含笑花藥發育的超微結構研究
2015-07-04 11:14:28何彤慧
西北植物學報 2015年2期
包 晗,何彤慧
(1 寧夏大學 生命科學學院,銀川750021;2 寧夏大學 西部生態與生物資源開發聯合研究中心,銀川750021)
被子植物的花藥結構復雜,組成花藥壁的表皮、藥室內壁、中層和絨氈層4層體細胞相互緊鄰,但形態、結構和功能大不相同。藥室中的花粉發育也是經歷了特殊的細胞分裂和分化方式最后形成了由3個細胞組成的雄配子體。花藥的發育程序受到嚴格的調控,通過許多不同于體細胞發育的方式最終成熟。被子植物的種類很多,花藥的發育方式也有所不同。至今為止還有很多植物花藥的發育特征不清楚。含笑(Micheliafigo)屬于木蘭科(Magnoliaceae)含笑屬(Michelia),為常綠喬木或灌木,葉革質,聚合蓇葖果穗狀。大多數種類花芳香襲人,樹形優美,是可提取精油及綠化、美化環境和庭園觀賞的重要樹種[1]。在被子植物中,含笑屬是比較原始的類型,但在木蘭科中又是較為進化的分類群。敖成齊[2]對含笑的花粉發育做了初步觀察,描述了含笑的花藥壁組成特征和小孢子發生類型。我們對含笑花藥進行了組織化學研究,揭示了該花藥發育中營養物質積累特征,也發現有些與其他被子植物不同的現象[3]。鑒于含笑的系統發育位置和其花藥發育的特征,值得對其花藥發育進行深入研究。本試驗通過對含笑花藥發育中的超微結構變化進行觀察,為研究木蘭科植物胚胎學發育機理提供原始資料。
1 材料和方法
含笑采自廈門大學校園內。分別取不同發育時期的花藥,迅速投入到含2.5%戊二醛、50mmol·L-1二甲胂酸鈉(pH 7.0)緩沖液配制的前固定液中,室溫下前固定3h。之后用相同緩沖液換洗3次,每次30min。清洗后的花藥轉入用50mmol·L-1二甲胂酸鈉(pH 7.0)、1%鋨酸配制的后固定液中,在4℃下固定過夜。第二天早晨用相同緩沖液換洗3 次,每次30 min。然后用梯度系列丙酮脫水,Spurr樹脂包埋。包埋的花藥用Leica Ultracut R 型切片機切成約為70~80nm 厚的切片,將切片撈在噴碳具膜的銅網上,自然晾干后用醋酸雙氧鈾-檸檬酸鉛雙重染色。使用JEM-2100HC 電子顯微鏡觀察,拍照。
2 觀察結果
含笑的花呈現出典型的原始花形態:每朵花中央是數十個離生心皮組成的雌蕊,其周圍有數十個花藥。但其花藥的形態與系統發育進化的被子植物花藥沒有明顯區別。在花藥的橫切面上,可看出花藥由藥壁和藥室組成,花藥壁從外向內,依次分化出表皮、藥室內壁、中層、絨氈層四類細胞結構,藥室中部是花粉發育的部位(圖版Ⅰ,1)。
2.1 造孢細胞時期
雄蕊原基形成后,在其中部形成一些體積較大的細胞,為次生造孢細胞。次生造胞細呈多邊形,細胞壁較薄,有明顯的核,位于細胞中央(圖版Ⅰ,2),核內的染色質呈彌散絲狀。此時的絨氈層細胞形態較規則,橫切面呈方形。有些絨氈層細胞面向藥室的內切向壁與細胞質分開,形成一透明區域。在絨氈層細胞質內分散著較多的小液泡(圖版Ⅰ,3)。
2.2 母細胞時期
