多花水仙HDS基因cDNA全長克隆與序列分析
2015-04-29 21:04:17吳用何炎森李科申艷紅李歡陳曉靜
熱帶作物學報 2015年10期
吳用 何炎森 李科 申艷紅 李歡 陳曉靜


摘 ?要 ?以‘黃花水仙2號和‘金盞銀臺為材料,水仙各自花瓣的cDNA為模板,采用RT-PCR和RACE技術克隆出2個HDS基因,分別命名為NtHDSY和NtHDSJ,測序結果表明,NtHDSY和NtHDSJ基因全長分別為2 592 bp和2 599 bp,2個基因均含有1個2 178 bp的開放閱讀框(ORF),編碼745個氨基酸。同源性分析表明,黃花水仙2號與金盞銀臺、鐵皮石斛、長春花、大豆、葡萄和蘋果的相似系數分別為:97.77%、79.29%、75.29%、75.51%、75.02%和74.40%。Real-time PCR分析表明:NtHDS基因在花瓣和副冠內的表達量在開花過程的花蕾期與盛花期存在明顯差異,推測該基因可能與多花水仙香氣物質的合成有關。
關鍵詞 ?多花水仙;HDS基因;MEP途徑;基因克隆;序列分析
中圖分類號 ?S682.21 ? ? ? ? ? 文獻標識碼 ?A
Cloning and Expression Analysis of HDS
Gene from Narcissus tazetta var
WU Yong1, HE Yansen1,3, LI Ke1,2, SHEN Yanhong1,2,
LI Huan1,2, CHEN Xiaojing1,2*
1 College of Horticulture,Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002,China
2 Institute of Genetics and Breeding in Horticuhural Plants,FAFU, Fuzhou, Fujian 350002,China
3 Mintai Horticulture Research Center, Fujian Academy of Agricultural Sciences, Zhangzhou,Fujian 363005, China
Abstract ?HDS is one of the important enzymes in the pathway of plant MEP. In this study, Huanghua Ⅱ and ?Jinzhanyintai ?were used as the experimental materials.The specific primers were designed according to the cloned HDS gene fragment and EST sequences. Two genes, named NtHDSY and NtHDSJ, were isolated from N. tazetta var by RACE and RT-PCR, and the fragments was about 2 592 and 2 599 bp, respectively. The open reading frame was 2 178 bp, encoding a polypeptide of 745 amino acids. Sequence analysis showed that the amino acid sequence of Huanghua Ⅱ shared 97.77%, 79.29%, 75.29%, 75.51%, 75.02% and 74.40% homologous with Jinzhanyintai, Dendrobium officinale, Catharanthus roseus, Glycine max, Vitis vinifera and Malus domestica,respectively. The real time RT-PCR showed that the transcription expression of HDS gene changed accordingly during flower blooming and flower organs, indicating that the possible role of HDS gene in the synthesis of aroma substances.
