基于ITS序列的梅花草屬Parnassia L.系統發育初步研究
2015-04-18 03:00:36柏夢焱李亞茹朱瓊潔葉成超劉碩然
大理大學學報 2015年12期
柏夢焱,李亞茹,朱瓊潔,葉成超,劉碩然
(大理大學農學與生物科學學院,云南大理 671003)
梅花草屬ParnassiaLinnaeus,1753根據P.palustris建立,是一類多年生草本植物,分布于北溫帶,主要分布于東亞和北美,極少數分布到歐洲及其他地區〔1-3〕。根據谷粹芝〔2〕、Gu和 Hultg?rd〔4〕的研究,中國分布有63種,其中49種為特有種,且集中分布于我國的西南部山區,該區是梅花草屬的現代分布中心和分化中心。
梅花草屬建立至今已有200多年的研究歷史。長期以來該屬的分類地位及其分類系統一直處于爭議狀態。根據形態學的研究,梅花草屬處于一個難以確定的系統位置,系統分類差異大,或作為虎耳草科Saxifragaceae的一個亞科,即梅花草亞科Parnassioideae;或獨立成一科,即梅花草科Parnassiaceae〔1,5-8〕。近幾年來隨著現代分子生物學的迅速發展,特別是PCR技術的運用及DNA測序方法的改進,大量分子序列數據的不斷累積,越來越多的基因片段被獲得并應用于系統發育的研究。特別是被子植物大系統的建立,關于梅花草屬的系統位置的觀點也趨于一致。Soltis等基于rbcL序列對廣義虎耳草科Saxifragaceae的研究表明梅花草屬與伯力木屬Brexia關系密切〔9〕;Savolainen等基于atpB-rbcL序列片段的研究認為梅花草屬與衛矛科Celastraceae、翅子藤科Hippocrateaceae和伯力木屬構成姊妹群〔10〕。APG的研究表明梅花草科包括梅花草屬和微型草屬Lepuropetalon兩屬,本科的系統關系與虎耳草科相距甚遠,而與衛矛科構成姊妹群,同屬于衛矛目〔11-12〕。顯然,分子系統學研究與以往主要根據形態學研究的結果有很大的差異。《中國植物志》以及西藏、四川、內蒙古、青海、秦嶺等各地方植物志的完成,使該屬及屬下分類單元的劃分和系統演化更為清晰。2005年吳丁對梅花草屬的分類系統學進行了詳細的研究,并提出了一個將梅花草屬植物劃分為12組的系統框架〔13〕,但是其在分類過程中,并不是所有的梅花草屬植物都運用了分子分析的手段。