從造孢細胞轉變為小孢子母細胞有兩個明顯的特征,細胞之間出現較大間隙并且在小孢子母細胞的表面形成一層較厚的胼胝質壁。在早期小孢子母細胞的細胞質內有豐富細胞器和小液泡,線粒體和質體的體積都較小,內部分化程度也較低(圖版Ⅰ,4)。隨著小孢子母細胞的發育,外表的胼胝質壁不斷增厚,小孢子母細胞原有的纖維素壁降解、消失(圖版Ⅰ,5)。有時在纖維素壁降解部位可看到電子密度較低、類似液泡的區域(圖版Ⅰ,6)。此時,絨氈層細胞的內切向壁開始降解,其質膜與細胞壁之間形成一層電子密度較低的區域;兩個絨氈層細胞之間的徑向壁也開始部分分開,呈現比較疏松的狀態。絨氈層細胞質的電子密度較以前增大,細胞質中形成了很多小液泡,其中有一些絮狀物質(圖版Ⅰ,7)。
2.3 四分體時期
小孢子母細胞經過減數分裂形成四分體,四分體小孢子的表面都被厚厚的胼胝質壁包裹。胼胝質中間有一電子密度較高的薄層(圖版Ⅰ,8)。四分體小孢子內有較多的片層膜結構,其中有些是標志合成細胞內蛋白的多聚核糖體。線粒體體積較小,內部的嵴不明顯,顯示出分化程度較低(圖版Ⅱ,1)。絨氈層細胞的細胞質依然濃厚,有較多質體和線粒體。內切向壁的透明區域仍保留,在內表面上形成了電子密度較高的圓形顆粒,很可能是前烏氏體(圖版Ⅱ,2)。
2.4 小孢子早期
當四分體小孢子的胼胝質壁變薄溶解后,小孢子從中釋放出來。有時游離小孢子細胞核中出現兩個核仁。游離小孢子表面的花粉外壁已形成,由明顯區分的3層結構組成(圖版Ⅱ,3)。小孢子細胞質濃厚,有豐富的細胞器,其中線粒體的體積較大,質體的體積較小(圖版Ⅱ,4)。絨氈層細胞的內切向壁完全消失,只有1層不規則的細胞質膜。在質膜內側,有一些將被分泌到藥室中的原烏氏體,其內部的電子密度較低,外部的電子密度較高,與花粉外壁的電子密度相似(圖版Ⅱ,5)。在小孢子形成花粉外壁時,有一電子密度很低的特定部位,扁平的液泡位于花粉壁和細胞質膜之間。這個部位的花粉外壁是不連續的,是將形成花粉萌發孔的部位(圖版Ⅱ,6)。
2.5 小孢子晚期
被子植物發育小孢子中無論是否形成大液泡,總是呈現出細胞核靠邊分布的特征,呈現出極性分布,這將為小孢子的不等分裂、開始配子體發育作準備。最初在單核中位小孢子中形成了許多小液泡。在細胞質中有較多的線粒體、質體及自體吞噬泡(圖版Ⅱ,7)。隨著小液泡數量的增加,小液泡融合形成大液泡,細胞核和細胞質被擠到細胞的邊緣區域(通常所說的單核靠邊期),小孢子細胞呈現明顯的極性。小孢子中大液泡的形成為小孢子發育晚期的特征。這時花粉外壁結構清晰,表層的覆蓋層和基部的基足層已完全形成,中間是連接覆蓋層和基足層的基粒棒層。雖然此時的花粉內壁還沒有形成,但在花粉外壁與質膜之間有一層電子密度較低的區域(圖版Ⅱ,8)。在小孢子晚期,絨氈層細胞中的基質物質減少,電子密度降低,但細胞內部結構仍完整。在面向藥室的內切向面上仍有一些烏氏體,有些已經脫離絨氈層表面,遷移到藥室中(圖版Ⅲ,1)。
2.6 二胞早期
小孢子經過一次不等有絲分裂后,形成了大小不等的兩個細胞,進入二胞花粉時期。小的為生殖細胞,其細胞核大,細胞質的比例較少,呈透鏡狀緊貼著花粉內壁;大的為營養細胞,繼承了原小孢子的絕大部分內含物。在剛形成的二胞花粉中,兩個細胞除了細胞核的大小有所差異外,細胞質的差異很小(圖版Ⅲ,2)。以后營養核率先遷移到細胞中央。在營養細胞質內又重新出現較多的質體和線粒體(圖版Ⅲ,3)。這時花粉內壁已經形成,與花粉外壁的厚度相近,但電子密度較低且由外向內降低(圖版Ⅲ,4)。二胞花粉早期時的絨氈層細胞已明顯退化,細胞質基質的電子密度進一步減小,膜狀結構也明顯減少,細胞器結構模糊。在絨氈層的內表面上仍有一些烏氏體(圖版Ⅲ,5)。
2.7 成熟時期