Key words ?Narcissus tazetta var; HDS gen; MEP pathway; Gene cloning; Sequence analysis
doi ?10.3969/j.issn.1000-2561.2015.10.017
花香是一系列低分子量、揮發性物質組成的復雜混合物,由花朵釋放來吸引和引導授粉的昆蟲[1],是園藝觀賞植物品質評價的一個重要指標,花香的合成是多種酶基因共同作用的結果。一般來說,花香物質可分為萜類、苯基/苯丙烷基類和脂肪酸衍生物3大類[2]。萜類在所有生物體內都有發現,然而在植物中尤其豐富多樣[3-4]。在多花水仙揮發性成分中,萜類物質所占的比例最多[5]。隨著生物技術研究的不斷深入,利用基因工程手段改良植物香氣物質的組成和含量已成為可能。
大部分的萜類化合物是由前體異戊烯基焦磷酸(isopentenyl diphosphate, IPP)碳骨架的五碳結構單元通過單萜或倍半萜合成酶的作用合成[6]。IPP合成的途徑有2條,分別是位于細胞質中的甲羥戊酸(mevalonic acid, MVA)途徑和位于質體中的甲基-D-赤蘚糖醇-4-磷酸(methyl-D-eryth-ritol-4-phosphate, MEP)途徑[7-8]。其中MEP 途徑是一條高等植物合成萜類次生代謝產物的重要代謝途徑(圖1)[9],該途徑是以3-磷酸甘油醛與丙酮酸為前體物質,通過7步不同的酶促反應生成萜類化合物的5碳單元前體IPP和二甲基烯丙基焦磷酸(dimethylallyl pyrophosphate,DMAPP)。HDS是MEP途徑中第6步催化反應的酶,屬于GCPE 蛋白家族, 定位在質體中,可催化2-C-甲基赤蘚醇-2,4-環焦磷酸(2-C-methy1-D-erythrito1 2,4-cyclodiphosphate,ME-cPP)生成羥基甲基丁烯基-4-磷酸(1-hydroxy-2-methy1-2-(E)-butenyl 4-diphosphate,HMBPP),HMBPP繼續在羥基甲基丁烯基-4-磷酸還原酶(1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase,HDR)的催化下生成IPP和DMAPP,IPP和DMAPP后經異戊烯基轉移酶類的延伸反應以及萜類合酶的修飾作用,合成種類繁多且功能各異的萜類化合物[10-11]。因此,HDS既是萜類物質前體IPP合成的關鍵酶,也是單萜和雙萜類芳香物質、類胡蘿卜素、VE和葉綠素等重要物質合成的關鍵調控位點[12]。目前,高等植物中對HDS基因開展的研究不多,僅從可可、番茄、鐵皮石斛、長春花、甜菊、杜仲、蘋果、黃花蒿等10多種植物中克隆出HDS基因cDNA全長序列,對基因功能的研究僅局限于少數模式植物[13-14]。為此,本研究以多花水仙為材料,根據本課題組構建的多花水仙花瓣抑制消減雜交cDNA文庫獲得的EST序列設計特異引物,利用RT-PCR和RACE技術克隆出水仙HDS基因全長,并對其進行生物信息學分析,為后續利用多花水仙的HDS基因改良其它植物花香的轉基因研究以及深入分析多花水仙MEP代謝途徑的分子機制提供一定的參考。
1 ?材料與方法
1.1 ?材料
實驗材料為多花水仙的2個品種,分別是取自漳州水仙生產基地的‘金盞銀臺和種植于福建農林大學園藝植物遺傳育種研究所的‘黃花水仙2號。‘金盞銀臺副冠橙黃色,花瓣白色;‘黃花水仙2號副冠和花瓣均為黃色,而副冠顏色較淺。分別取花蕾期和盛花期2個時期的水仙花朵的副冠和花瓣,液氮速凍后置于-80 ℃下儲藏備用。
1.2 ?方法