在屬下、種間等系統學研究中,由于ITS片段具有引物通用、片段長短適中等特點,因此已得到廣泛應用〔14-15〕,同時Yang等〔16〕在2011年利用DNA條碼技術鑒別梅花草屬物種的研究中,表明在單序列分析中ITS片段具有最高的準確性,達86.7%;在多序列組合分析中,ITS+trnH-psbA序列片段具有最高準確性,達90%;因此,本研究將利用梅花草屬植物的ITS序列,通過三種不同的方法對所構建的發育樹之間的差異進行比較分析,并結合吳丁所創建的梅花草屬植物的現代屬下分類系統,補充分析未采用分子手段進行分類的幾個物種,使這個分類系統更具權威性,種間關系更加清晰,為梅花草屬植物系統進化及相關研究提供參考。
1 材料與方法
1.1 研究材料來源 研究所用的梅花草屬rDNAITS序列片段來源于NCBI網站(National Center for Biotechnology Information)。共收集到30條,包括ITS1、5.8S和ITS2,片段總長度661 bp,由于微型草Lepuropetalon spathulatum與內類群趨異率較小,且與其他外群相比具有最好的同源性,因此選擇微型草作為外群。
1.2 序列編輯及分析 首先序列矩陣上傳至MUSCLE(Multiple Sequence Comparison by Log-Expectation)進行排序〔17〕,再運用Bioedit7.0.9.0將排序后的矩陣手動完善序列〔18〕。
1.2.1 鄰接法 序列分析采用軟件MEGA5.1,依據最大復合可能性原則,分枝的可靠性評價使用Bootstrap分析啟發性搜索,1 000次重復取樣,各序列矩陣中的空位和缺失作刪除處理〔19〕。
1.2.2 最大似然法 序列分析上傳至ATGC:Phy-ML生物信息學在線平臺,選用GTR為替代模型,選用SPR&NNI來提高樹形的穩定性,“靴帶”分析設置為100次循環,各序列矩陣中的空位和缺失作刪除處理〔20〕。
