鞘氨醇單胞菌TP-3原生質體制備與再生的研究
2015-04-24 02:44:53周明明李曉雁陳彥梅黃海東
食品工業科技 2015年22期
周明明,李曉雁,陳 悅,陳彥梅,黃海東,*
(1.天津農學院農學與資源環境學院,天津300384;2.天津農學院食品科學與生物工程學院,天津300384)
鞘氨醇單胞菌TP-3原生質體制備與再生的研究
周明明1,李曉雁2,陳 悅2,陳彥梅1,黃海東1,*
(1.天津農學院農學與資源環境學院,天津300384;2.天津農學院食品科學與生物工程學院,天津300384)
研究了EDTA預處理、菌齡、溶菌酶濃度、酶解溫度及酶解時間對鞘氨醇單胞菌TP-3原生質體制備與再生的影響。結果表明:10 mmol/L EDTA對菌體進行預處理,菌齡18 h,酶濃度60 μg/mL,酶解溫度32℃,酶解時間30 min時,菌株原生質體的形成率可達96.1%,再生率達32.2%。本文為進行原生質體融合選育優良微生物菌種奠定了基礎。
鞘氨醇單胞菌,原生質體,影響因素
食品膠是一類天然高分子聚合物的統稱,這類物質充分水化后能形成黏稠、滑膩或凝膠狀的高分子水溶膠,在食品加工中可以起到增稠、乳化、膠凝等作用[1-2]。食品膠按來源可分為植物膠、動物膠、微生物膠、海藻膠和化學改性膠,其中微生物膠性能優良、用途廣泛、資源取之不竭,與其他來源的食品膠相比,生產周期短、不受氣候、地理環境和自然災害的影響,其產品研發和應用日益受到重視[3-6]。
鞘氨醇單胞菌是合成微生物膠的重要微生物資源,該菌屬的菌株能合成包括結冷膠在內的一類生物聚合物,統稱為鞘氨醇膠。菌株TP-3是從土壤中分離到的一株鞘氨醇單胞菌屬的新種Sphingomonas sanxanigenens,其發酵合成的產物命名為鞘氨醇膠Ss,此膠具有優越的增稠性和剪切稀釋性,在含鈣離子的水溶液中低濃度即可形成凝膠[7-8]。將菌株TP-3的發酵液pH調至3.0左右,鞘氨醇膠Ss即可沉淀出來,與其他鞘氨醇膠所采用的有機溶劑沉淀法提取相比,酸沉提取能夠顯著地降低生產成本。本文對菌株TP-3的原生質體制備與再生條件進行了研究,為利用原生質體融合選育優良菌株,進一步提高該菌株的產物合成量和生產性能奠定基礎。
1 材料與方法
1.1 材料與儀器
菌種S.sanxanigenens TP-3 由本實驗室保藏;固體培養基(g/L) 蔗糖15.0,蛋白胨5.0,牛肉膏3.0,酵母膏3.0,瓊脂粉15.0,pH7.0;液體培養基(g/L) 蔗糖15.0,蛋白胨2.5,磷酸氫二鉀2.5,磷酸氫二銨1.5,硫酸鎂0.1,酵母粉1.5,pH7.0;補充基本培養基(BNK)(g/L) 蔗糖181.0,蛋白胨5.0,牛肉膏3.0,瓊脂粉20.0,pH7.0;再生補充培養基(BZNK)(g/L) BNK配方中瓊脂改為8.0;磷酸緩沖液 0.1 mol/L磷酸氫二鈉,0.1 mol/L磷酸二氫鈉,pH7.0;高滲緩沖液0.1 mol/L磷酸氫二鈉,0.1 mol/L磷酸二氫鈉,0.8 mol/L甘露醇,pH7.0;原生質體穩定液(SMM) 0.5 mol/L蔗糖,20 mol/L MgCl2,0.02 mol/L順丁烯二酸,pH6.5;溶菌酶 用SMM溶液配制,終濃度0.5 mg/mL,過濾除菌備用;磷酸氫二鉀、磷酸氫二銨、硫酸鎂、磷酸氫二鈉、磷酸二氫鈉、甘露醇、MgCl2、順丁烯二酸等 均為分析純;酚藏花紅染色劑 購自Sigma公司。
