膠孢炭疽菌侵染柱花草葉片的顯微觀察
2015-04-15 08:04:01楊茂霞林國彪陳彩虹王薈羅麗娟
草業學報 2015年5期
楊茂霞,林國彪,陳彩虹,王薈,羅麗娟
(海南大學農學院,海南省熱帶生物資源可持續利用重點實驗室,海南 海口 570228)
?
膠孢炭疽菌侵染柱花草葉片的顯微觀察
楊茂霞,林國彪,陳彩虹,王薈,羅麗娟*
(海南大學農學院,海南省熱帶生物資源可持續利用重點實驗室,海南 海口 570228)
為了研究膠孢炭疽菌在柱花草葉片上的侵染進程和植株接種病原菌后的反應,用膠孢炭疽菌孢子懸浮液接種生長6周的熱研2號柱花草幼苗,顯微觀察接種后炭疽菌在柱花草葉片上的侵染過程,統計植株病情指數。結果表明,接種4 h時孢子開始萌發、少量形成附著胞;6 h孢子大量萌發,12 h時大量附著胞形成并形成侵染釘;36 h時開始產生菌絲;48 h時菌絲大量產生糾結成網狀;60 h時開始形成分生孢子梗;72 h時產生新的分生孢子。接種72 h部分病原菌完成了一個侵染循環。同時柱花草發病癥狀為:葉片24 h黃化,有侵染點細胞膜破裂,48 h出現褐色病斑,72 h病斑出現黑褐色壞死線、點狀菌核、絨毛狀菌絲,96 h細胞崩解死亡,組織液外流。8 d病情指數為54.52,11 d有55%的接種植株死亡。
柱花草;柱花草炭疽菌;侵染;顯微結構;病害癥狀
柱花草(Stylosanthesespp.)是熱帶及亞熱帶地區最重要的豆科牧草之一,廣泛分布于澳大利亞北部、亞洲東南部、南美和非洲,具有草質好、產量高、耐旱、耐酸性貧瘠等特點[1],在我國的南方省區廣泛種植。炭疽病是危害柱花草生產的主要因素之一,該病造成莖葉枯萎脫落,嚴重影響產量[2]。柱花草炭疽病主要由刺盤胞屬膠孢炭疽菌(Colletotrichumgloeosporioides)引起,自1975年在南美的哥倫比亞爆發后迅速蔓延[3]。在我國華南地區,高溫高濕多雨條件下發病極快,3~5 d連續高溫多雨天氣柱花草葉片均發黃,出現不同程度的褐色病斑[4-5]。因此培育優良的抗病品種對柱花草的推廣種植至關重要。
炭疽菌屬(Colletotrichum)真菌在世界范圍內危害多種重要經濟作物,引起植物炭疽病。炭疽菌采用不同的致病機制來瓦解寄主的防御機制,包括胞內半活體侵染,角質層下層侵染,胞內死體侵染[6]。膠孢炭疽菌是一種半活體營養寄生菌((hemibiotroph),侵染特點包括在侵染初期短暫的活體營養階段和后期的死體營養兩個階段[7]。前人在柱花草膠孢炭疽菌的遺傳多樣性[8-10]、各柱花草品種的抗病性等方面的研究均有大量報道[11-12]。對柱花草炭疽病發病過程的研究則多集中在侵染中后期的寄主表型,缺乏病原菌侵染植株前期的細胞學過程,具體侵染行為、侵染時期、侵染方式等侵染循環的研究。
本研究通過對膠孢炭疽菌侵染柱花草葉片的動態顯微觀察及植株的病情統計,明確膠孢炭疽菌對柱花草葉片的侵染過程及寄主對病原菌侵染的反應,在細胞學水平上了解病原菌-寄主植物互作機制,對抗病育種及病害防治有重要意義。
1 材料與方法
1.1 實驗材料
供試柱花草品種為熱研2號(S.guianensiscv. Reyan2)。種子來自熱帶農業科學院儋州牧草中心,2014年5月14日播種,10 cm×10 cm苗盆每盆1粒,盆土為滅菌土∶腐殖土∶蛭石按2∶1∶1混合,置于海南大學農學院基地大棚內生長6周,種植期間大棚溫度為25~35℃,每天或隔天澆水保持土壤適當水分。
接種用病原菌為采自儋州牧草種質圃的柱花草病葉,單菌落分離的膠孢炭疽菌。將病原菌接種于直徑8 cm的PDA培養皿上,在溫度28℃條件下暗培養7 d,用無菌牙簽對平板劃線,培養3~5 d,直到平板大量產孢,無菌水沖洗孢子,雙層無菌紗布過濾,將洗出的孢子用無菌水配成濃度為1×106個/mL的孢子懸浮液,4℃保存備用,接種前加入0.2%的Tween-20。