1.2.3 貝葉斯推理法 序列分析采用Mrbayes-3.1.2軟件,設置進行1 000 000次取樣,每10代抽取一棵樹,共得到100 000棵樹,前25 000棵樹舍棄,剩余樹經估算得到的50%主要規則一致樹及后驗概率,取得0.004 4的分散頻率的平均方差,達到了小于0.01的標準。
2 結果
基于上述3種構樹方法,所得NJ樹、ML樹和BI樹如圖1、2、3所示。
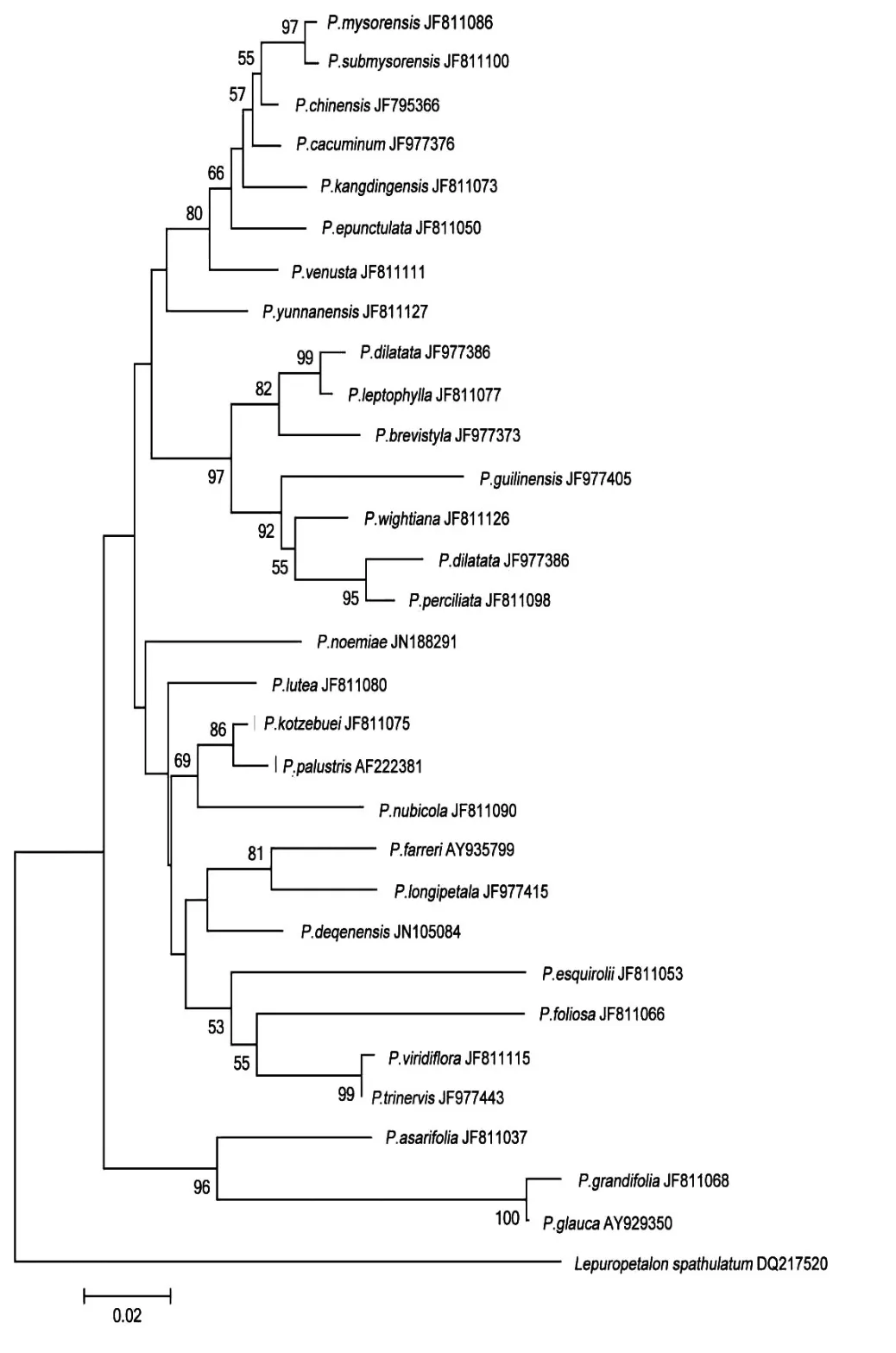
圖1 根據ITS序列構建的NJ樹
結果表明,NJ樹和ML樹的分枝的靴帶支持率整體不高,其中NJ樹少數分枝支持率達到100;ML樹少數分枝支持率達到100,最低為4,系統樹的中下部分枝支持率偏低。
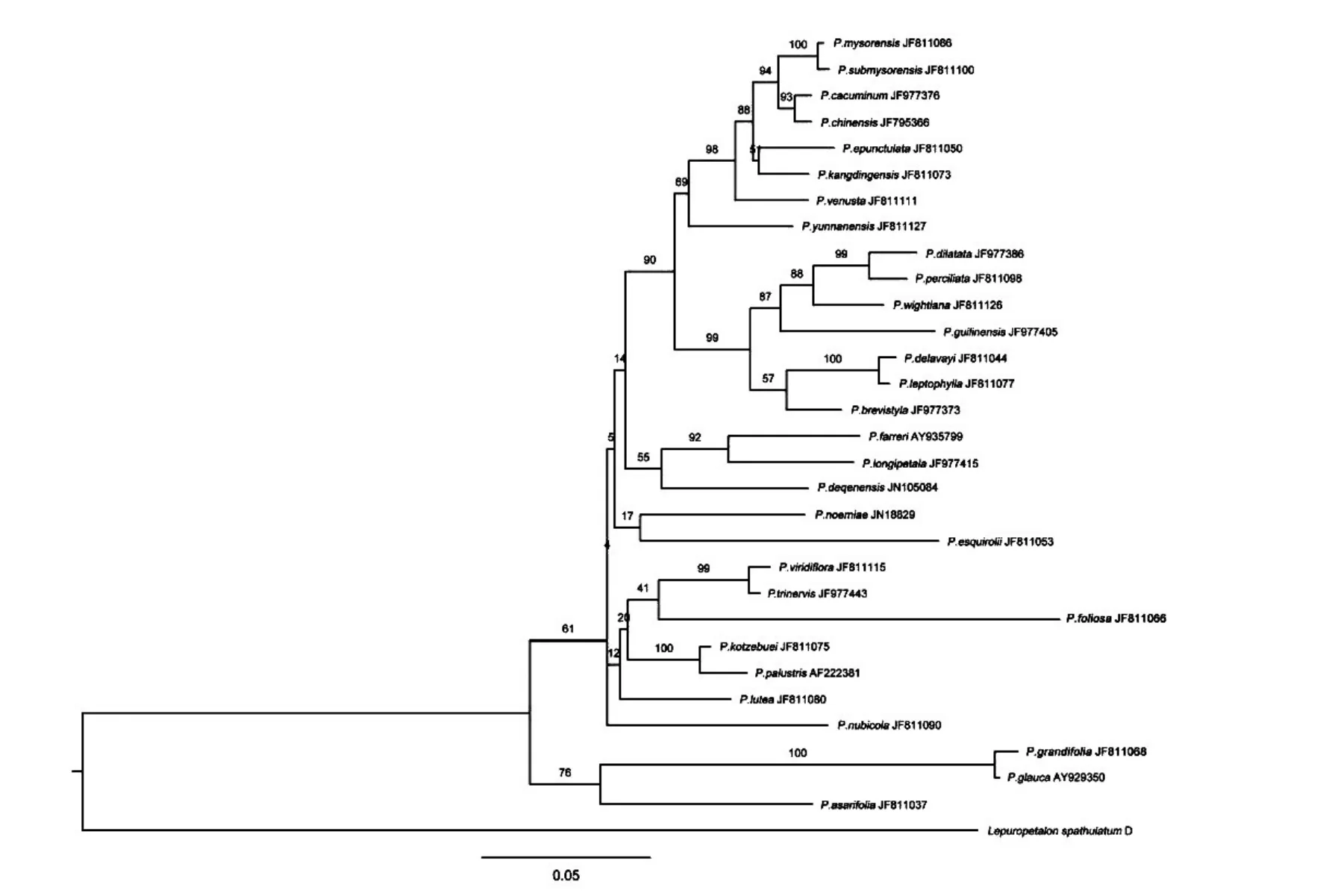
圖2 根據ITS序列構建的ML樹

圖3 根據ITS序列構建的BI樹
扁平三裂組Sect.nectarotrilobos的凹瓣梅花草P.mysorensis、近凹瓣梅花草P.submysorensis、高山梅花草P.cacuminum、中國梅花草P.chinensis、無斑梅花草P.epunctulata、妖媚梅花草P.venusta、康定梅花草P.kangdingensis、類三脈梅花草P.pusilla和柱形組Sect.saxifragastrum的云南梅花草P.yunnanensis8個種均位于樹的頂端,在BI樹的靴帶支持率均在96%以上,而在NJ樹和ML樹上的分枝支持率不高。
扁平五裂組Sect.allolobos的寬葉梅花草P.dilatata、厚葉梅花草P.perciliata、雞肫梅花草P.wightiana、桂林梅花草P.guilinensis4個種和長藥隔組Sect.xiphosandra的突隔梅花草P.delavayi、細裂梅花草P.leptophylla、短柱梅花草P.brevistyla3種構成姐妹群,且在BI樹的靴帶支持率達100%,但在NJ樹和ML樹上的分枝支持率不高。
外群微型草均處于系統樹的基部,長葉組Sect.longiloba的長枝梅花草P.grandifolia、灰梅花草P.glanca、細辛葉梅花草P.asarifola3個種均處于系統樹的亞基部,在BI樹的靴帶支持率達99%,但在NJ樹和ML樹上的分枝支持率不高。