KYC-100C搖床 上海福瑪公司;TGL16A臺式高速離心機 潮南凱達公司;WH-2微型旋渦混合儀 上海滬西公司;BA410熒光顯微鏡 廈門Motic公司;NDJ-1旋轉粘度計 上海精密公司;STARTER 2100實驗室pH計 奧豪斯儀器(上海)有限公司;DHG-9078A型電熱恒溫鼓風烘干箱 上海精宏實驗設備有限公司;MCO-15A型恒溫培養箱 上海市躍進醫療器械一廠;BCM-1000A生物潔凈工作臺 蘇州安泰潔凈有限公司;FA2004電子天平 上海精密天平;電子顯微鏡 日本Philips EM400-ST。
1.2 實驗方法
1.2.1 菌種活化 取保藏的TP-3菌種劃線接種到平板上,30℃恒溫培養72 h,挑取單菌落在試管斜面上蛇形劃線,30℃恒溫培養72 h。
1.2.2 菌株發酵曲線測定 取5 mL無菌水將斜面試管中的菌體沖洗下來,按10%的接種量將其接種到液體培養基中,30℃、180 r/min搖床培養72 h。每隔6 h取一次樣,并測定發酵液的pH、粘度、菌數和鞘氨醇膠Ss產量。
1.2.3 發酵參數的測定 使用校準后的pH計測定發酵液的酸堿度;在6 r/min的條件下,使用3號轉子測定發酵液的粘度;發酵液無菌取樣后,用無菌水進行梯度稀釋,取0.1 mL稀釋樣涂布于固體平板培養基上,30℃恒溫培養3 d后進行菌落計數,計算發酵的菌數指標;發酵過程中取樣,加入1 mol/L的鹽酸調節發酵液pH至3.0,沉淀的產物過濾后80℃恒溫干燥至恒重,稱重得發酵產物量。
1.2.4 原生質體的制備 菌株TP-3液體培養至對數生長前期,取適量菌液在8000 r/min條件下離心5 min,棄上清液,用磷酸緩沖液洗滌菌體細胞。將菌體懸浮于適量的SMM緩沖液中,使溶液中細胞濃度約為108~109CFU/mL。取0.1 mL菌懸液,用無菌水稀釋到10-5、10-6、10-7,涂布平板培養基,30℃培養4 d后得菌落數A(菌落總數)。取剩余的菌懸液,在其中加入溶菌酶,混合均勻后于37℃保溫30 min,4000 r/min離心10 min,棄上清液,用高滲緩沖液洗滌除酶,取菌液0.1 mL,用無菌水稀釋到10-2、10-3、10-4,涂布平板培養基,進行剩余菌數B(未被酶裂解的剩余細胞)的測定。
1.2.5 原生質體的再生 取0.1 mL酶解后的原生質體懸液,用SMM緩沖液稀釋到10-3、10-4、10-5,各取1 mL加入底層BNK培養基的中央,再倒入上層BZNK培養基,混勻,30℃培養4~5 d后計數,將菌落數計為C(再生菌落數)。原生質體形成率與再生率的計算公式如下:原生質體形成率(%)=(A-B)/A×100,原生質體再生率(%)=(C-B)/(A-B)×100。
1.2.6 原生質體制備與再生的影響因素