1.2 接種方法
噴霧器均勻噴灑上述孢子懸浮液,直到植株葉片滴水。對照用加有0.2%Tween-20的無菌水噴灑,處理對照各40株,重復3次。接種植株28℃暗培養12 h,再置于溫度28℃,光照4000 lx,光照時長14 h,濕度大于90%的密閉接種室內培養,觀察并記錄植株病害癥狀,并于接種后第8天統計各接種植株炭疽病病情級別,據此計算病情指數,病情分級標準參照Kelemu等[8]的文獻記載。
1.3 材料的制備與顯微觀察
分別于接種后0,4,6,9,12,24,36,48,60,72,96,120 h,取處理、對照柱花草第3和4片完全展開葉。將采集葉片切成直徑5 mm小塊,用于制備不同樣品觀察。
制備觀察孢子形態及侵染特征的樣品。借鑒方中達[13]的真菌染色方法加以改進,將樣品用飽和的水合氯醛抽真空脫色36 h,期間12 h換1次脫色液。然后取出小葉用無菌水洗去殘留溶液,吸水紙吸干,0.5%的水溶苯胺藍(aniline blue)加熱沸騰后染色3 min,洗去染液,50%甘油保存。鏡檢時用水作浮載劑。顯微鏡觀察(Olympus BX51)接種后各時間點的病原菌形態發育特征,并統計孢子萌發,產生附著胞、侵染釘、菌絲、分生孢子梗和新分生孢子的時間點和百分率。孢子萌發,附著胞、侵染釘、菌絲產生每次隨機統計30個孢子,分生孢子梗產生及產孢每次隨機統計30個侵染點,3次重復。
制備觀察細胞致死及胼胝質產生的樣品。采用臺盼藍染色寄主細胞,如果膠孢炭疽菌侵入細胞膜,造成細胞膜被破壞,臺盼藍染液進入寄主胞質,寄主細胞將會被染成藍色。健康的柱花草細胞,臺盼藍不會進入胞質,細胞不會被染上顏色。參照Stone等[14]的方法,將樣品用乳酚(V酚∶V甘油∶V乳酸∶V水=1∶1∶1∶1)及2倍體積的乙醇脫色(抽真空15~30 min),65℃水浴30~45 min直到葉片透明,然后用乳酚沖洗,之后將樣品置于含2.5 mg/mL的臺盼藍染色液中(乳酚油配制),沸水浴2 min,冷卻染色1 h,然后在乳酚中脫色1 h。之后置于脫色苯胺藍染液(150 mmol/L K2HPO4,pH 9.5,0.01% Trypan Blue)中復染15 min,50%甘油保存。鏡檢用水作浮載劑。
制備電子顯微鏡觀察樣品。將采集葉片于冷凍干燥機(SCIENTZ-10N)中冷凍干燥9 h,然后平整貼于粘有導電黑膠的圓形銅臺上,送入真空鍍膜機中噴金,在S-3000N型掃描電鏡下進行觀察和拍照。
1.4 數據統計與分析

2 結果與分析
2.1 病害癥狀與病情指數
接種后的柱花草葉片大多從葉尖開始發病,病癥逐步向葉基蔓延。后期少數幼葉從中部發病,向四周蔓延。接種12 h內無明顯癥狀,24 h左右葉尖略微黃化,且病狀擴大。接種48 h,葉片開始出現0.5~1.0 mm大小褐色軟腐水漬狀病斑。接種72 h,病斑出現黑褐色壞死線、點狀菌核、絨毛狀菌絲病癥(圖1)。隨著病斑的擴大,發病葉片脫落,發病莖稈從植株中部靠上的位置折斷,接種后8 d統計病情指數為54.52,為感病。11 d統計死亡株數,55%的接種植株死亡。
2.2 接種孢子在柱花草葉片上的侵染循環
接種孢子首先附著于植株表面,接種4 h病菌孢子從端部或近端部萌發出棒狀芽管,6 h大量孢子萌發。隨后芽管頂端開始膨大,分化成特異的侵染結構附著胞,12 h超過50%的孢子產生附著胞,附著于寄主葉片表面,附著胞產生侵染釘。隨后分生孢子的端部再出芽產生第2個附著胞,或從芽管上分化出側枝產生附著胞,這個現象在接種24 h到72 h均觀察到。接種36 h孢子產生有分枝的菌絲,同時伴隨著接種孢子異化成有隔、間距短的粗壯菌絲。接種48 h大量菌絲生成,相互交錯成網狀,集聚在葉片上。接種60 h內生菌絲產生短棒狀的分生孢子梗,突破寄主表皮細胞在葉片表面呈花環狀排列。接種72 h部分孢子梗上產生新的分生孢子,成串排列,完成一個侵染循環,成熟孢子脫落附著在葉片上準備下一次侵染循環。病原菌通過不斷循環侵染植株,導致病害蔓延,直至整株發病死亡(圖2)。