現代梅花草屬植物的分類系統由吳丁在2005年建立,在建立此分類系統的過程中,采用了分子分析的手段,但是并沒將所有的物種囊括其中,在本文研究的30個梅花草物種中,有11個物種不在其梅花草屬屬下分類系統時所構建的系統發育樹中,如表1所示。基于3種不同構樹方法的比較研究,BI樹的拓撲結構較好的表現出了梅花草屬植物的親緣關系,因此本文依據此11個物種在BI樹的拓撲結構(如圖3)所處位置和現代分類系統所處位置進行比較分析。
從BI樹的拓撲結構來看,長葉組的云梅花草和無葉組的黃花梅花草構成一支,但是靴帶支持率低于50%。齒裂組的格德梅花草、長爪梅花草和柱形組的長瓣梅花草構成一個單系類群,支持率為99%,其中齒裂組的長爪梅花草和柱形組的長瓣梅花草形成一個小分枝,支持率為100%。

表1 通過系統發育學研究其分類地位的11個物種及其所屬組
由BI樹的拓撲結構可知,長枝組的長枝梅花草、灰梅花草、細辛葉梅花草處于系統樹的基部,構成一個單系類群,靴帶支持率為99%,其中長枝梅花草和灰梅花草關系更為密切,形成一小分枝,支持率為100%;梅花草組的克氏梅花草和梅花草構成一支,形成一個單系類群,支持率為100%;長藥隔組的細裂梅花草和突隔梅花草、短柱梅花草構成單系類群,支持率為90%,其中細裂梅花草和突隔梅花草關系更加密切,形成一小分枝,支持率為100%;扁平五裂組的寬葉梅花草和雞肫梅花草、厚葉梅花草、桂林梅花草構成一個單系類群,支持率為99%,其中寬葉梅花草和厚葉梅花草形成一小分枝,支持率為100%。
3 討論
3.1 3種不同構樹方法的比較分析 由鄰接法、最大似然法、貝葉斯推理法分析得到的系統發育樹在拓撲結構上保持較高的一致性,僅在部分端、部分枝表現出差異。NJ樹和ML樹的靴帶支持率不高,說明NJ樹和ML樹上各組的系統關系可信度較低。而BI樹分枝的后驗概率較高,大多數為100%,且基本都在95%以上,說明BI樹上各組的系統關系可信度較高。關于上述3種方法的適用性,基于本研究的數據貝葉斯推理法所得的系統發育樹獲得了最高的后驗概率,即拓撲結構表現出較高的穩定性。