1.2.6.1 EDTA預處理對原生質體制備與再生的影響 酶解前,在菌齡18 h,酶濃度100 μg/mL,酶解溫度37℃,酶解時間30 min的固定條件下,加入2~14 mmol/L濃度梯度的EDTA對菌懸液進行預處理,測定原生質體形成率和原生質體再生率。
1.2.6.2 青霉素濃度對原生質體制備與再生的影響用10 mmol/L EDTA對菌體進行預處理,菌齡為18 h,酶濃度100 μg/mL,溫度37℃,酶解時間30 min的固定條件下,加入0~100 μg/mL的青霉素預處理2 h,測定原生質體形成率和原生質體再生率。
1.2.6.3 菌齡對原生質體制備與再生的影響 依據1.2.2發酵曲線中的結果,用10 mmol/L EDTA對菌體進行預處理,酶濃度100 μg/mL,溫度37℃,酶解時間30 min的固定條件下,選擇6~36 h的細菌培養物制備原生質體,測定原生質體形成率和原生質體再生率。
1.2.6.4 溶菌酶濃度對原生質體制備與再生的影響用10 mmol/L EDTA對菌體進行預處理,菌齡18 h,溫度37℃,酶解時間30 min的固定條件下,取10~120 μg/mL的溶菌酶制備原生質體,測定原生質體形成率和原生質體再生率。
1.2.6.5 酶解溫度對原生質體制備與再生的影響 用10 mmol/L EDTA對菌體進行預處理,菌齡18 h,酶濃度100 μg/mL,酶解時間30 min的固定條件下,選擇17~47℃范圍的酶解溫度制備原生質體,測定原生質體形成率和原生質體再生率。
1.2.6.6 酶解時間對原生質體制備與再生的影響 用10 mmol/L EDTA對菌體進行預處理,菌齡18 h,酶濃度100 μg/mL,酶解溫度37℃的固定條件下,酶解時間為10~70 min制備原生質體,測定原生質體形成率和原生質體再生率。
1.2.7 原生質體制備條件的驗證 在上述原生質體各制備因素的基礎上,得到原生質體制備與再生的最佳條件,進行各影響因素的綜合實驗,得出菌株原生質體的最高形成率和再生率。
1.2.8 菌體細胞與原生質體的觀察 用接種環取菌懸液于干凈的載玻片上,晾干,加結晶紫染液進行染色,用水沖洗并晾干,在顯微鏡下進行觀察。細菌電子顯微鏡觀察方法見參考文獻[9],配制0.01%的酚藏花紅染色劑[10],在原生質體溶液中按1∶1的比例加入染色劑,將混合溶液滴于載玻片上,加蓋玻片,在熒光顯微鏡下進行觀察。
1.3 數據處理
實驗進行三次重復,應用軟件Origin 6.1和Excel 2003對得出的數據進行分析處理。
2 結果與分析
2.1 菌株TP-3的發酵曲線
在實驗菌株發酵培養的過程中,進行pH、發酵液粘度、菌數和鞘氨醇膠Ss產物量的測定,結果如圖1所示。從細胞生長曲線可以看出,菌株TP-3在培養8 h后進入對數生長期,46 h進入穩定期,此時菌數達1.5×1010CFU/mL;產物鞘氨醇膠Ss的合成曲線與細胞生長曲線近乎平行,在60 h達到最高24.1 g/L,結晶紫染色后鏡檢發現,在發酵培養的全部階段均能觀察到纖毛狀的產物包裹在細胞周圍,說明鞘氨醇膠Ss是一種初級代謝產物;發酵培養在32 h之前,發酵液的pH基本穩定,隨著產物合成量的增加,發酵液粘度逐漸增加,產物鞘氨醇膠Ss屬于酸性多糖,因此32 h之后發酵液的pH略有降低。由于整個發酵周期中,胞外均存在高分子多糖,不利于原生質體的制備,提示應該選擇發酵液粘度低、產物合成少的發酵階段,進行原生質體制備的研究。