以各侵染結構統計比率達到50%作為閾值,統計各侵染結構形成高峰期,得出柱花草炭疽菌感染寄主的5個關鍵時期:1)接種4~6 h為孢子萌發高峰期;2)接種9~12 h為附著胞及形成高峰期;3)接種36~48 h為菌絲形成高峰期;4)接種60~72 h為分生孢子梗形成高峰期;5)接種72~120 h為產孢高峰期。前兩個時期為侵染前期,柱花草葉片無明顯癥狀;第3個時期為病原物的潛育期,植株葉片出現病斑;后兩個時期為侵染后期,為炭疽菌第2次侵染宿主提供病原物,植株病斑擴大,有明顯病癥。
2.3 附著胞在寄主葉片上的發育進程及病原菌對寄主侵染結構的超微觀察
附著胞生長發育過程如下:首先分生孢子芽管頂端開始膨大(圖3A)。膨大部分生長形成有吸附作用的中間凹陷,四周凸起的吸盤狀結構,此時的附著胞顏色較淺,胞質較稀疏(圖3B)。當吸盤結構與柱花草葉片接觸一段時間后,其吸附口緊貼葉片,吸附于葉片表面,呈心形、水滴形及圓形等各種不規則形態,附著胞在形態的分化成熟過程中,附著胞胞壁逐漸黑色素化,胞質稠密(圖3C)。當附著胞附著在寄主葉片表面后,在中間位置形成1個圓形小亮點,即侵染釘,侵染釘穿透寄主角質層和細胞壁,開始第1次侵染(圖3D)。在第1次侵染之后,孢子會從兩端繼續萌發出芽管,多數形成2~3個,少數形成4~5個附著胞多位點侵染(圖3E、F)。
當病原菌識別寄主后,開始產生一系列的侵染結構:附著胞、侵染釘、侵染菌絲。除了通過上述的附著胞產生侵染釘侵染寄主(圖3D),電鏡掃描寄主上表皮觀察到接種后24 h病原菌從附著胞處形成侵染菌絲定植于寄主細胞(圖4A),形成內生菌絲在寄主細胞間蔓延。接種后36 h,寄主表面附著胞、表面菌絲穿透寄主細胞直接侵染寄主(圖4B)。

圖1 接種后熱研2號柱花草發病癥狀Fig.1 The symptom of S. guianensis cv. Reyan2 after inoculating

圖2 接種后膠孢炭疽菌在柱花草上的侵染循環Fig.2 Infecting cycle of C. gloeosporioide after inoculating Stylosanthese A:0 h接種孢子附著在葉片上 Inoculated spores adhere to the leaves at 0 h; B:6 h孢子萌發長出芽管 Germination of germ tube at 6 h; C:12 h孢子產生附著胞 Formation of appressorium at 12 h; D:24 h孢子產生多個附著胞 Formation of more than one appressoria at 24 h; E:36 h孢子上產生多條菌絲 Formation of multi-hyphea on one spore at 36 h; F:48 h生成大量菌絲且糾結成網狀 Formation of affluent hyphea at 48 h; G:60 h產生分生孢子梗 Generation of conidiophores at 60 h; H:72 h產生分生孢子 Production of new spores; C:分生孢子Conidium; GT:芽管Germ tube; Ap:附著胞Appressoria; PA:第1個附著胞Primary appressoria; SA:第2個附著胞Second appressoria; H:菌絲Hyphea; Co:分生孢子梗Conidiophores; NC:新產生的分生孢子New spores.
