超表達葡萄糖脫氫酶促進大腸桿菌生長
2015-04-24 02:44:54裴慶慧葛喜珍田平芳
食品工業科技 2015年22期
關鍵詞:生長
鄭 躍,裴慶慧,葛喜珍,田平芳,*
(1.北京化工大學生命科學與技術學院,北京100029;2.北京聯合大學生物化學工程學院,北京100023)
超表達葡萄糖脫氫酶促進大腸桿菌生長
鄭 躍1,裴慶慧2,葛喜珍2,田平芳1,*
(1.北京化工大學生命科學與技術學院,北京100029;2.北京聯合大學生物化學工程學院,北京100023)
通過PCR擴增了肺炎克雷伯氏菌(Klebsiella pneumoniae)和大腸桿菌(Escherichia coli)各自的葡萄糖脫氫酶基因KPgdh和ECgdh,構建了表達載體pET-28a-KPgdh和pET-28a-ECgdh,轉化大腸桿菌E.coli BL21,獲得了重組菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)。經IPTG誘導,聚丙烯酰胺凝膠電泳顯示重組菌實現了葡萄糖脫氫酶的高效表達,且葡萄糖脫氫酶活性分別是空質粒對照菌E.coli BL21(pET-28a)的8.5倍和12倍。重組菌的生長速度快于空質粒對照菌E.coli BL21(pET-28a),并與原代菌E.coli BL21持平。如果扣除質粒復制造成的代謝負荷,過表達葡萄糖脫氫酶促進了大腸桿菌的生長。
大腸桿菌,肺炎克雷伯氏菌,葡萄糖脫氫酶,超表達,細胞生長
葡萄糖脫氫酶(Glucose dehydrogenase)廣泛存在于生物體,如牛心臟、大腸桿菌、洋蔥伯克霍爾德菌(Burkholderiacepacia)、弱氧化葡糖桿菌(Gluconobacter suboxydans)、枯草芽孢桿菌(Bacillus subtilis)等[1-4]。葡萄糖脫氫酶在食品領域有重要的作用,它參與磷酸戊糖途徑,以NAD+(NADP+)為輔基,特異催化β-D-葡萄糖生成葡萄糖酸[5-6],進而與鈣鹽反應生成可促進人體補鈣吸收的葡萄糖酸鈣[7]。D-核糖是一種核苷酸衍生物,可用于生產調味品、香精等食品添加劑,還具有保健的功效[8]。葡萄糖脫氫酶是合成D-核糖的限速酶,其活力與D-核糖的產量密切相關[9]。此外葡萄糖脫氫酶和果糖轉移酶組成的系統能夠有效降低葡萄糖的含量,從而大大提高低聚果糖的純度[10]。肺炎克雷伯氏菌雖有一定致病性,但能高產3-羥基丙酸、1,3-丙二醇、2,3-丁二醇等高值化合物,是有潛力的工業微生物[11];肺炎克雷伯氏菌能利用甘油、葡萄糖等多種碳源快速生長,較大腸桿菌而言,其生長更旺盛,碳源代謝能力更強。研究其葡萄糖脫氫酶有望獲得生長迅速的工程菌。
本研究以大腸桿菌E.coli BL21為宿主,以pET-28a為載體,由IPTG誘導T7啟動子來表達不同來源的GDH基因,測定催化活性及生長速度,旨在探索超表達葡萄糖脫氫酶對菌體生長的促進作用,且研究了葡萄糖脫氫酶異源表達對其活性是否存在影響,為葡萄糖脫氫酶的開發及其在食品領域的應用奠定基礎。
1 材料與方法
1.1 材料與儀器
大腸桿菌E.coli DH5α、肺炎克雷伯氏菌K.pneumoniae MGH78578//大腸桿菌E.coli BL 21、質粒pET-28a 由本實驗室保存;限制性內切酶EcoRⅠ、SacⅠ和HindⅢ TaKaRa公司;T4 DNA連接酶 NEB公司;Taq DNA聚合酶、蛋白質分子質量標記、DNA分子量Marker、硫酸卡那霉素 北京全新拓達公司;質粒提取試劑盒、DNA產物回收試劑盒以及切膠回收試劑盒 北京博邁德生物公司。
JA1003B型電子天平 上海越平科學儀器有限公司;TGL-16B型高速離心機 上海安亭科學儀器廠;LRH70型培養箱 上海一恒科技有限公司;GT9612型PCR儀 BIOGENER;H2O3-PRO金屬浴 金銀杏生物科技有限公司;THZ-C型恒溫振蕩器 太倉市儀器設備廠;10011624型分光光度計 上海奧普勒儀器有限公司;FluorChemTM5500凝膠成像儀 安萊(美國);DYY-8C型電泳儀 北京市六一儀器廠;SCL-10A型高效液相色譜 島津(日本);TY-80A/S型脫色搖床 江蘇榮華儀器制造有限公司。