圖1 菌株TP-3的發酵曲線Fig.1 Fermentation growth curve of strain TP-3
2.2 原生質體的制備與再生
2.2.1 EDTA預處理對原生質體制備與再生的影響鞘氨醇單胞菌TP-3為革蘭氏陰性細菌,結構和成分較革蘭氏陽性菌復雜,特別是細胞外壁中的脂多糖及多糖類,會影響溶菌酶的作用。Minghua D等發現,制備原生質體時,在添加溶菌酶之前用EDTA預處理,可以提高原生質體的形成率[11]。本實驗用EDTA對菌懸液進行預處理,結果如圖2所示。

圖2 EDTA預處理對原生質體形成率與再生率的影響Fig.2 Effect of EDTA pretreatment on protoplast formation and regeneration of strain TP-3
隨著EDTA濃度的增大,原生質體的形成率在10 mmol/L之前逐漸提高;原生質體的再生率則相反,呈下降趨勢。以原生質體形成率與再生率的乘積最大,作為判斷原生質體制備最佳條件的依據[12],以此標準,選擇10 mmol/L的濃度作為EDTA預處理的最佳濃度。
2.2.2 青霉素濃度對原生質體制備與再生的影響在菌株TP-3液體培養的過程中加入一定量的青霉素,可以在一定程度上提高原生質體的形成率(圖3)。與抑制革蘭氏陽性菌的機理不同,青霉素可以與革蘭氏陰性菌細胞膜上的青霉素結合蛋白PBPS發生結合,從而干擾細胞壁代謝,并造成細胞形變[13]。

圖3 青霉素濃度對原生質體形成率與再生率的影響Fig.3 Effect of concentration of penicillin on protoplast formation and regeneration of strain TP-3
從實驗結果可以看出,青霉素的加入明顯降低了原生質體再生率;在濃度低于20 μg/mL時,使用青霉素有利于原生質體的形成,但濃度繼續升高后,由于細胞存活率下降,實驗過程中的剩余細胞數量迅速下降,原生質體形成率也相應降低。
2.2.3 菌齡對原生質體制備與再生的影響 菌齡對原生質體的制備與再生很重要,處于對數生長期的細胞生理狀態相對一致,細胞分裂迅速,對酶的敏感性強,易于原生質體化和再生[14]。菌齡對原生質體制備與再生的影響,結果如圖4所示。
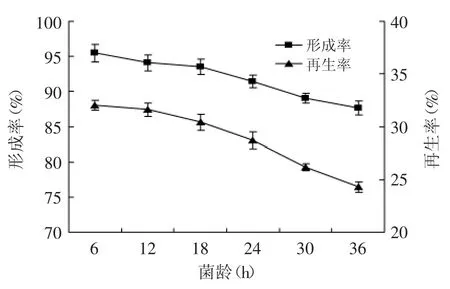
圖4 菌齡對原生質體形成率與再生率的影響Fig.4 Effect of cell age on protoplast formation and regeneration of strain TP-3
原生質體的形成率和再生率都隨著菌齡的增加而下降。說明隨著發酵時間的延長,菌體細胞周圍的多糖產物逐漸增多,影響溶菌酶的作用效果;但菌齡過短時細胞數量偏低,不利于后續的原生質體融合研究,因此選擇菌齡為18 h作為原生質體制備與再生的最佳發酵時間。
