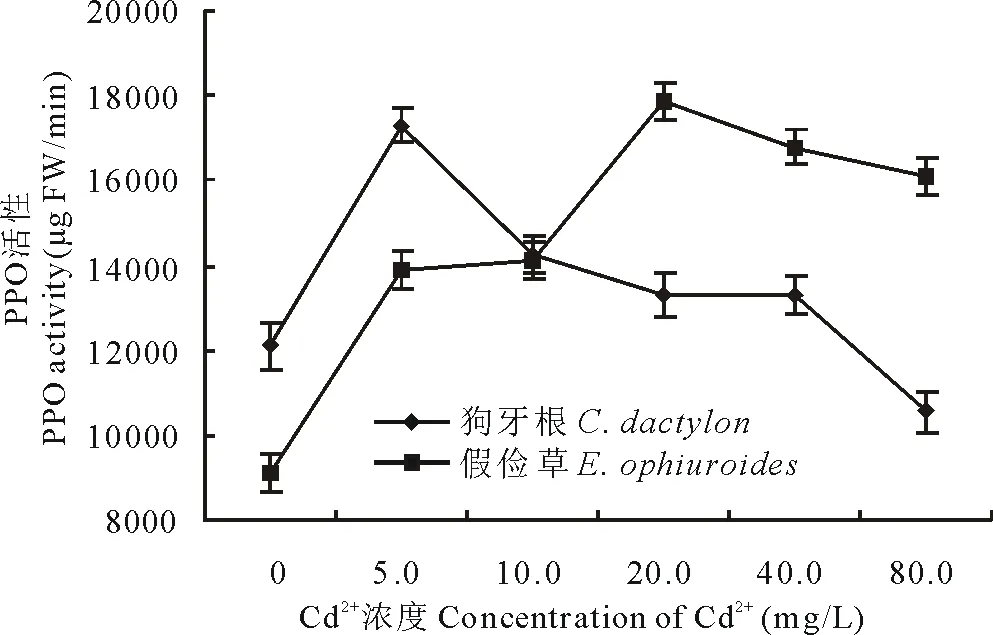
圖14 Cd2+對兩草種PPO活性的影響 Fig.14 Effects of Cd2+ on the PPO activity of two turfgrass species
3 討論
本研究結果顯示,Cd2+對假儉草和狗牙根種子萌發和幼苗生長具有一定的影響作用,隨Cd2+濃度的升高,種子的發芽率、發芽指數、根長、幼苗長度等都顯著降低,且具有明顯的濃度效應,這一結果與前人的研究結果一致[9]。Cd2+抑制狗牙根和假儉草種子萌發可能是Cd2+抑制種子內醇脫氫酶、淀粉酶、蛋白酶和酸性磷酸酶的活性,即抑制種子內儲藏淀粉和蛋白質的分解[16],從而影響種子萌發所需的物質和能量,致使種子萌發受到抑制。根是植物吸收和積累重金屬的主要器官,高濃度Cd2+脅迫使根細胞膜透性增加,損傷根系,干擾植物對水分和養分的吸收,抑制植物生長。所以高濃度Cd2+對兩種草種幼苗生長具有明顯抑制作用,而在80.0 mg/L Cd2+脅迫下,種子出現胚根不生長而胚芽生長的現象,體現了種子為逃避Cd2+對胚根的傷害而傷害全株,改變生物量的分配,將胚內有限的養分資源優先分配到胚芽的求生策略。Cd2+對兩草種的抑制作用有所不同,這可能與兩種植物的遺傳特性和其種子結構有關[11]。
葉綠素作為光合作用的重要色素,在光合作用中起著吸收、傳遞及轉化光能的作用。其含量可受重金屬脅迫的影響而下降,導致植物正常的生長發育受阻[17-18]。本試驗中,狗牙根和假儉草的葉綠素含量均隨Cd2+濃度的增加而降低。這可能是由于在Cd2+的脅迫下,葉綠體內的酶活性比例失調,破壞了葉綠體結構和功能,導致葉綠素含量下降。多酚氧化酶(PPO)是植物呼吸作用中重要的功能酶之一,作為植物呼吸鏈的末端氧化酶,它直接把呼吸底物的中間產物氧化脫出的電子傳遞給O2,催化酚及類似化合物被分子態氧氧化為醌類反應,該酶活性的下降將會對植物的呼吸作用產生影響[19]。試驗中,隨著處理濃度的增加,狗牙根與假儉草PPO活性先升高后降低,由此推測,Cd2+不僅影響狗牙根與假儉草的光合作用,一定程度上還影響呼吸作用。
植物在Cd2+脅迫下細胞膜透性遭到破壞,細胞膜透性增加,MDA含量會隨著Cd2+處理濃度的升高而增加[20]。狗牙根和假儉草在Cd2+濃度為80.0 mg/L時MDA含量達到峰值。隨著細胞膜膜脂過氧化的加強,自由基含量也隨之升高,CAT、POD是植物體內主要的抗氧化酶,能清除和減少植物體內的自由基,可防止細胞內過量自由基對植物造成氧化損傷[21]。本試驗中,隨著處理濃度增加,狗牙根CAT、POD活性與假儉草POD活性均呈先升高后降低,在80.0 mg/L時達到最低,這與Li 等[22]研究結果一致。可能是由于隨著Cd2+濃度的增加,植物細胞內產生的自由基逐漸積累,當在植物的耐受范圍內,為維持自由基含量與抗氧化保護系統之間的平衡,植物啟動應激機制,進而使抗氧化酶活性逐漸增加;而當脅迫濃度超出了耐受范圍,抗氧化保護系統遭到破壞,不足以平衡植物體內產生的自由基,導致植物體內過多的自由基積累,對植物產生傷害,使抗氧化酶活性不斷降低[23]。相對狗牙根CAT活性,假儉草CAT活性隨著Cd2+濃度的升高而顯著降低,說明假儉草較狗牙根更易受到Cd2+傷害。
高濃度Cd2+(≥20.0 mg/L)明顯抑制了狗牙根和假儉草的種子萌發和幼苗生長,而80.0 mg/L Cd2+脅迫達到兩草種的全致死條件,但Cd2+濃度≤20.0 mg/L,兩草種均有一定的耐受能力。這為重金屬污染日益嚴重的城市園林綠化土壤和Cd污染土壤的凈化治理中,有關草坪植物的選擇與通過播種途徑應用提供了參考依據。另外,本試驗僅研究了Cd2+脅迫對草坪植物種子及幼苗生長的影響,但對于Cd2+脅迫對狗牙根和假儉草生長的影響機理,以及草坪植物對重金屬Cd2+污染土壤修復效率等還有待進一步系統研究。