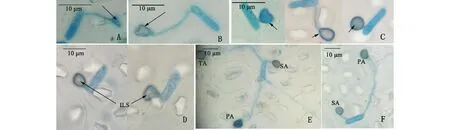
圖3 附著胞在植株上的生長發育進程Fig.3 Developmental progress of the appressoria on the plant A:芽管頂端膨大開始分化附著胞 Enlargement of the top of the germ tube; B:膨大部分形成吸盤狀結構附著胞 The enlargement part turns into appressoria with sucker shape and light color; C:吸盤狀附著胞吸附在寄主表面呈現心形、水滴形和圓形等不規則形態且細胞壁內黑色素加深 The appressoria adsorbs on the cuticle with the color become dark and presents irregular shapes such as heart, water-drop and roundness; D:附著胞中間亮點部分為侵染釘 Formation of penetration peg (internal light spot) and penetration; E、F:孢子形成多個附著胞多位點侵染Formation of multi-appressorium and penetration by more than one points; ILS:內部亮點Internal light spot; PA:第1個附著胞Primary appressoria; SA:第2個附著胞Second appressoria; TA:第3個附著胞Third appressoria.
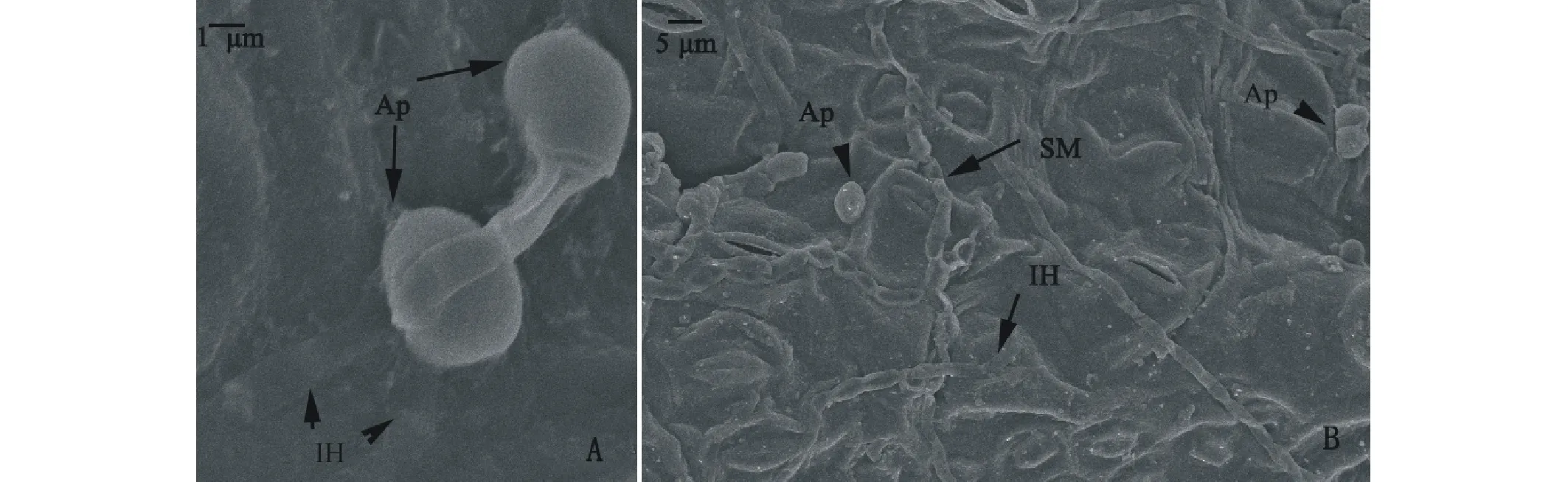
圖4 膠孢炭疽菌侵染柱花草葉片掃描電鏡觀察Fig.4 Observation of C. gloeosporioides infecting the leaves of Stylosanthese by SEM A:24 h侵染菌絲侵染柱花草葉片 Penetration hpyhea penetrate host leaves at 24 h; B:36 h附著胞及菌絲均侵染葉片 Plentiful appressorium and hpyhea penetrate host leaves at 36 h. PH:侵染菌絲Penetration hpyhea; SM:葉片表面菌絲Surface mycelium; IH:內生菌絲Internal hpyhea; Ap:附著胞Appressoria.