2.2.4 溶菌酶濃度對原生質體制備與再生的影響經EDTA預處理后,實驗菌株細胞壁表面的脂多糖層的Ca2+、Mg2被螯合去除,破壞了脂多糖的結構,使溶菌酶可以作用于細胞壁的肽聚糖[15]。用不同濃度的溶解酶進行原生質體的制備與再生實驗,結果如圖5所示。
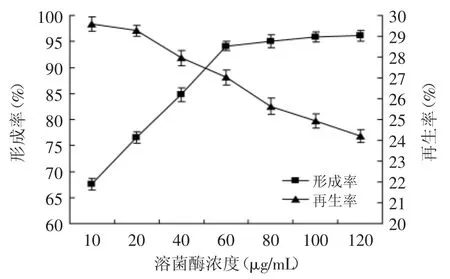
圖5 酶濃度對原生質體形成率與再生率的影響Fig.5 Effect of concentration of lysozyme on protoplast formation and regeneration of strainTP-3
隨著溶菌酶量的增加,原生質體形成率呈上升趨勢,由于細胞壁破壞影響菌體活性,再生率呈下降趨勢。溶菌酶濃度在10~60 μg/mL范圍內,原生質體形成率迅速增長,繼續提高溶菌酶濃度,原生質體形成率增加不明顯,但原生質體再生率迅速下降。原因可能是隨著酶濃度的增加,酶不僅酶解了細胞壁,還對早期形成的原生質體細胞膜造成了損傷,再生率下降[12]。依據原生質體形成率與再生率乘積最大原則[12],選擇溶菌酶濃度為60 μg/mL作為原生質體制備與再生的最佳條件。
2.2.5 酶解溫度對原生質體制備與再生的影響 原生質體制備時,低于酶解最適溫度時,酶活性低,酶解時間長;溫度高于酶解最適溫度時,酶蛋白逐漸變性而使酶失活,不利于原生質體的形成[16]。本文對不同的酶解溫度進行了實驗,結果如圖6所示。

圖6 酶解溫度對原生質體形成率與再生率的影響Fig.6 Effect of temperatures of lysozyme on protoplast formation and regeneration of strainTP-3
隨著溫度的升高,原生質體的形成率呈上升趨勢,再生率則持續下降,酶解溫度達到47℃時,再生率降為0。隨著溫度的升高溶菌酶活性增強,因此原生質體形成率增高,再生率的下降一方面是由于溶菌酶對細胞的損傷,另一方面,菌株TP-3的最適發酵溫度為28~30℃,溫度過高對菌體本身的內酶系統造成破壞,使原生質體無法再生[17]。選擇32℃為最佳酶解溫度。
2.2.6 酶解時間對原生質體制備與再生的影響 溶菌酶作用時間也是影響原生質體制備的重要因素,酶解時間不足,細胞不能完全去壁;酶解時間過長,會使早期釋放的原生質體膜受到傷害,使原生質體破碎或再生能力減弱[18]。本文對不同的酶解時間進行了實驗,結果如圖7所示。
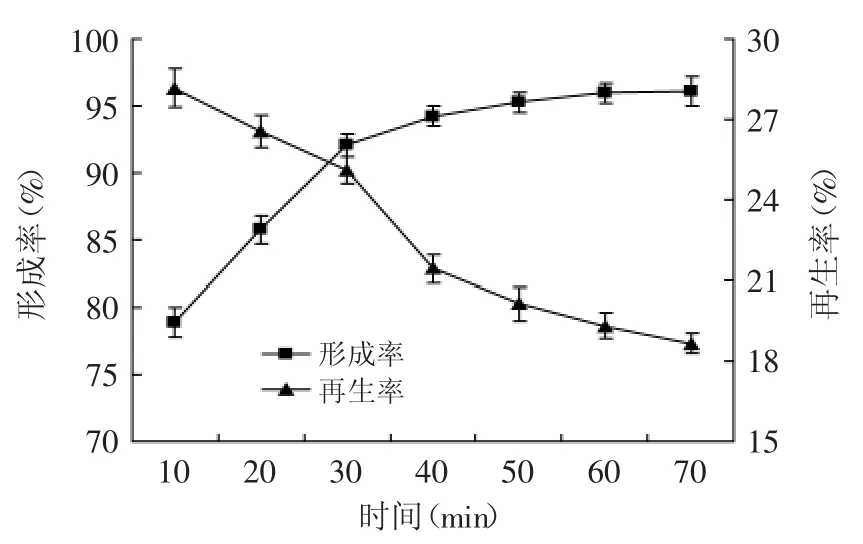
圖7 酶解時間對原生質體形成率與再生率的影響Fig.7 Effect of operational times of lysozyme on protoplast formation and regeneration of strainTP-3
在酶作用10~30 min之間,原生質體的形成率隨酶解時間延長而迅速提高,之后增速緩慢,再生率則呈下降趨勢。酶解30 min后,原生質體再生率的迅速下降,說明過長的酶解時間影響了原生質體的活性,因此選擇30 min作為原生質體形成與再生的最佳酶解時間。