[1] Xu L J, Zhang M L, Yang H. Research progress of bioremediation technology of cadmium polluted soil. Journal of Nanjing Normal University (Natural Science Edition), 2011, 34(1): 102-106.
[2] Yang S Y, Wang F, Xie J C. Plant toxicity of heavy metals and the tolerant mechanisms of plants. Journal of Anhui NormaI UniverSity (Natural Science), 2004, 27(1): 71-74.
[3] Liu J X, Sun Z Y, Gou P,etal. Response of photosynthetic physiology of perennial ryegrass(Loliumperenne) to Cd2+stress. Acta Prataculturae Sinica, 2012, 21(3): 191-197.
[4] Zhou X Q, Mo C K. The plant heavy metal coercion and its antioxidation system. Journal of Xinjiang Education Institute, 2003, (2): 102-107.
[5] Liu J X. Research of Physiological Response Mechanism ofZoysiajaponicaon Cd Stress[D]. Beijing: China Forestry Science Research Institute, 2009: 11-15.
[6] Ding J J, Pan Y Z, Liu S L,etal. Effect and mechanisms of soil cadmium stress onDianthuschinensisseedling growth. Acta Prataculturae Sinica, 2013, 22(6): 77-85.
[7] Liu S L, Shi X S, Pan Y Z,etal. Effect of cadmium stress on growth, accumulation and distribution of biomass and nutrient inCatharanthusroseus. Acta Prataculturae Sinica, 2013, 22(3): 154-161.
[8] Liu Z L, He X Y, Chen W. Effects of cadmium stress on the growth and physiological characteristics ofLonicerajaponica. Chinese Journal of Applied Ecology, 2009, 20(1): 40-44.
[9] Liao G, Zhang Z F, Wang J L,etal. Effects of cadmium (Cd) concentration on seed germination ofRumexmaritimusandGeraniumcarolinianum. Grassland and Turf, 2011, 31(1): 47-49.