1.2.1 ?總RNA的提取與反轉錄 ? 使用多糖多酚植物總RNA快速提取試劑盒(購自北京白泰克生物有限公司),按照說明書要求分別提取‘金盞銀臺和‘黃花水仙2號副冠和花瓣總RNA。3′-RACE模板按照Revert AidTM First Strand cDNA Synthesis Kit試劑盒說明書(購自Fermenta)進行逆轉錄。5′-RACE模板按照Super SMARTerTM RACE cDNA ?Amplification Kit試劑盒說明書(購自Contech公司)進行逆轉錄。按照SYBRPrime Script RT-PCR Kit(Perfect RealTime)試劑盒說明書(購自TaKaRa)逆轉錄的cDNA用于qRT-PCR。
1.2.2 ?‘黃花水仙2號HDS全長cDNA的克隆 ? ? 根據差減文庫獲得的基因片段設計RACE引物(表1),引物合成與測序分別委托北京六合華大基因科技股份有限公司和上海博尚生物科技股份有限公司。
以‘黃花水仙2號的cDNA為模板,分別進行5′-RACE和3′-RACE的擴增。PCR擴增反應條件:9 ℃預變性5 min;94 ℃變性30 s,50~58 ℃退火30 s,72 ℃延伸2 min,35循環;72 ℃延伸10 min。5′端序列的克隆:以HDS5′-1和UPM為引物進行第一輪擴增,將第一輪反應產物稀釋10倍作為模板,以HDS5′-2和UPM作為引物擴增第二輪。3′端序列的克隆:以HDS3′-1和AUAP為引物進行第一輪擴增,將第一輪產物擴增10倍做為模板,以HDS3′-2和AUAP為模板進行第二輪擴增。
1.2.3 ?‘黃花水仙2號與‘金盞銀臺ORF的克隆 ? 通過DNAMAN軟件對已知的HDS片段序列和克隆得到的3′和5′端序列進行拼接,并在DNAMAN軟件對其ORF進行預測。再根據ORF兩端序列設計特異性上下游引物HDS-U和HDS-A,對‘黃花水仙2號和‘金盞銀臺的編碼區序列進行擴增,退火溫度為55 ℃。通過凝膠電泳回收目的片段,與PGEM載體連接并轉化DH5α(Escherichia coli.),經菌液鑒定后送往博尚生物科技股份有限公司進行測序。
1.2.4 ?序列分析 ? 氨基酸多重序列比對和CDD功能區域分析使用NCBI中的BLASTX(http://blast.ncbi.nlm. nih.gov/Blast.cgi);氨基酸理化分析使用ExPASy-ProtParam tool(http://web.expasy.org/protparam);構建系統進化樹使用MEGA5.05中的鄰近相法neighbor-joining tree;氨基酸序列信號肽預測使用SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP);氨基酸序列跨膜預測使用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/);亞細胞定位使用softberry(http://linux1.softberry.com/berry.phtml)。
1.2.5 ?‘黃花水仙2號和‘金盞銀臺qRT-PCR分析 ? 以Actin作為內參基因,2種水仙的cDNA模板均稀釋5倍,HDSY-L和HDSY-R為上下游引物來檢測HDSY基因在2種水仙中不同時期的轉錄水平。使用25 μL qRT-PCR反應體系:SYBR Premix Ex TaqTM(2×)12.5 μL,PCR Forward Primer(10 μmol/L)0.5 μL,PCR Reverse Primer(10 μmol/L)0.5 μL,cDNA 2.0 μL,ddH2O 9.5 μL,每個樣品設置3次重復。qRT-PCR程序為:Stage1:預變性,94 ℃,3 min;Stage2: PCR反應,94 ℃變性15 s;56 ℃退火35 s,40循環。Stage3:Dissociation。反應結束后,分析Bio-Rad CFX96中的熔解曲線和擴增曲線,導出數據。從不同時期的相關基因擴增曲線中得到Ct值,通過action的校正最終得到目的基因的相對表達量。