圖8 TP-3菌體細胞(7000×)Fig.8 Cell of strainTP-3(7000×)

圖9 菌株TP-3的原生質體(1000×)Fig.9 Protoplast of strainTP-3(1000×)
2.3 原生質體制備條件的驗證
根據上述實驗結果,得到原生質體制備與再生的最佳條件為:10 mmol/L EDTA預處理,菌齡18 h,溶菌酶濃度60 μg/mL,酶解溫度32℃,酶解時間30 min。在該條件下進行驗證實驗,結果表明原生質體形成率和再生率分別達到96.1%和32.2%。對原生質體制備前后的菌體形態進行觀察,圖8為菌體細胞的電子顯微鏡觀察結果,可以看出菌株TP-3的細胞為桿狀,大小為0.6~0.9 μm×1.5~2.0 μm,細胞周圍可以觀察到明顯的纖毛狀產物,鞘氨醇膠Ss。菌株TP-3的原生質體進行酚藏花紅染色,在熒光顯微鏡下觀察(圖9),大多數菌體呈類球狀,直徑0.9~1.2 μm,原生質體周圍仍然能觀察到多糖產物。
3 結論與討論
本文對菌株TP-3的原生質體制備條件進行了優化,發現EDTA預處理、菌齡、溶菌酶濃度、酶解溫度和時間等因素對原生質體的形成率與再生率影響明顯。由于菌株TP-3外包裹著產物Ss膠,影響了溶菌酶對菌株的作用,進而影響原生質體的制備。通過優化實驗得出原生質體制備與再生的最佳條件:10 mmol/L EDTA預處理,菌齡18 h,酶濃度60 μg/mL,酶解溫度32℃,酶解時間30 min,在此條件下菌株原生質體的形成率可達96.1%,再生率達32.2%。菌株TP-3是鞘氨醇單胞菌屬的新種,該菌株的原生質體制備與再生條件的研究還未見報道。實驗中發現加入20 μg/mL濃度以下的青霉素,可干擾細胞壁合成,原生質體形成率有所提高,但使用青霉素后細菌存活率降低,不利于后期原生質體融合育種,因此選擇不加青霉素進行預處理。菌株TP-3原生質體周圍仍可以觀察到多糖產物的分泌,進一步說明鞘氨醇膠Ss合成后連接在細胞膜的脂載體上,并不分泌到細胞壁外。
Ss是鞘氨醇膠家族的一個新品種,增加產物量、提高產品粘度、降低膠凝臨界濃度等諸多研究工作有待開展。對菌株進行傳統誘變,獲得目的性狀提高的突變株后,通過多輪遞推式原生質體融合,可以快速得到發酵性狀更優良的菌株。原生質體的制備和再生是決定融合效率的關鍵因素,本文為菌株TP-3進行原生質體融合及基因組改組等育種工作奠定了基礎。
[1]Vasiliki E,Maria M,Ioanna M,et al.Compression of gellan gels.PartⅡ:effect of sugars[J].Food Hydrocoll,2010,24(4):392-397.
[2]胡國華.功能性食品膠[M].北京:化學工業出版社,2003:1-22.
[3]Prajapati VD,Jani GK,Zala BS,et al.An insight into the emerging exopolysaccharide gellan gum as a novel polymer[J]. Carbohydr Polym,2013,93(2):670-678.