[10] Ge C J, Chen Q B, Yu H M,etal. Effect of Cd on germination and inhibition of root elongation of tropical forage plants. Chinese Journal of Tropical Crops, 2008, 10(5): 567-571.
[11] Bai R Q, Cao G P, Sun H,etal. Toxicity of cadmium on the seed germination and growth ofAlthaearoseaandOrychophragmusviolace. Acta Agriculturae Boreali-Sinica, 2009, 24(2): 134-138.
[12] Huang C Q, Zhang Y F, Liu G D. Research and improvement in germplasm resources ofCynodondactylon. Acta Agrectir Sinica, 2011, (5): 531-538.
[13] Liu H Q, Zhou S B, Xie C J. Progresses on germplasm resources of Centipedegrass (Eremochloaophiuroides) and its grazing-useage. Pratacultural, 2008, (1): 59-65. [14] Gao J F. Plant Physiology Experiment Tutorial[M]. Beijing: Higher Education Press, 2006.
[15] Li H S. Principle and Technology of Plant Physiological and Biochemical Experiments[M]. Beijing: Higher Education Press, 2003: 164-167.
[16] Shi N N, Chen Z W, Jia X Y. Effects of cadmium (Cd) stress on the sprouting of rice and its hydrolase activities. Agro-Environmental Protection, 1999, 18(5): 213-216.
[17] Shao G S, Chen M X, Wang W X,etal. Iron nutrition affects cadmium accumulation and toxicity in rice plants. Plant Growth Regulation, 2007, 53: 33-42.
[18] Wang K R, Zhou J L, Gong H Q,etal. Phytotoxic effect of soil cadmium pollution on ramie. Chinese Journal of Applied Ecology, 2000, 11(5): 773-776.
[19] Yang J R, He J Q, Jiang W R. Effects Cd pollution on plant physiological and biochemical. Agro-Environmental Protection, 1995, 14(5): 193-197.
[20] Zhang L H, Li P J, Li X M,etal. Effects of cadmium stress on the growth and physiological characteristics of wheat seedlings. Chinese Journal of Ecology, 2005, 24(4): 458-460.
[21] Ewa S P, Maria D, Zbigniew K. Lipid peroxidation and antioxidative response inArabidopsisthalianaexposed to cadmium and copper. Acta Physiologiae Plantarum, 2010, 32: 169-175.
[22] Li F T, Qi J M, Zhang G Y,etal. Effect of cadmium stress on the growth,antioxidative enzymes and lipid peroxidation in two kenaf (HibiscuscannabinusL.) plant seedlings. Journal of Integrative Agriculture, 2013, 12(4): 610-620.
[23] Deltoro V I, Gimeno C, Calatayud A,etal. Effects of SO2fumigations on photosynthetic CO2gas exchange, chlorophyll a fluorescence emission and antioxidant enzymes in lichensEverniaprunastriandRamalinafarinacea. Plant Physiology, 1999, 105: 648-654.
參考文獻:
[1] 徐良將, 張明禮, 楊浩. 土壤重金屬鎘污染的生物修復技術研究進展. 南京師大學報(自然科學版), 2011, 34(1): 102-106.
[2] 楊世勇, 王方, 謝建春. 重金屬對植物的毒害及植物的耐性機制. 安徽師范大學學報(自然科學版), 2004, 27(1): 71-74.
[3] 劉俊祥, 孫振元, 勾萍, 等. 鎘脅迫下多年生黑麥草的光合生理響應. 草業學報, 2012, 21(3): 191-197.
[4] 周希琴, 莫燦坤. 植物重金屬脅迫及其抗氧化系統. 新疆教育學院學報, 2003, (2): 102-107.
[5] 劉俊祥. 結縷草對Cd脅迫生理響應機制的研究[D]. 北京: 中國林業科學研究院, 2009: 11-15.
[6] 丁繼軍, 潘遠智, 劉柿良, 等. 土壤重金屬鎘脅迫對石竹幼苗生長的影響及其機理. 草業學報, 2013, 22(6): 77-85.
[7] 劉柿良, 石新生, 潘遠智, 等. 鎘脅迫對長春花生長, 生物量及養分積累與分配的影響. 草業學報, 2013, 22(3): 154-161.
[8] 劉周莉, 何興元, 陳偉. Cd脅迫對金銀花生態特征的影響. 應用生態學報, 2009, 20(1): 40-44.