1.3 ?數據處理
采用2-△△ct(Livak)法,用Excel軟件處理和分析數據。
2 ?結果與分析
2.1 ?多花水仙HDS基因cDNA全長克隆
以提取的‘黃花水仙2號總RNA反轉錄獲得的cDNA為模板進行PCR擴增,經電泳后發現,5′RACE和3′RACE均獲得約800 bp的片段(圖2)。經過測序并將結果同其原始已知HDS序列進行拼接,得到長度為2 483 bp的基因片段,通過BLAST程序檢索,發現該序列與葡萄(登錄號:XP 002285130.1)、長春花(登錄號: AAO24774.1)和鐵皮石斛(登錄號: AHN91473.1)的HDS基因mRNA序列的同源性分別達到68.64%、66.48%和71.16%,可初步確定該基因片段為‘黃花水仙2號的HDS基因片段。
2.2 ?‘黃花水仙2號和‘金盞銀臺HDS基因ORF的克隆與分析
根據上述拼接的‘黃花水仙2號cDNA全長,分別設計用于基因3′RACE和5′RACE序列擴增的特異引物(HDS-U, HDS-A),對‘黃花水仙2號和‘金盞銀臺水仙HDS基因全長cDNA進行PCR擴增,分別擴增出2 592 bp和2 599 bp大小的條帶。測序結果表明,‘黃花水仙2號ORF為2 238 bp,編碼745個氨基酸;金盞銀臺ORF為2 238 bp,編碼745個氨基酸。‘黃花水仙2號和‘金盞銀臺的HDS基因分別命名為NtHDS40Y(登錄號: KM593243)和NtHDS40J(登錄號:KM593244)。
2.3 ?多花水仙HDS基因的生物信息學分析
利用ExPASy-ProtParam推測2種水仙HDS蛋白,蛋白分子量、等電點、分子式、GRAVY(Grand average of hydropathicity)值、不穩定指數(表2)等基本理化性質,2種水仙的HDS蛋白均屬于穩定蛋白。經TMHMM2.0預測,2種類型水仙的HDS蛋白跨膜螺旋數均為0,不形成跨膜區域,均不是跨膜蛋白。經SignalP4.1預測,2種類型水仙的HDS蛋白均無信號肽,故為非分泌蛋白。經Softberry在線工具分析,亞細胞均定位于膜結合葉綠體中。經NCBI-CDS在線分析,推測此蛋白屬于GcpE Superfamily的成員。
根據開放閱讀框推導的氨基酸進行同源序列比對分析(圖3), ‘黃花水仙2號與鐵皮石斛(Dendrobium officinale)、 長春花(Catharanthusroseus)、 大豆(Glycine max)、 葡萄(Vitis vinifera)、 蘋果(Malus domestica)的相似性分別為: 85.66%、 84.97%、 84.70%、 84.03%和82.01%。說明這個基因的保守性較高,‘黃花水仙2號與‘金盞銀臺也存在一定的氨基酸差異,相似性為97.32%。
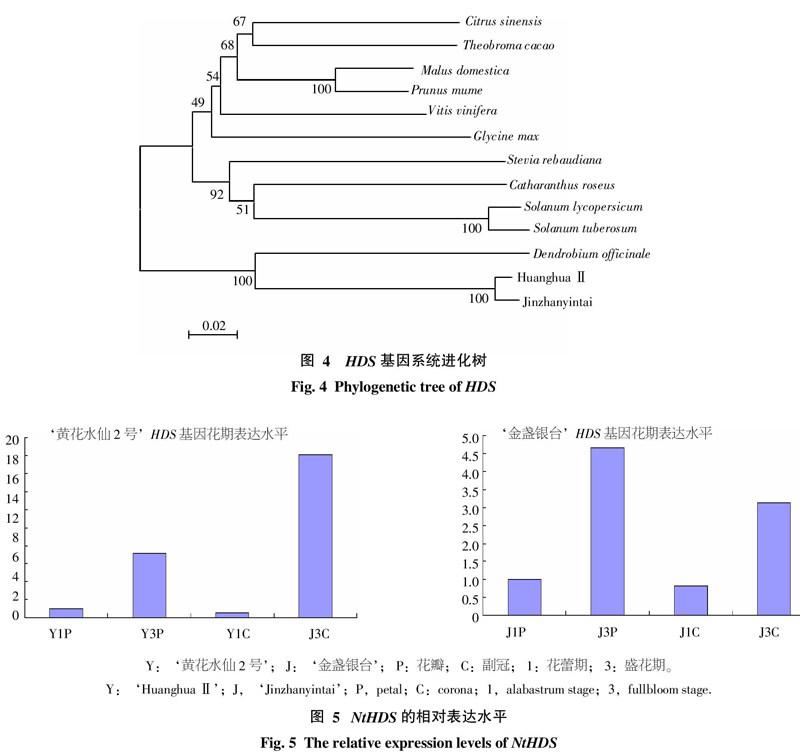
為了研究HDS的進化關系,用MEGA5.05軟件構建了HDS的系統進化樹(圖4),分析表明,‘黃花水仙2號與‘金盞銀臺處于同一分支上,與‘黃花水仙2號進化距離比較遠的是雙子葉植物甜橙、可可、蘋果、梅和葡萄,與單子葉植物鐵皮石斛進化最為相近,馬鈴薯和番茄同屬于茄科,因此它們處在進化樹同一分支上。總體上來看,進化樹基本上與植物分類學相一致。