1.2 培養基的配制
LB液體培養基:蛋白胨10 g/L,酵母粉5 g/L,氯化鈉10 g/L,pH7.0;LB固體培養基需加入瓊脂15 g/L;M9發酵培養基:5×M9鹽溶液 200 mL,1 mol/L MgSO4·7H2O 2 mL,20%葡萄糖20 mL,1 mol/L CaCl20.1 mL,添加去離子水至1000 mL;5×M9鹽溶液(200 mL):Na2HPO4·12H2O 17.1 g,KH2PO43 g,NaCl 0.5 g,NH4Cl 1 g。
1.3 基因克隆與表達
1.3.1 葡萄糖脫氫酶基因的克隆 根據GenBank公布的肺炎克雷伯氏菌和大腸桿菌的gdh基因序列設計引物。分別在肺炎克雷伯氏菌gdh基因上下游引入EcoRⅠ和SacⅠ酶切位點,大腸桿菌gdh基因上下游分別引入EcoRⅠ和HindⅢ酶切位點。分別以肺炎克雷伯氏菌和大腸桿菌基因組DNA為模板擴增其各自的gdh基因。設計引物如表1所示。
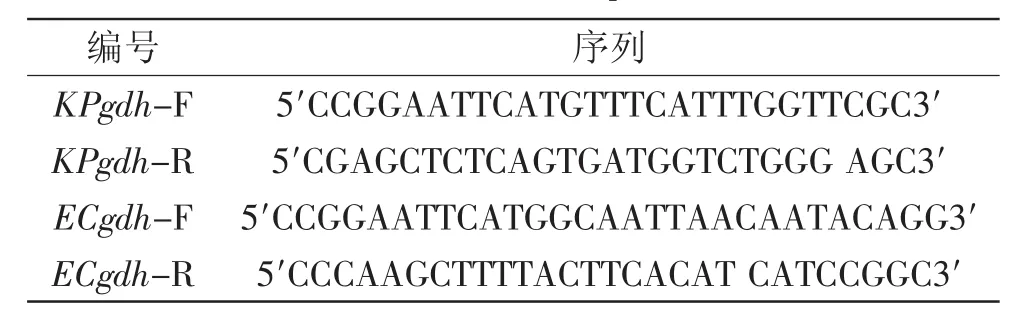
表1 引物序列Table 1 Primer sequences
1.3.2 表達載體的構建 PCR產物回收后酶切,與載體pET-28a連接,連接產物轉化至E.coli DH5α,涂布含有硫酸卡那霉素的抗性平板,37℃靜置培養18 h,挑取陽性克隆,進行菌落PCR及質粒雙酶切鑒定。
1.3.3 重組質粒轉化表達宿主 制備大腸桿菌E.coli BL 21感受態,分別將鑒定正確的兩個重組質粒轉化至E.coli BL21,得到重組菌BL21(pET28a-KPgdh)和BL21(pET28a-ECgdh)。
1.3.4 重組菌的表達 分別接種空白菌E.coli BL21(pET-28a)、重組菌BL21(pET28a-KPgdh)和BL21(pET28a-ECgdh)于M9發酵培養基,pET-28a質粒因采用T7啟動子,需用IPTG誘導來表達葡萄糖脫氫酶。由于IPTG對菌體生長有一定影響,故本實驗在37℃,200 r/min培養至OD600=0.6時,加入IPTG至終濃度1 mmol/L,37℃低溫誘導6 h,收集菌體進行SDSPAGE分析,5%(質量分數)濃縮膠,10%分離膠。
1.4 分析方法
以未轉化重組質粒的原代大腸桿菌E.coli BL21及攜帶空質粒的大腸桿菌E.coli BL21(pET-28a)作為空白對照,每隔3 h測一次菌體濃度和葡萄糖含量,生物量測定采用比濁法[12],葡萄糖含量測定采用3,5-二硝基水楊酸法[13]。取發酵液稀釋至合適倍數,生長曲線和葡萄糖含量分別用分光光度計測定吸光度OD600和OD520。
1.5 酶活性的測定
本研究采用超聲破碎細胞,取發酵液離心8000 r/min,收集菌體,在4℃條件下用pH7.4的PBS緩沖液重懸,超聲破碎,破碎條件為100 W,工作3 s,間隔2 s,循環50次,12000 r/min離心5 min,取上清作為粗酶液。
酶活測定體系為2 mL,即200 μmol/L D-葡萄糖,4 μmol/L NAD,1.2 mmol/L Tris-HCl(pH8.0),20 nmol/L MnSO4,200 μL粗酶液。
酶活測定反應在37℃進行,通過檢測340 nm處吸光度的增量來確定反應體系中NADH的生成量,酶單位定義:37℃下,每分鐘催化生成1 μmol NADH的酶量即為一個酶活力單位。
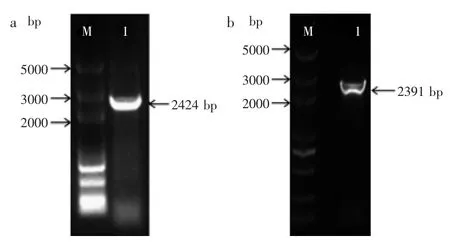
圖1 K.pneumoniae和E.coli中gdh基因的PCR擴增Fig.1 PCR amplification of KPgdh and ECgdh genes from K.pneumoniae and E.coli