對比上述三棵系統發育樹可以看出,在三棵樹中物種分布基本一致。但是其中有幾組的梅花草屬植物網結在一起,形成多個單枝,根據吳丁〔13〕提出的該屬可能的演化路線圖,唇形組的龍場梅花草更靠近基部;長葉組的云梅花草和無葉組的黃花梅花草關系密切;齒裂組、柱形組與扁平三裂組、扁平五裂組具有較近的親緣關系,對比三棵系統發育樹,BI樹的拓撲結構較好地表現出了梅花草屬植物的親緣關系。
3.2 分子手段對現代梅花草屬植物分類系統的論證從結果中可分析得出:齒裂組的長爪梅花草和柱形組的長瓣梅花草形成一個小分枝,支持率為100%,同時結合長瓣梅花的形態特征,其退化雄蕊不具備齒裂組的退化雄蕊扁平,低倒梯形,頂端具5~7齒的特點,而符合柱形組的退化雄蕊特點,因此長瓣梅花草仍屬于柱形組。而其他10種梅花草在BI樹上表現出來的拓撲結構均與梅花草屬現代分類系統一致。
3.3 梅花草屬植物的系統關系
3.3.1 三脈梅花草與綠花梅花草親緣關系分析在Yang等〔16〕的利用DNA條碼技術鑒定梅花草屬一文中,試圖推斷三脈梅花草可能是綠花梅花草的祖先,并通過兩者在進化樹上的位置進行說明,但是其樣本數量較小且樹枝穩定性不高,并未作出明確結論。本文通過Genbank搜集綠花梅花草和三脈梅花草的ITS片段,從其所構成的BI系統發育樹來看,綠花梅花草和三脈梅花草兩者同屬一枝,枝長相近,且支持率達到100%,拓撲結構穩定。此外,本文還選取了二者ITS序列片段進行同源性計算分析,兩者的同源性高達99.1%,ITS片段基本一致。因此通過本文采用的分子手段的分析,可以推斷三脈梅花草可能是綠花梅花草的祖先,但在形態學分類尚未作出改變前,我們仍需要更大的種群樣本進行分析以得到更穩定可靠的分子數據,進一步確定兩者的親緣關系。
3.3.2 類三脈梅花草與中國梅花草親緣關系分析在1941年“中國的梅花草”一文中類三脈梅花草被處理為中國梅花草的異名,但在現代梅花草屬下分類系統中將類三脈梅花草和中國梅花草同分在扁平三裂組〔13〕,以此本文試圖通過分子學手段推斷類三脈梅花草是否應處理為中國梅花草的異名。