到開花時,花粉發育成熟。成熟花粉的外壁形成電子密度較高的3層結構,最外層為覆蓋層,中間為基粒棒層,底部為基足層。細胞質內有一大液泡,內部有脂滴分布(圖版Ⅲ,6)。在細胞質中有一些淀粉粒和脂滴,但淀粉粒較少(圖版Ⅲ,7)。原來在絨氈層的位置形成了絨氈層膜,緊貼中層。其中還有一些烏氏體結構(圖版Ⅲ,8)。
3 討 論
3.1 液泡的變化
液泡是植物細胞內一種特殊的細胞器,也是細胞內膜系統的重要組成部分。在植物細胞的生命活動中,液泡發揮著多種重要的生理功能:參與并調節細胞新陳代謝;控制多種細胞物質的合成、積累和運輸;維持細胞內環境穩定;緩沖外界環境對植物生長發育的影響;提高物種適應環境變化和生存能力;在細胞遺傳信息的調控下發生和發展,代表著植物細胞結構與功能的轉變[4-6]。
液泡的功能多樣,在不同的時間和空間出現的液泡具有不同的功能。在含笑花粉發育中,細胞內的液泡發生了三次比較明顯的變化。第一次是在小孢子母細胞形成胼胝質壁的過程中,可能與胼胝質壁的形成有關。小孢子母細胞合成胼胝質并用分泌小泡轉移至細胞壁外,形成胼胝質壁。第二次是在小孢子母細胞減數分裂之前,原小孢子母細胞的纖維素壁降解,在降解區域形成電子密度較低的液泡,推測是小孢子母細胞分泌的纖維素酶將纖維素細胞壁降解的過程。第三次是在晚期小孢子中形成大液泡將細胞質擠到邊緣,使細胞產生極性,這將為小孢子的不等分裂做準備,使小孢子進入雄配子體發育。其結果是小孢子不等分裂形成一個大的營養細胞和一個小的生殖細胞。細胞核的位移是小孢子不對稱分裂的前提條件之一,保證了不對稱分裂的正常進行[7-8]。小孢子中央大液泡對其未來不對稱性質的分裂以及兩個細胞的不同命運起了重要決定作用。通常單核靠邊期的小孢子核位于萌發孔的對面,而大液泡將細胞核擠到細胞邊緣應是隨機分布。有實驗證明微管對于小孢子核的定位有重要作用[9-10]。在小孢子發育中,大液泡將細胞核擠到細胞邊緣,而細胞骨架則將細胞核定位在萌發孔對面的位置,共同作用創建了單核靠邊的晚期小孢子結構。然而,目前對小孢子中的大液泡形成過程很少有報道。
3.2 花粉壁的形成
成熟花粉壁包括內壁和外壁。外壁由孢粉素構成,分為外壁外層和外壁內層。外壁外層主要由基柱(pilum)組成,基柱包括頭部和棒兩部分。如果基柱頭部合并,即在基柱上形成1層覆蓋層(tectum),棒是分開的。通常外壁可分為2種類型:具覆蓋層外壁(tectate type)和具基柱外壁(pilate type)。具覆蓋層外壁包含覆蓋層、基粒棒層和外壁內層或基足層。基柱型外壁在基柱之間有空隙通向外部[11]。花粉內壁主要為多糖性質的纖維素組成。
在不同植物中,花粉內外壁的結構和組成成分不同。花粉壁的多樣性不僅表現在其表面雕紋差異,還表現在發生過程上[11]。在Platanusacerifolia中,胼胝質壁在花粉外壁的構建和后來外壁的沉積具有積極的作用[12]。在含笑花粉發育中,四分體時期的絨氈層細胞就有烏氏體向藥室中分泌。四分體小孢子釋放后,絨氈層細胞表面的烏氏體大量出現,可觀察到烏氏體向藥室內遷移,烏氏體內含有孢粉素物質,可能用于構建花粉外壁。隨著花粉發育,花粉外壁結構在二胞早期完成,而花粉內壁在二胞花粉時才開始構建。