2 結果與討論
2.1 gdh基因的克隆
2.1.1 gdh基因的克隆 分別以肺炎克雷伯氏菌和大腸桿菌基因組DNA為模板,PCR擴增葡萄糖脫氫酶基因gdh,結果如圖1所示。
2.1.2 表達載體的構建及轉化E.coli BL21 提取表達載體質粒并進行雙酶切鑒定,結果如圖2所示。酶切結果與理論預期相符,表明重組質粒構建成功。將重組質粒分別轉化至大腸桿菌中,陽性克隆分別命名為BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh),用于蛋白表達及發酵實驗。
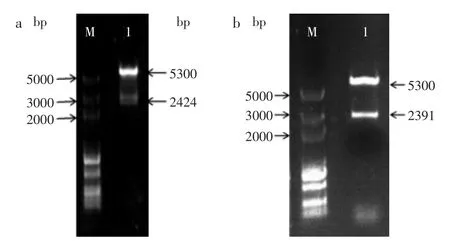
圖2 pET-28a-gdh的雙酶切鑒定Fig.2 Identification of pET-28a-gdh by restriction digestion
2.2 重組菌中葡萄糖脫氫酶基因的表達
將重組質粒分別轉化E.coli BL 21,重組菌和對照菌搖瓶培養,進行總蛋白的SDS-PAGE分析。由圖3可知,重組菌中的葡萄糖脫氫酶均高效表達,分子量大小約為87000 u,符合預期。

圖3 兩種重組大腸桿菌的蛋白電泳Fig.3 SDS-PAGE analysis of two recombinant E.coli strains
2.3 重組菌的發酵
用M9培養基發酵,以空質粒E.coli BL21(pET-28a)和原代菌E.coli BL21為對照組,以重組菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)為實驗組,發酵24 h,每3 h用分光光度計測量其600 nm處的吸光值,繪制生長曲線,結果如圖4。由圖4可知,原代菌E.coli BL21在培養9~12 h后達到平穩期,而攜帶空質粒的對照菌E.coli BL21(pET-28a)因質粒負荷而生長滯后。無論重組菌BL21(pET-28a-KPgdh)還是BL21(pET-28a-ECgdh),其生物量均高于攜帶空質粒的對照菌E.coli BL21(pET-28a),而與原代菌基本持平。重組菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)攜帶的質粒大于空質粒,其中的葡萄糖脫氫酶過表達必然消耗轉錄和翻譯資源,盡管如此,這兩菌的生物量均高于空質粒重組菌,表明過表達葡萄糖脫氫酶促進了菌體生長。Zhu等在研究枯草芽孢桿菌的核黃素生成時,發現過表達葡萄糖脫氫酶能促進菌體生長,重組菌的生長速率和生物量均高于對照菌[14]。Duan等的報道也與本實驗結果相符[15]。這些結果表明葡萄糖脫氫酶的表達與細胞生長有關。

圖4 重組大腸桿菌的生長曲線Fig.4 Growth curve of two recombinant E.coli strains
2.4 葡萄糖剩余量的測定
用M9培養基發酵,以空質粒E.coli BL21(pET-28a)和原代菌E.coli BL21為對照組,以重組菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)為實驗組,發酵24 h。由圖5可知,無論重組菌BL21(pET-28a-KPgdh)還是BL21(pET-28a-ECgdh),其葡萄糖消耗均與原代菌E.coli BL21基本持平,而空質粒重組菌E.coli BL21(pET-28a)的葡萄糖消耗最慢。該結果表明重組菌BL21(pET-28a-ECgdh)和BL21(pET-28a-KPgdh)中葡萄糖脫氫酶超表達促進葡萄糖消耗,也從而促進了菌體生長。
葡萄糖脫氫酶參與磷酸戊糖途徑,該途徑是葡萄糖消耗的重要途徑,結果表明構建的重組菌株在0~9 h的葡萄糖消耗速度大于空質粒菌株,且隨著葡萄糖脫氫酶活性的提高,重組菌的生長速度也快于攜帶空質粒的菌株,基本與原代菌持平。如果扣除質粒復制和轉錄等造成的代謝負荷,葡萄糖脫氫酶超表達明顯促進了菌體生長。