由于在Genbank中沒有找到類三脈梅花草的ITS序列,而在2008年康定梅花草歸并入類三脈梅花草〔21〕,則用康定梅花草的ITS序列代替類三脈梅花草的ITS序列與中國梅花草的ITS片段構建BI樹進行分析。但在樹中發現兩者并沒有處于一枝,但通過中國梅花草和康定梅花草的ITS片段同源性分析結果來看,相似性達到96.7%,兩者ITS片段非常相似,兩個結果相矛盾,結合康定梅花草被歸并入類三脈梅花草時是認為其子房位置存在連續性變異這一理由,所以只能推斷類三脈梅花草和中國梅花草具有非常相近的親緣關系,并不能證明為一個種,但本研究沒有到模式產地進行調查和核對模式標本,只能從分子角度得以論證。
3.3.3 高山梅花草與中國梅花草親緣關系分析觀察本研究所構建的BI樹可以發現,中國梅花草和高山梅花草同屬一枝,支持率為100%,且枝長基本相等;并且二者ITS片段同源性相似性達到99.06%,從分子角度分析二者具有非常近的親緣關系,筆者認為從分子學角度考慮,高山梅花草可以歸并入中國梅花草。
同時對二者的外觀形態進行了比較,形態特征極近似,二者僅有的區別在中國梅花草花瓣呈倒卵形,瓣片近圓形,基部有密集長流蘇狀毛,退化雄蕊裂片等長;而高山梅花草花瓣呈匙形,先端圓,基部有少許短流蘇狀毛,退化雄蕊裂片不等長。
對二者地理分布進行分析可知,中國梅花草分布于云南西北部、西藏東南部、四川、尼泊爾、不丹和緬甸;高山梅花草分布于我國青海東南部和四川西部和云南,而尚未查閱到其在國外有分布。通過在地圖上分析發現,兩者在國內分布范圍基本一致而中國梅花草在國外也有分布,如圖4。結合吳丁在2005年對梅花草屬植物地理分布的研究,可以推斷二者分化中心位于喜馬拉雅-橫斷山脈,二者可能擁有共同的祖先,且分化程度不高。
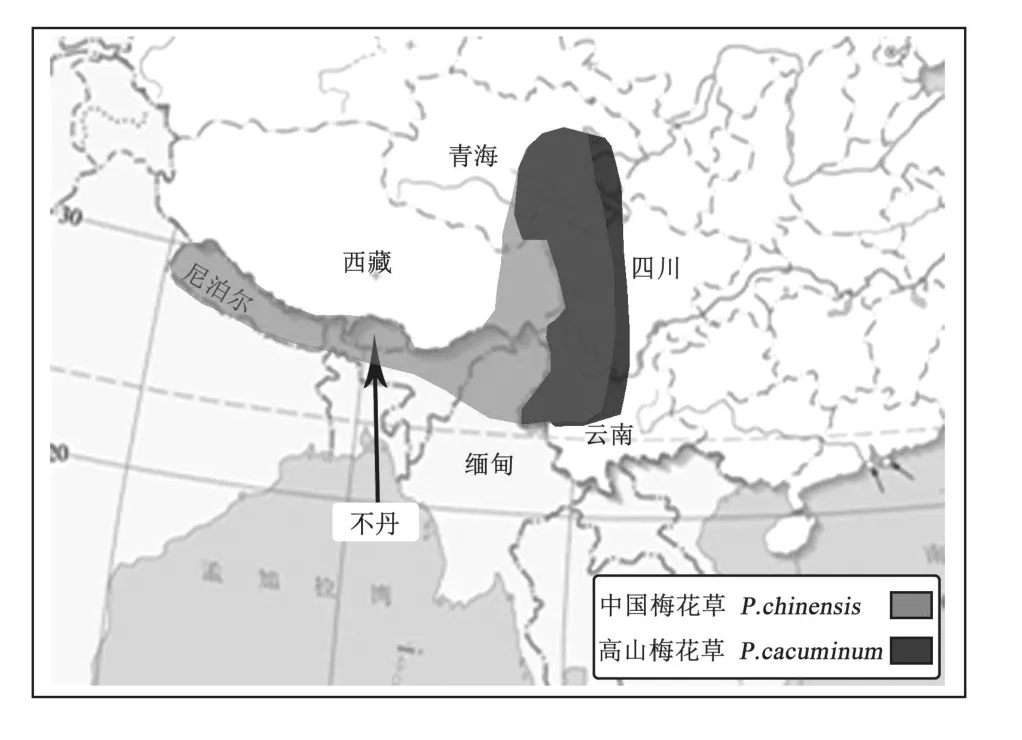
圖4 中國梅花草與高山梅花草的地理分布區域圖
綜合上述分子序列、形態和地理分布對二者親緣關系的分析,可以得出二者外觀形態可能存在連續性變異,可能為一個種,需要更多其他分子片段的序列分析進一步證實高山梅花草與中國梅花草的親緣關系。
〔1〕吳征鎰,路安民,湯彥承,等.中國被子植物科屬綜論〔M〕.北京:科學出版社,2003:357-385.