木蘭科是最原始的被子植物類群之一,花粉形態亦表現出其原始性。含笑外壁超微結構有覆蓋層,基粒棒層和外壁內層或基足層的分化,基粒棒層有小柱形成,但不典型,很少直立小柱,一般小柱傾斜或懸掛在覆蓋層的表面,是為顆粒-初生柱狀層。但外壁內層較薄,內壁加厚。Walker曾指出被子植物花粉外壁結構的演化趨勢是從無結構層→顆粒→初生柱狀層→覆蓋層柱狀層[13]。含笑花粉壁比起木蘭屬有些種類花粉外壁為無結構層,內具小裂隙或大小不等的小腔穴,不形成小柱,屬于進化類群。但是比起鵝掌楸,單性木蘭有小柱典型,短而直立,直徑很小的類群,又相對原始[14-15]。這些特征表明木蘭科含笑屬植物外壁結構的多樣性和原始性。
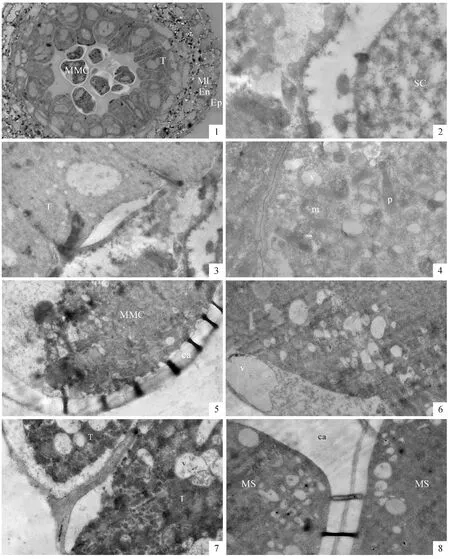
圖版Ⅰ 含笑花藥發育超微結構PlateⅠ Ultrastructural changes in pollen development of M.figo

圖版Ⅱ T.絨氈層;v.液泡;m.線粒體;p.質體;n.細胞核;ub.烏氏體;s.復合淀粉粒PlateⅡ T.Tapetum;v.Vacuole;m.Mitochondria;p.Plasmid;n.Nucleus;ub.Ubisch body;s.Compound starch grains
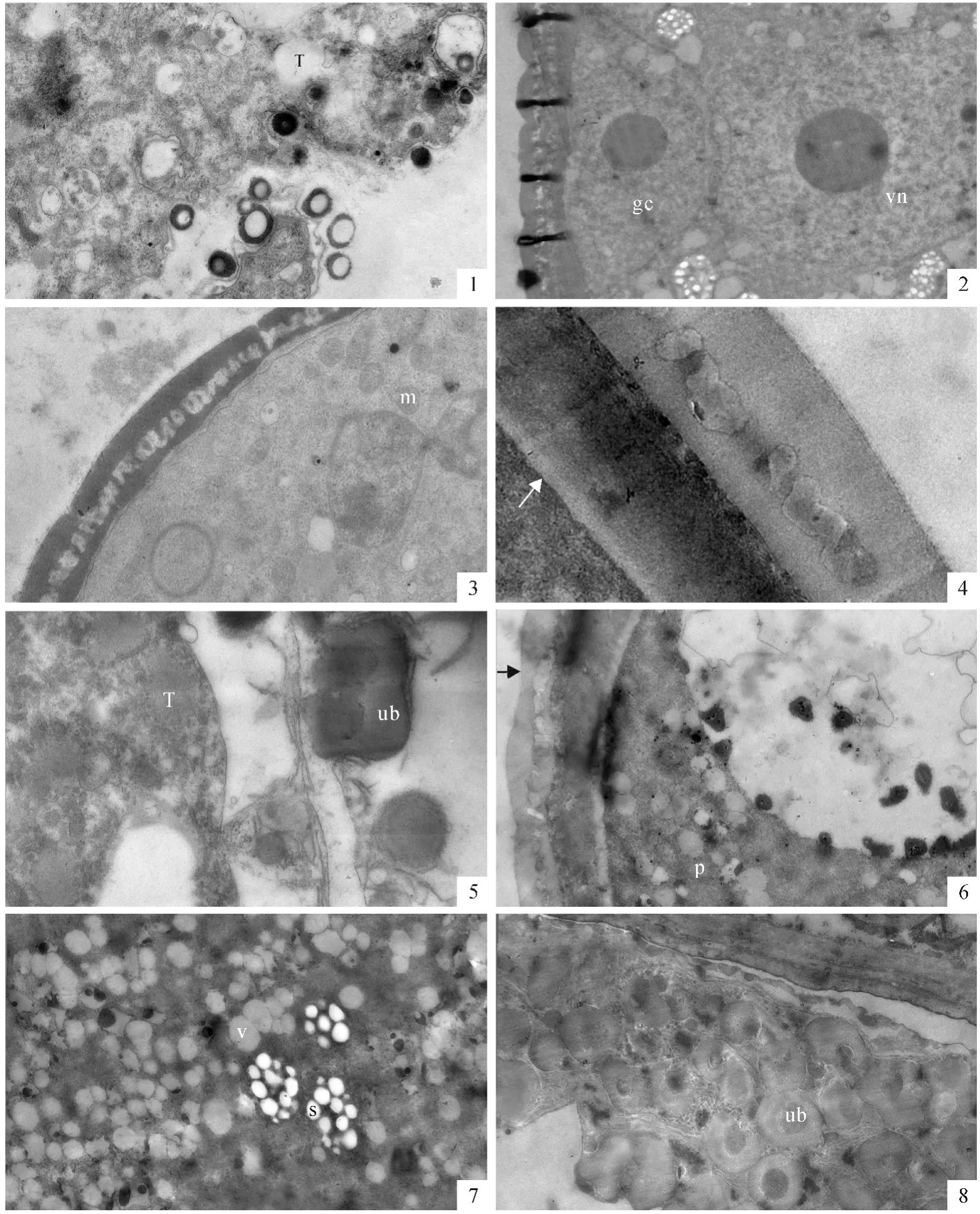
圖版Ⅲ T.絨氈層;gc.生殖細胞;vn.營養細胞核;p.質體;m.線粒體;ub.烏氏體;s.淀粉粒;v.液泡PlateⅢ T.Tapetum;gc.Generative cell;vn.Vegetative nucleus;p.Plasmid;m.Mitochondria;ub.Ubisch body;s.Starch grain;v.Vacuole
[1]ZHANG X H(張新華).Advances in research ofMicheliaLinn[J].Guihaia(廣西植物),2007,27(5):712-719(in Chinese).
[2]AO CH Q(熬成齊).Microsporogenesis,development of male gametophyte inMicheliafigoand their systematic significance[J].Guihaia(廣西植物),2007,27(6):836-839(in Chinese).
[3]BAO H(包 晗),FU SH(傅 劭).Histochemical observation of polysaccharides and lipids on the developing anthers ofMicheliafigo[J].ActaBot.Boreal.-Occident.Sin.(西北植物學報),2013,33(10):1 981-1 982(in Chinese).
[4]LIANG X R(廖祥儒),CHEN T(陳 彤),LIU X L(劉小麗).Formation and function of vacuoles[J].ChineseJournalofCellBiology(細胞生物學雜志),2002,24(2):95-101(in Chinese).
[5]WANG L J(汪良駒),LIU Y L(劉友良).Vacules of pant cells and their physiological functions[J].PlantPhysiologyCommunication(植物生理學通訊),1998,34(5):394-400(in Chinese).