2.5 葡萄糖脫氫酶酶活性的測定
酶活測定反應體系為2 mL,在37℃進行酶催化反應,通過檢測340 nm下的吸光度變化來確定反應體系中NADH的生成量,并用考馬斯亮藍R250標定其總蛋白含量,計算比活力。酶活定義為37℃下每分鐘催化生成1 μmol NADH的酶量,酶活測定結果如圖6所示。由圖6可知,重組大腸桿菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)的葡萄糖脫氫酶活性分別是空質粒對照菌E.coli BL21(pET-28a)的8.5倍和12倍。換言之,KPgdh的活性低于ECgdh,推斷與異源表達有關。由于大腸桿菌對密碼子有偏好性,而肺炎克雷伯氏菌葡萄糖脫氫酶基因KPgdh(GenBank accession No.YP_001335612)與大腸桿菌葡萄糖脫氫酶基因ECgdh(GenBank accession No.YP_003052795)的序列同源性僅為53%,KPgdh作為外源基因在大腸桿菌中表達其酶活性會降低。葡萄糖脫氫酶活性也取決于底物、輔因子、能荷水平乃至信號等多種因素。
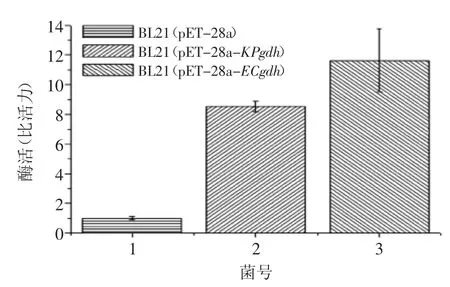
圖6 葡萄糖脫氫酶活性Fig.6 Enzymatic activities of GDH
3 結論
本文為研究超表達不同來源葡萄糖脫氫酶對菌體生長的促進作用,分別以肺炎克雷伯氏菌和大腸桿菌基因組DNA為模板,PCR擴增了各自的葡萄糖脫氫酶基因KPgdh和ECgdh,與pET-28a載體連接后轉化至大腸桿菌BL21,獲得了重組菌BL21(pET-28a-KPgdh)和BL21(pET-28a-ECgdh)。經IPTG誘導,重組菌可實現gdh的高表達,且證實異源表達的葡萄糖脫氫酶活力明顯降低。重組菌以葡萄糖作為碳源在遲緩期和對數生長期生長迅速,在此時期葡萄糖消耗速率與野生菌持平,生物量較空質粒對照菌有所提高。該研究具有雙重意義:a.葡萄糖脫氫酶是糖代謝的關鍵酶,本研究發現重組菌中該酶的活性很高且菌體生長加快,因此為葡萄糖的高效利用及促進細菌生長提供了思路;b.葡萄糖脫氫酶在醫藥和食品工業有著重要作用,但目前價格昂貴,該研究為其發酵生產及在食品領域應用奠定了基礎。
[1]Sakamoto K,Miyoshi H,Matsushita K,et al.Comparison of the structural features of ubiquinone reduction sites between glucose dehydrogenase in Escherichia coli and bovine heart mitochondrial complex I[J].Eur J Biochem,1996,237(1):128-135.
[2]Tsuya T,Ferri S,Fujikawa M,et al.Cloning and functional expression of glucose dehydrogenase complex of Burkholderia cepacia in Escherichia coli[J].J Biotechnol,2006,123(2):127-136.
[3]Navanietha Krishnaraj R,Chandran S,Pal P,et al.Molecular modeling and assessing the catalytic activity ofglucose dehydrogenase of Gluconobacter suboxydans with a new approach forpowergeneration in a microbialfuelcell[J].Current Bioinformatics,2014,9(3):327-330.