[9] 廖鋼, 張志飛, 王俊麗, 等. Cd脅迫對野老鸛草和長刺酸模種子萌發的影響. 草原與草坪, 2011, 31(1): 47-49.
[10] 葛成軍, 陳秋波, 俞花美, 等. Cd脅迫對2種熱帶牧草種子發芽與根伸長的抑制效應. 熱帶作物學報, 2008, 10(5): 567-571.
[11] 白瑞琴, 晁公平, 孫華, 等. 重金屬Cd脅迫對蜀葵、二月藍種子萌發和幼苗生長的毒害效應研究. 華北農學報, 2009, 24(2): 134-138.
[12] 黃春瓊, 張永發, 劉國道. 狗牙根種質資源研究與改良進展. 草地學報, 2011, (5): 531-538.
[13] 柳后起, 周守標, 謝傳俊. 假儉草種質資源研究進展. 草業科學, 2008, (1): 59-65.
[14] 高俊鳳. 植物生理學實驗教程[M]. 北京: 高等教育出版社, 2006.
[15] 李合生. 植物生理生化實驗原理與技術[M]. 北京:高等教育出版社, 2003: 164-167.
[16] 施農農, 陳志偉, 賈秀英. Cd脅迫下水稻種子的萌芽生長及體內水解酶的活性變化. 農業環境保護, 1999, 18(5): 213-216.
[19] 楊居榮, 賀建群, 蔣婉茹. Cd污染對植物生理生化的影響. 農業環境保護, 1995, 14(5): 193-197.
[20] 張利紅, 李培軍, 李雪梅, 等. 鎘脅迫對小麥幼苗生長及生理特性的影響. 生態學雜志, 2005, 24(4):458-460.
Effects of Cd2+on the seed germination and seedling growth ofCynodondactylonandEremochloaophiuroides
QIN Hua-Meng, PENG Ling-Li, YANG Xue, WU Ya-Jiao, LIAO Yuan-Lin, LI Xi, CHEN Qi-Bing,CAI Shi-Zhen*
TheCollegeofLandscapeArchitecture,SichuanAgriculturalUniversity,Chengdu611730,China
The effect of Cd2+concentration on the germination and growth ofCynodondactylonandEremochloaophiuroideshas been studied. The results showed that the seed germination rate, germination vigor, germination index, vigor index, root length and seedling length of the two turfgrass species decreased with increases in Cd2+concentration. Chlorophyll content in leaves decreased slightly and the MDA contents in leaves showed a rising trend after an initial drop with increasing Cd2+concentration. The CAT activities ofC.dactylonincreased at first and then decreased with increasing concentrations. At a Cd2+concentration of 5.0 mg/L, CAT activities were significantly higher than that of CK, while the CAT activities ofE.ophiuroidesin all treatments were lower than that of CK. The POD activities of the two turfgrass species increased at first and then decreased with increasing Cd2+concentration, reaching its peak at 10.0 mg/L. The PPO activities ofC.dactylonandE.ophiuroideswere higher than CK when the concentrations were at 5.0, 10.0, 20.0 and 40.0 mg/L. With increasing Cd2+concentrations, the PPO activities of both species showed a declining trend following an initial rise. The two turfgrass species have a certain degree tolerance of Cd2+, with the tolerance ofC.dactylongreater than that ofE.ophiuroides. Both turfgrass species can thus be used in soils with low levels of Cd2+pollution.
cadmium;Cynodondactylon;Eremochloaophiuroides; seed germination; seedling growth
10.11686/cyxb20150512
http://cyxb.lzu.edu.cn
2014-11-13;改回日期:2014-12-31
四川省教育廳重點項目(12ZA116)資助。
岑畫夢(1991-),男,四川樂山人,在讀碩士。E-mail:70790545@qq.com *通訊作者Corresponding author. E-mail:415455088@qq.com
岑畫夢, 彭玲莉, 楊雪, 吳亞嬌, 廖源林, 李西, 陳其兵, 蔡仕珍. Cd2+對狗牙根、假儉草種子萌發及幼苗生長的影響. 草業學報, 2015, 24(5): 100-107.
Qin H M, Peng L L, Yang X, Wu Y J, Liao Y L, LI X, Chen Q B, Cai S Z. Effects of Cd2+on the seed germination and seedling growth ofCynodondactylonandEremochloaophiuroides. Acta Prataculturae Sinica, 2015, 24(5): 100-107.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