2.4 ?多花水仙HDS基因在不同發育期的表達分析
根據實時熒光定量PCR擴增獲得的擴增曲線和相關數據,制得‘黃花水仙2號和‘金盞銀臺花瓣與副冠在不同時期HDS基因的表達水平統計圖(圖5)。分析表明,NtHDSY和NtHDSJ的表達水平在花瓣(P)和副冠(C)上的變化趨勢類似,在花蕾期向盛花期轉變過程中,花瓣和副冠中的HDS表達水平均呈上升趨勢。可能是由于NtHDS是花香的主要成分萜類物質合成途徑中的重要酶類基因,表達水平的高低可能與花香物質的合成量多少有關。在花蕾期,花瓣沒有綻放,無明顯香味,NtHDS表達量相對偏低;而在盛花期,花瓣綻放,花香濃郁,NtHDS的表達量也較高。
3 ?討論與結論
HDS是植物萜類物質合成MEP途徑中催化第六步反應所需的酶,該途徑產生的萜類吲哚生物堿(TIAs)是類異戊二烯生物合成的前體物質, 因此HDS被認為是類異戊二烯生物合成途徑中代謝紐帶的一個理想的目標基因[15]。本研究分離并且描述了2種多花水仙中的HDS基因。依據生物信息學的方法,對多花水仙HDS基因進行多角度的分析結果表明:HDS基因編碼的蛋白質無跨膜結構。從不同植物HDS基因編碼氨基酸序列進行同源性比對的結果可知,不同植物HDS基因序列存在一定的差異性,包括同屬水仙屬的‘黃花水仙2號和‘金盞銀臺之間,也存在2.68%的差異性,但從總體上來看,不同植物間HDS基因序列的保守性較高;基于不同植物HDS基因編碼氨基酸序列的進化樹上分析可知,不同植物HDS蛋白的功能在進化上是較為保守的,2個品種水仙在進化上處于同一分支上,而與2種水仙在進化上更為相近的是單子葉植物而不是雙子葉植物,同屬茄科的番茄和馬鈴薯在進化樹上也處于同一分支,這些均與植物分類學一致。
多花水仙開花過程中香味的變化較為明顯。在花蕾期,香氣物質合成水平較低,花瓣包裹較為嚴密,無明顯香味,此時不需要昆蟲授粉;而在盛花期,香氣物質合成水平相對較高,花瓣展開,柱頭分泌粘液,花藥散出花粉,花香濃郁,大量香氣物質的釋放會吸引授粉的昆蟲,這一時期正是授粉的最佳時期。本研究結果顯示,HDS的轉錄水平在開花過程中花蕾期和盛花期呈明顯的時序性。盛花期HDS基因無論是在花瓣還是副冠中,它的表達量都明顯高于花蕾期,這與花香物質的合成是一致的。由于盛花期,植物需要合成更多的香氣物質去吸引昆蟲前來授粉,異花授粉產生的植物多樣性有利于生物對環境的適應,從而來保證后代遺傳的多樣性和穩定性,盛花期花香物質的大量合成是多花水仙與授粉昆蟲協同進化的結果。
目前,高等植物中對HDS基因開展的研究不多,對于基因功能的研究局限于少數模式植物[16-18],尚未涉及有關多花水仙HDS基因的報道。水仙是中國十大名花之一,也是福建省的省花,具有非常高的觀賞價值,因此克隆出多花水仙萜類代謝途徑中的HDS基因具有很好的應用前景。一方面擴充了多花水仙的基因庫,可以給植物花香相關的轉基因研究直接提供目的基因,另一方面能夠對多花水仙MEP途徑分子機制的深入研究奠定基礎。茉莉酸甲酯(MeJA)和紫外(UV)可使得MEP途徑中HDS上游的一個酶基因MECs的表達上調,被稱為誘導子,然而包含茉莉酸甲酯(MeJA),紫外(UV), ?脫落酸(ABA)和丙烯腈(ASA)在內的誘導子對于HDS基因的表達均無直接響。因此,需進一步研究能夠使HDS表達上調的誘導子,再利用基因工程的手段,以期增加TIAs的產量,有效地改良植物花香,更好的誘導昆蟲前來授粉,從而保證了植物群體遺傳的多樣性和穩定性,提高園藝植物觀賞價值,同時對植物花香精油產業的發展有一定推動作用。
參考文獻
[1] 張 ?強, 田彥彥, 孟月娥, 等. 植物花香基因工程研究進展[J]. 基因組學與應用生物學, 2009, 28(1): 159-166.