[4]Park HJ,Jung J,Choi H,et al.Enantioselective bioconversion using Escherichia coli cells expressing Saccharomyces cerevisiae reductase and Bacillus subtilis glucose dehydrogenase[J].J Microbiol Biotechnol,2010,20(9):1300-1306.
[5]Siddappaji MH,Scholes DR,Bohn M,et al.Overcompensation in response to herbivory in Arabidopsis thaliana:The role of glucose-6-phosphate dehydrogenase and the oxidative pentosephosphate pathway[J].Genetics,2013,195(2):589-598.
[6]Stover NA,Dixon TA,Cavalcanti AR.Multiple independent fusions of glucose-6-phosphate dehydrogenase with enzymes in the pentose phosphate pathway[J].PLoS One,2011,6(8):e22269.
[7]Sangeetha PT,Ramesh MN,Prapulla S G.Recent trends in the microbial production,analysis and application of Fructooligosaccharides[J].Trends in Food Science&Technology,2005,16(10):442-457.
[8]周玨,魏轉,孫文敬,等.D-核糖的生產及應用[J].食品科技,2014,39(14):273-278.
[9]張錦芳,籍小濤,杜連祥,等.D-葡萄糖脫氫酶活力測定方法的研究-短小芽孢桿菌在D-核糖生產中的應用[J].天津輕工業學院學報,2001,38(3):37-40.
[10]唐江濤,覃益民,楊梅,等.利用重組葡萄糖脫氫酶提高低聚果糖純度[J].食品研究與開發,2007,28(12):63-67.
[11]Ashok S,Raj S M,Rathnasingh C,et al.Development of recombinant Klebsiella pneumoniae△dhaT strain for the coproduction of 3-hydroxypropionic acid and 1,3-propanediol from glycerol[J].Applied microbiology and biotechnology,2011,90(4):1253-1265.
[12]Spiller E,Sch?ll A,Alexy R,et al.A microsystem for growth inhibition test of Enterococcus faecalis based on impedance measurement[J].Sensors and Actuators B:Chemical,2006,118(1):182-191.
[13]趙凱,許鵬舉,谷廣燁.3,5-二硝基水楊酸比色法測定還原糖含量的研究[J].食品科學,2008,29(8):534-536.
[14]Zhu YB,Chen X,Chen T,Shi S B,Zhao XM Over-expression of glucose dehydrogenase improves cell growth and riboflavin production in Bacilla subtilis[J].Biotechnol Lett,2006,28(20):1667-1672.
[15]Duan YX,Chen T,Chen X,et al.Overexpression of glucose-6-phosphate dehydrogenase enhances riboflavin production in Bacillus subtilis[J].Appl Microbiol Biotechnol,2010,85(6):1907-1914.
Overexpression glucose dehydrogenase can promote the growth of recombinant Escherichia coli
ZHENG Yue1,PEI Qing-hui2,GE Xi-zhen2,TIAN Ping-fang1,*
(1.College of Life Science and Technology,Beijing University of Chemical Technology,Beijing 100029,China;2.College of Biochemical Engineering,Beijing Union University,Beijing 100023,China)
The genes encoding glucose dehydrogenase(gdh)from Klebsiella pneumoniae and Escherichia coli were amplified by polymerase chain reaction(PCR).Subsequently,the corresponding expression vectors pET-28a-KPgdh and pET-28a-ECgdh were constructed and cloned into E.coli BL21,resulting in two recombinant strains E.coli(pET-28a-KPgdh)and E.coli(pET-28a-ECgdh).SDS-PAGE analysis showed the high level expression of both KPgdh and ECgdh upon IPTG induction.Compared with the control strain E.coli(pET-28a)which harbored an empty vector,the recombinant strains E.coli(pET-28a-KPgdh)and E.coli(pET-28a-ECgdh)consumed more glucose,and the gdh activity toward glucose increased by 8.5-and 12-fold,respectively.If plasmid burden was not counted,overall these results indicated that gdh overexpression facilitated cell growth.
Escherichia coli;Klebsiella pneumoniae;glucose dehydrogenase;overexpression;cell growth
TS201.3
A
1002-0306(2015)22-0189-05
10.13386/j.issn1002-0306.2015.22.031
2015-02-02
鄭躍(1989-),女,碩士,研究方向:微生物與分子生物學,E-mail:yueyue_0307@126.com。
*通訊作者:田平芳(1965-),男,博士,教授,研究方向:合成生物學,E-mail:tianpf@mail.buct.edu.cn。
國家自然科學基金(21076013,21276014)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