2.4 寄主細胞對膠孢炭疽菌侵染的反應
接種后0~24 h均未觀察到寄主細胞被臺盼藍染液染上藍色,24 h部分膠孢炭疽菌附著胞侵染點細胞被染色,表明接種后24 h膠胞炭疽菌初次侵入寄主細胞(圖5A、B),在侵染釘穿透寄主細胞壁后,形成初生菌絲穿透細胞膜,使寄主細胞染色。而部分菌產生多個附著胞、侵染釘多次侵染仍然沒有造成寄主細胞膜的破裂,侵染點細胞未著色(圖5C)。接種后96 h寄主細胞崩解,組織液外流粘附在葉片表面(圖5D、E),此時的病原菌已經進入了第2次侵染循環。
由于胼胝質與脫色苯胺藍反應,產生熒光反應。對各時間點采集葉片進行脫色苯胺藍染色、熒光顯微鏡觀察,均未檢測到熒光,說明寄主感病前期無胼胝質的形成。

圖5 膠孢炭疽菌侵染致死寄主細胞及寄主細胞的崩解Fig.5 The host cells killed by C. gloeosporioides and collapse A、B:接種后24 h附著胞侵入寄主細胞,臺盼藍進入細胞,侵染點細胞被染上藍色 The blue host cell near the infection point indicated that the appressoria have infected the host after 24 h inoculated; C:接種后48 h孢子產生5個附著胞,5個位點侵染寄主,無任何侵染點著色,未穿透寄主細胞膜 The colorless cell near the infection point indicated that the spores haven’t infected host after five times inoculated; D、E:96 h寄主細胞崩解,大量褐色組織液外流,粘附在葉片表面 After 96 h inoculated the host cell collapses, a lot of brown tissue liquid flows out and adheres to the leaf surface.
3 結論與討論
膠孢炭疽菌與豆炭疽菌(C.lindemuthianum)的侵染模式相同,均屬于胞內半活體營養菌(intracellular hemibiotrophy),與寄主建立了兩個營養關系:活體營養和死體營養[16-17]。活體營養階段,寄主細胞保持活力,病原菌形成一系列特殊的侵染結構。為了侵入寄主的表皮細胞,病原菌在識別寄主后開始萌發長出芽管,然后快速分化出高度特異的侵染結構附著胞。附著胞附著在寄主表面后,其內的滲透活性復合物逐漸合成黑色素,黑色素對寄主細胞壁有選擇滲透性,結合附著胞的滲透力會產生巨大的膨壓,形成侵染釘侵入寄主,寬的初生菌絲形成。半活體營養菌侵染過程中活體營養階段都較短,一般為一天到幾天[18-19]。之后窄的次生菌絲產生,其為更具有破壞性的死體營養菌,次生菌絲的產生標志著侵染進入了死體營養階段。死體營養階段,菌絲需穿透寄主細胞壁,在細胞間蔓延[20-22]。本實驗僅觀察到侵染釘、侵染菌絲的侵染,及菌絲在細胞內的蔓延;但未能從形態上直接區別初生菌絲和次生菌絲,因此不能確定整個侵染過程中活體營養階段和死體營養階段的分界點。最后寄主表面出現分生孢子梗并產孢,完成一個侵染循環,且確定其在接種后72 h就完成。
Ranathunge等[23]用平頭炭疽菌接種辣椒,孢子萌發始于接種后2 h,12 h大量產生附著胞,12~24 h侵染釘大量產生,72 h菌絲侵染。Vinijsanun等[10]用B型膠孢炭疽菌侵染愛德華柱花草(Endeavour),24 h產生附著胞和侵染釘,72 h內生菌絲形成,此時寄主細胞并未崩解。而本研究用B型菌侵染熱研2號柱花草,發病提前,接種后72 h病原菌完成侵染循環,96 h寄主細胞崩解。發病進程的差異與病原菌致病性、寄主抗病性、接種菌量和培養條件等有關。病原菌的致病性強,寄主感病,接種孢子濃度大,培養條件高溫高濕,則植株發病迅速,反之,植株發病較慢。本研究通過對接種葉片的侵染形態觀察,明確了炭疽菌各侵染結構的形成過程及形成的關鍵時期,并揭示了柱花草的整個發病過程、壞死及崩解反應;為柱花草與膠孢炭疽菌的互作,抗病相關代謝產物及抗病基因的研究提供了依據,同時也為病害防控奠定了基礎。
[1] Irwin J A G, Cameron D F, Lenne J M. The Biology and Agronomy ofStylosanthes[M]. Sydney, Australia: Academic Press, 1984: 73-101.