[4]Bradbeer JF,Hancocks R,Spyropoulos F,et al.Self-structuring foods based on acid-sensitive low and high acyl mixed gellan systems to impact on satiety[J].Food Hydrocoll,2014,35(100):522-530.
[5]張偉,沈年漢,陳秀芳,等.幾種微生物多糖的特性及在食品工業中的應用[J].食品工業科技,2010,31(6):358-360.
[6]朱桂蘭,童群義.微生物多糖的研究進展[J].食品工業科技,2012,33(6):444-448.
[7]Haidong H,Wei W,Ting M,et al.Sphingomonas sanxanigenens sp.nov.,isolated from soil[J].Int J Syst Evol Microbiol,2009,59(4):719-723.
[8]Huang HD,Wang W,Ma T,et al.Analysis of molecular composition and properties of a novel biopolymer[J].Chem J Chinese Univ,2011,30:324-327.
[9]胡冰.生物電子顯微鏡技術[M].南京:南京農業大學出版社,2004:99-101.
[10]龍建友,唐世榮,吳文君.原生質體融合技術對秦嶺霉素產量提高的影響[J].中國農業科學,2007,40(7):1416-1421.
[11]Minghua D,Sara Z,Thomas R,et al.Visualization of protoplast fusion and quantitation of recombination in fused protoplasts of auxotrophic strains of Escherichia coli.[J].Metabolic Engineering,2005,7(1):45-52.
[12]岑沛霖.工業微生物學[M].北京:化學工業出版社,2001:335-344.
[13]張鳳凱,金少鴻.β-內酰胺類抗生素的作用靶位-青霉素結合蛋白[J].國外醫藥:抗生素分冊,2000,21(3):107-110.
[14]曾獻春,孟冬麗.乳酸菌原生質體制備與再生研究[J].食品科學,2006,27(10):269-272.
[15]施巧琴,吳松剛.工業微生物育種學[M].北京:科學出版社,2003:302-303.
[16]韓光杰,孫俊,李傳明,等.蘇云金芽孢桿菌原生質體的制備及大質粒轉化[J].江蘇農業科學,2014,42(11):37-40.
[17]甘邱鋒,張曉琳,王潔穎,等.刺糖多孢菌原生質體制備與再生條件優化[J].中國抗生素雜志,2011,36(1):18-24.
[18]宋愛環,李紅葉,劉小紅.指狀青霉(Penicillium digitatum)原生質體制備和再生條件[J].農業生物技術學報,2004,12(2):197-201.
Conditions for protoplast preparation and regeneration by Sphingomonas sanxanigenens TP-3
ZHOU Ming-ming1,LI Xiao-yan2,CHEN Yue2,CHEN Yan-mei1,HUANG Hai-dong1,*
(1.College of Agronomy and Resources&Environment,Tianjin Agricultural University,Tianjin 300384,China;2.College of Food Science and Biotechnology,Tianjin Agricultural University,Tianjin 300384,China)
The effect of EDTA pretreatment,cell age and the concentration,temperature and work time of lysozyme on protoplast formation and regeneration by Sphingomonas sanxanigenens TP-3 were studied.The results showed that when cell age was 18 h,pretreated by 10 mmol/L EDTA,using 60 μg/mL of lysozyme,enzymolysis at 32℃for 30 min,the protoplast formation rate and regeneration rate respectively reached 96.1%and 32.2%. This work lay the foundation for protoplast fusion to breed good microbial strain.
Sphingomonas;protoplast;influence factors
TS201.1
A
1002-0306(2015)22-0184-05
10.13386/j.issn1002-0306.2015.22.030
2015-03-19
周明明(1990-),女,碩士研究生,主要從事微生物多糖方面的研究,E-mail:zmmydx@126.com。
*通訊作者:黃海東(1972-),男,博士,教授,主要從事資源細菌及工程方面的研究,E-mail:hhaidong@126.com。
國家自然科學基金項目(31571790);天津市大學生創新創業項目(201410061062)。