[6]WANG W(王 瑋).The plant vacuole[J].JournalofBiology(生物學雜志),1994,6:9-10(in Chinese).
[7]TWELL D.Mechanisms of microspore polarity and differential cell fate determination in developing pollen[M]//CRESTI M,CAI G,MOSCATELLI A.Fertilization in Higher Plants.Berlin Heidelberg:Springer-Verlag,1999:201-215.
[8]PERIASAMY K,SWANY B G L.Morphology of the tapetum in angiosperms[J].Curr.Sci.,1966,35:427-430.
[9]TERASAKA O,NIITSU T.Unequal cell division and chromatin differentiation in pollen grain cells.Ⅱ.Microtubule dynamics associated with the unequal cell division[J].Bot.Mag.Tokyo,1990,103:133-142.
[10]ZEE S Y,YE X L.Microtubule reorganization during pollen development of rice(OryzasativeL.)[J].Protoplasma,2000,21:188-201.
[11]胡適宜.被子植物生殖生物學[M].北京:高等教育出版社,2005:29-92.
[12]MARIA S C,JESUS M,JUAN S C.Pollen grain and ubisch body development inPlatanusacerifolia[J].ReviewofPalaeobotanyand Palynology,1995,85:63-84.
[13]WALKER J W.Evolution of exine struture in pollen of premitive angiosperms[J].Amer.J.Bot.,1974,61:891-902.
[14]WEI ZH X(韋仲新),WU ZH Y(吳征鎰).Pollen ultrastructure of liriodendron and its systematic significance[J].ActaBotanicaYunnanica(云南植物研究),1993,15(2):163-166(in Chinese)..
[15]XI Y ZH(席以珍),ZHANG Y L(張玉龍),LIN Q(林 祁),etal.A study of pollen wall ultrastructure ofKmeriaseptentrionalis[J].BulletinofBotanicalResearch(木本植物研究),2000,20(4):385-398(in Chinese).