[2] Sonoda R M. Incidence ofColletotrichumleaf spot and stem canker on introductions and selections ofStylosantheshumilis. Plant Disease Reporter, 1973, 57(9): 747-749.
[3] Baldion R W, Lozano J C, Grof B. Evaluation of resistance ofStylosanthesspp. anthracnose (Colletotvichumgloeospovioides). Fitopathologia, 1975, 10(2): 104-108.[4] Feng S F, Li F E, He C Z. Bionomics and epidemiology go anthracnose onStylosanthesspp..Chinese Journal of Tropical Crops, 1994, 15(1): 87-94.
[5] Yi K X, Zheng J L, Xi J G,etal. Preliminary study of epidemic factors ofColletotrichumgloeosporioidesin stylo. Chinese Journal of Tropical Agriculture, 2014, 34(2): 74-78.
[6] Bailey J A, Jeger M. Colletotrichum: Biology, Pathology and Control[M]. Wallingford: UK CAB International Press, 1992: 88-120.
[7] Mendgen K, Venue M H. Plant infection and the establishment of fungal biotrophy. Trends in Plant Science, 2002, 7(8): 352-356.
[8] Kelemu S, Skinner D Z, Badel J L. Genetic diversity in South AmericanColletotrichumgloeosporioidesisolates fromStylosanthesguianensis, a tropical forage legume. European Journal of Plant Pathology, 1999, 105(3): 261-272.
[9] Kelemu S, Badel J L, Moreno C X,etal. Virulence spectrum of South American isolates ofColletotrichumgloeosporioideson selectedStylosanthesguianensisgenotypes. Plant Disease, 1996, 80: 1355-1358.
[10] Vinijsanun T, Irwin J A G, Cameron D F. Host range of three strains ofColletotrichumgloeosporioidesfrom tropical pasture legumes, and comparative histological studies of interactions between type B disease-producing strains andStylosanthesscabra(Non-host) andS.guianensis(Host). Australian Journal of Botany, 1987, 35(6): 655-677.
[11] Zhang W L, Guo Z F. Preliminary identification of the pathogen of stylo anthracnose disease and assessment of resistance ofStylosanthesspp..Guangdong Agricultural Sciences, 2007, 2: 51-54.
[12] Irwin J A G, Cameron D F. Two diseases inStylosanthesspp. caused byColletotrichumgloeosporioidesin Australia, and pathogenic specialization within one of the causal organisms. Crop and Pasture Science, 1978, 29(2): 305-317.
[13] Fang Z D. Research Methods of Plant Desease[M].Third edition. Bengjing: China Agriculture Press, 1998: 104-116.
[14] Stone J M, Heard J E, Asai T,etal. Simulation of fungal-mediated cell death by fumonisin B1 and selection of fumonisin B1-resistant (fbr)Arabidopsismutants. The Plant Cell, 2000, 12: 1811-1822.
[15] Wang S L, Liang P, Liu W B,etal. Cytological analysis of compatible interactions between rubber tree andOidiumheveae. Plant Protection, 2014, 40(3): 26-36.
[16] Connell R J, Bailey J A, Richmond D V. Cytology andphysiology of infection ofPhaseolusvulgarisbyColletotrichumlindemuthianum. Plant Pathology, 1985, 27: 75-98.
[17] Zhang J Z, Xu T. Cytological characteristics of the infection in different species, varieties and organs of persimmon byColletotrichumgloeosporioides. Mycosystema, 2005, 24(1): 116-122.
[18] Deising H B, Werner S, Wernitz M. The role of fungal appressoria in plant infection. Microbes Infect, 2000, 2(13): 1631-1641.
[19] Mendgen K, Deising H. Infection structures of fungal plant pathogens- a cytological and physiological evaluation. New Phytologist, 1993, 124(2): 193-213.