[2] Campbell T L,Brown E D. Characterization of he depletion of 2-C-Methyl-D-Erythritol-2,4-Cyclodiphosphate ?synthase in Escherichia coli and Bacillus subtilis[J]. Bacteriol, 2002, 184:5 609-5 618.
[3] Chappell J. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants[J]. Ann Rev Plant Physiol Plant Mol Biol, 1995, 46: 521-547.
[4] McGravey D J, Croteau T. Terpene metabolism[J]. Plant cell,1995, 7: 1 015-1 026.
[5] Kuzuyama T, Seto H. Diversity of the biosynthesis of the isoprene units[J]. Nat Prod Rep, 2003, 20: 171-183.
[6] Rodriguez-Concepcio'N M, Querol J, Lois L M, et al. Bioinformatic and molecular analysis of hydroxymethylbuteny1 diphosphate synthase (GCPE) gene expression during carotenoid accumulation in ripening tomato fruit[J]. Planta, 2003, 217:476-482.
[7] Liao Z, Chen M, Gong Y, et al. A new geranylgerany1 diphosphate synthase gene from Ginkgo biloba, which intermediates the biosynthesis of the key precursor for ginkgolides[J]. DNA Seq, 2004, 15: 153-158.
[8] Chappell J. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants[J]. Ann Rev Plant Physiol Plant Mol Biol, 1995, 46: 521-547.
[9] Yue Zheng, Min Chen, Chunxian Yang, et al. Cloning and characterization of MECS and HDS genes from Rauvolfia verticillata[J]. Plant Omics, 2011, 4(2): 82-92.
[10] Dudareva N, Andersson S, orlova I, et al. The nonmenalonate pathway supports monoterpene and Sequiterpene formation in snapdragon flowers[J]. Pore Nat1 Acad Sci USA, 2005, 102(3): 933-938.
[11] 姜 ?丹, 榮齊仙, 袁慶軍, 等. 白花丹參HDS基因的全長克隆與原核表達分析[J]. 藥學學報, 2014, 11: 27.
[12] Pichersky D N. Biochemistry and molecular aspects of floral scent[J]. Plant Physiol, 2000, 122: 627-634.
[13] Gil M J, Coego A, Mauch-Mani B, et al. The Arabidopsis csb3 mutant reveals a regulatory link between salicylic acid-mediated disease resistance and the methyl-erythritol 4-phosphate pathway[J]. Plant, 2005, 44: 155-166.
[14] Kim S M, Kim S U. Characterization of 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate synthase(HDS)gene from Ginkgo biloba[J]. Mol Biol Rep, 2010, 37: 973-979.
[15] Altincicek B, Duin E C, Reichenberg A et al. LytB protein catalyzes the terminal step of the 2-Cmethyl-D-erythritol-4-phosphate pathway of ?isoprenoid ?biosynthesis[J]. FEBS Lett,2002, 532: 437-440.
[16] Manuel R C, Jordi Q, Luisa M L, et al. Bioinformatic and molecular analysis of hydroxymethylbutenyl diphosphate synthase (GCPE) gene expression during carotenoid accumulation in ripening tomato fruit[J]. Planta, 2003, 217:476.
[17] Sang Min K. Soo Un K. Characterization of 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate synthase (HDS) gene from Ginkgo biloba[J]. Mol Biol Rep, 2010, 37: 973-979.
[18] Jordi Q, Narciso C, Santiago I, et al. Functional analysis of the Arabidopsis thaliana GCPE protein involved in plastid isoprenoid biosynthesis[J]. FEBS Lett, 2002, 514: 343-346.