〔2〕谷粹芝.中國梅花草屬校訂〔J〕.植物研究,1987,7(1):1-61.
〔3〕 KORTA J.Anatomical analysis ofParnassia palustrisL.〔J〕.Acta Biologica Cracoviensia Series Botanica,1972,15(1):31-37.
〔4〕谷粹芝,李振宇,黃蜀瓊.中國植物志:52卷:第1分冊〔M〕.北京:科學出版社,2000:126-269.
〔5〕ARBER A.On the structure of the androecium inParnassiaand its bearing on the affinities of the genus〔J〕.Annals of Botany,1913,27(107):491-510.
〔6〕ARBER A.The anatomy of the stamens in certain Indian species ofParnassia〔J〕.Annals of Botany,1915,29(113):159-160.
〔7〕BENSEL C R,PALSER B F.Floral anatomy in the Saxifragaceae sensu lato.I.Introduction,Parnassioideae and Brexioideae〔J〕.American Journal of Botany,1975,62(7):661-675.
〔8〕谷粹芝,陸玲娣,黃淑美,等.中國植物志〔M〕.北京:科學出版社,1995:1-66.
〔9〕SOLTIS D E,PAMELA S S,MICHAEL T C,et al.rbcL sequence divergence and phylogenetic relationships in Saxifragaceae sensu lato〔J〕.Proceedings of the National Academy of Sciences,1990,87(12):4640-4644.
〔10〕SAVOLAINEN V,SPICHIGER R,MANEN J F.Polyphyletism of Celastrales deduced from a chloroplast noncoding DNA region 〔J〕.Molecular Phylogenetics and Evolution,1997,7(2):145-157.
〔11〕The Angiosperm Phylogeny Group.An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants:APG II〔J〕.Botanical Journal of the Linnean Society,2003,141(4):399-436.
〔12〕The Angiosperm Phylogeny Group.An ordinal classification for the families of flowering plants〔J〕.Annals of the Missouri Botanical Garden,1998,85(4):531-553.
〔13〕吳丁.梅花草屬植物的系統演化與生物地理〔D〕.昆明:中國科學院昆明植物研究所,2005.
〔14〕TABERLET P,GIELLY L,PAUTOU G,et al.Universal primers for amplification of three non-coding regions of chloroplast DNA〔J〕.Plant Molecular Biology,1991,17(5):1105-1109.
〔15〕田欣,李德銖.DNA序列在植物系統學研究中的應用〔J〕.云南植物研究,2002,24(2):170-184.
〔16〕YANG J B,WANG Y P,MOELLER M,et al.Applying plant DNA barcodes to identify species of Parnassia(Parnassiaceae)〔J〕.Molecular Ecology Resources,2012,12(2):267-275.
〔17〕Birney Research Group.Multiple Sequence Comparisonby LogEpecttion〔EB/OL〕.〔2005-08-26〕.http://www.ebi.ac.uk/Tools/msa/muscle.
〔18〕HALL T A.BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT〔R〕.Nucleic acids symposium series,1999,41:95-98.
〔19〕TAMURA K,PETERSON D,PETERSON N,et al.MEGA5:Molecular Evolutionary Genetics Analysis using Maximum Likelihood,Evolutionary Distance,and Maximum Parsimony Methods〔J〕.Molecular Biology and Evolution,2011,28(10):2731-2739.
〔20〕GUINDON S,DUFAYARD J F,LEFORT V,et al.New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies:Assessing the Performance of PhyML 3.0〔J〕.Systematic Biology,2010,59(3):7-21.
〔21〕吳丁,盧金梅,王紅.中國梅花草屬(梅花草科)一些種類的訂正〔J〕.云南植物研究,2008,30(6):657-661.
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
北京測繪(2020年12期)2020-12-29 01:33:58
電子制作(2018年18期)2018-11-14 01:48:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