[20] Muhammed Z, Brlansky R H, Timmer L W. Intection of flower and vegetative tissues of Citrus byColletrichumacutatumandC.gloeosporioides. Mycological Society of America, 1996, 88(1): 121-128.
[21] Binyamini N, Nacel M S. Latent infection in Avocado fruit due toColletotrichumgloeosporioides. Phytopathology, 1971, 62: 592-594.
[22] Coates L M, Muirhead I F, Irwin J AG,etal. Initial infection processes byColletotrichumgloeosporioideson avocado fruit. Mycological Research, 1993, 97(11): 1363-1370.
[23] Ranathunge N P, Mongkolporn O, Ford R,etal.Colletotrichumtruncatumpathosystem onCapsicumspp: infection, colonization and defence mechanisms. Australasian Plant Pathology, 2012, 41(5): 463-473.
參考文獻:
[4] 馮淑芬, 李鳳娥, 何朝族. 筆花豆炭疽病菌生物學特性和流行條件研究. 熱帶作物學報, 1994, 15(1): 87-94.
[5] 易克賢, 鄭金龍, 習金根, 等. 柱花草炭疽病流行因子初探. 熱帶農業科學, 2014, 34(2): 74-78.
[11] 張偉麗, 郭振飛. 柱花草炭疽病菌初步鑒定及柱花草抗性分級研究. 廣東農業科學, 2007, 2: 51-54.
[13] 方中達. 植病研究方法[M]. 第三版. 北京: 中國農業出版社, 1998: 104-116.
[15] 萬三連, 梁鵬, 劉文波, 等. 橡膠樹與白粉病菌Oidiumheveae親和互作組織細胞學研究. 植物保護學報, 2014, 40(3): 26-36.
[17] 張敬澤, 徐同. 柿樹炭疽菌侵染不同柿樹種、品種和部位的細胞學特征. 菌物系統, 2005, 24(1): 116-122.
Microscopic observation ofStylosantheseinfected byColletotrichumgloeosporioides
YANG Mao-Xia, LIN Guo-Biao, CHEN Cai-Hong, WANG Hui, LUO Li-Juan*
AgronomyofHainanUniversity,HainanProvincialKeyLaboratoryofTropicalBiologicalResourcesSustainableUtilization,Haikou570228,China
In order to study the disease progress of Stylo anthrocnose and the inoculated plants reaction, the 6 weeks old ofStylosantheseguianensiscv. Reyan2 inoculated by the suspension liquid ofColletotrichumgloeosporioidesspores. Microscopic observations of the leaves revealed the infecting process and counting the disease index through plant symptoms. Spores germinated and a few spores formed appressorium at 4 h. Most of spores germinated at 6 h, and a large quantity of geminating spores formed appressorium at 12 h and began to form penetration pegs. The inoculated spores began to produce hyphae at 36 h, a lot of hyphae formed and staggered into a mesh on the surface of the leave at 48 h. The geminating spores began to form conidiophores at 60 h. Numerous of new spores began to produce at 72 h. Parts of inoculated spores have finished disease cycle in 72 h. In the same time, plants appeared disease symptom gradually: leaves color became yellow, the host cell membrane rupture at 24 h, brown necrosis appeared at 48 h, then dead line,sclerotinite,mycelium appeared on the leaves surface at 72 h. The host cell began to collapse at 96 h. Disease index is 54.52 at 8 days, 55% of inoculated plants dead at 11 days.
Stylosanthese; Stylo anthrocnose; infection; microstructure; desease symptom
10.11686/cyxb20150521
http://cyxb.lzu.edu.cn
2015-01-27;改回日期:2015-03-12
國家自然科學基金(31360575)和國家牧草產業技術體系熱帶牧草育種(CARS-35-03)資助。
楊茂霞(1989-),女,四川達州人,在讀碩士。E-mail:18789231651@163.com *通訊作者Corresponding author. E-mail:luoljd@126.com
楊茂霞, 林國彪, 陳彩虹, 王薈, 羅麗娟. 膠孢炭疽菌侵染柱花草葉片的顯微觀察. 草業學報, 2015, 24(5): 175-181.
Yang M X, Lin G B, Chen C H, Wang H, Luo L J. Microscopic observation of Stylo infected byColletotrichumgloeosporioides. Acta Prataculturae Sinica, 2015, 24(5): 175-181.
