低溫脅迫對克恩氏冬青超微結(jié)構(gòu)及細(xì)胞內(nèi)Ca2+分布的影響
2015-07-31 00:36:39周余華唐義明趙明明
江蘇農(nóng)業(yè)科學(xué) 2015年3期
周余華 唐義明 趙明明
摘要:在低溫脅迫條件下,分析克恩氏冬青細(xì)胞內(nèi)的超微結(jié)構(gòu)和Ca2+分布與抗寒性的關(guān)系。結(jié)果表明:隨著低溫脅迫的增強(qiáng),細(xì)胞內(nèi)細(xì)胞器受損程度也增強(qiáng),到-16 ℃時(shí),細(xì)胞膜系統(tǒng)出現(xiàn)了嚴(yán)重的損傷,同時(shí)細(xì)胞內(nèi)Ca2+沉淀量隨溫度的降低而增加。
關(guān)鍵詞:克恩氏冬青;低溫脅迫;葉片細(xì)胞;超微結(jié)構(gòu);Ca2+分布
中圖分類號: Q945.78 文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2015)03-0166-03
克恩氏冬青(Ilex×koehneana ‘Emily Bruner)為冬青屬常綠闊葉小喬木,是近期引進(jìn)的歐洲冬青雜交品種。許多冬青屬在低溫脅迫條件下有較好的耐寒性,可以引種到北方地區(qū)[1],一般來說,植物細(xì)胞內(nèi)Ca2+分布及超微結(jié)構(gòu)會(huì)隨著低溫的變化而呈一定的相關(guān)性[2],Ca2+作為胞內(nèi)第二信使調(diào)節(jié)著植物體內(nèi)的許多代謝和發(fā)育過程,因而起重要作用[3-6]。為進(jìn)一步研究克恩氏冬青的生態(tài)適應(yīng)性及其栽培和推廣的制約因素,于2012年對克恩氏冬青的耐寒性進(jìn)行系統(tǒng)研究,解析不同溫度條件下克恩氏冬青的解剖結(jié)構(gòu)和低溫脅迫的關(guān)系,從而深入研究克恩氏冬青的細(xì)胞學(xué)機(jī)制,為克恩氏冬青選育及栽培提供理論依據(jù)。
1 材料和方法
1.1 材料
2012年6月將6種克恩氏冬青扦插于南京林業(yè)大學(xué)內(nèi),待生根后及時(shí)上盆,放置于樹木園溫室內(nèi)待用。
1.2 方法
將樣品在25 ℃下放置1周進(jìn)行預(yù)冷,然后移至人工氣候室(-40~25 ℃),以5 ℃為1個(gè)階梯逐級降溫,分別降到0、-8、-16 ℃時(shí)各維持48 h,及時(shí)采取低溫處理好的植物頂端葉片迅速切成0.5 mm×0.5 mm的組織塊,并沉入到用 0.2 mol/L 磷酸緩沖液(pH值7.2)配制的2%多聚甲醛和2.5%戊二醛混合液中,室溫黑暗初固定6 h;然后用含2%焦銻酸鉀的緩沖液(pH值 7.6)洗滌3次,每次約0.5 h;再轉(zhuǎn)移到含2%焦銻酸鉀的緩沖液(pH值7.6)配制的1%鋨酸中,在4 ℃冰箱內(nèi)固定過夜。將2次固定過的組織塊用重蒸水洗滌4次后,再用pH值為10.0的重蒸水(用0.1 mol/L KOH 調(diào)節(jié)pH值)洗滌2次,每次約0.5 h。隨后經(jīng)系列冷乙醇脫水,環(huán)氧丙烷過渡,Epon812(環(huán)氧樹脂)包埋,LKB-V型超薄切片機(jī)切片,切片經(jīng)0.5%醋酸雙氧鈾染色,在H-600 型透射電子顯微鏡下觀察照相。
2 結(jié)果與分析
2.1 不同低溫脅迫下克恩氏冬青葉片細(xì)胞的超微結(jié)構(gòu)
2.1.1 0 ℃條件下克恩氏冬青葉片細(xì)胞的超微結(jié)構(gòu)
由圖1可知,葉綠體仍然緊貼細(xì)胞壁排列,細(xì)胞核大部分較正常,少部分細(xì)胞核的核膜變得模糊,同樣近一端分布,液泡中有少量高電子密度的嗜鋨物質(zhì)分布;葉綠體中嗜鋨顆粒含量仍然較少且不含淀粉粒,其中葉綠體類囊體片層排列雖然規(guī)則、有序,但少部分清晰可見,大部分已模糊不清,線粒體略正常。
2.1.2 -8 ℃條件下克恩氏冬青葉片細(xì)胞的超微結(jié)構(gòu)
由圖2可知,葉綠體仍然緊貼細(xì)胞壁排列,細(xì)胞核較正常,仍然近一端分布,液泡中含少量高電子密度的嗜鋨物質(zhì);葉綠體中嗜鋨顆粒含量仍較多,同樣不含淀粉粒,其中類囊體片層排列仍然規(guī)則、有序;同樣,葉綠體大部分類囊體片層排列規(guī)則、有序且清晰可見,少部分葉綠體類囊體片層模糊,線粒體略正常。
2.1.3 -16 ℃條件下克恩氏冬青葉片細(xì)胞的超微結(jié)構(gòu)
由圖3可知,葉綠體已不貼細(xì)胞壁排列,葉綠體及其他細(xì)胞器散落在細(xì)胞中;細(xì)胞中含有很少量的高電子密度嗜鋨物質(zhì),多數(shù)葉綠體被膜消失,所含嗜鋨顆粒含量較多,不含淀粉粒,其中葉綠體的類囊體片層擴(kuò)張并且不清晰,細(xì)胞核、線粒體受損(圖3中的箭頭所指的部位為受損部位)。
2.2 低溫條件對克恩氏冬青葉片中Ca2+分布的影響
2.2.1 0 ℃條件下克恩氏冬青葉片中Ca2+的分布情況
由圖4可知,在0 ℃低溫下,克恩氏冬青葉片中的細(xì)胞間隙有Ca2+沉淀,在細(xì)胞壁外側(cè)、葉綠體膜和液泡膜上的Ca2+沉積比細(xì)胞器上多,細(xì)胞核上有少量沉淀。
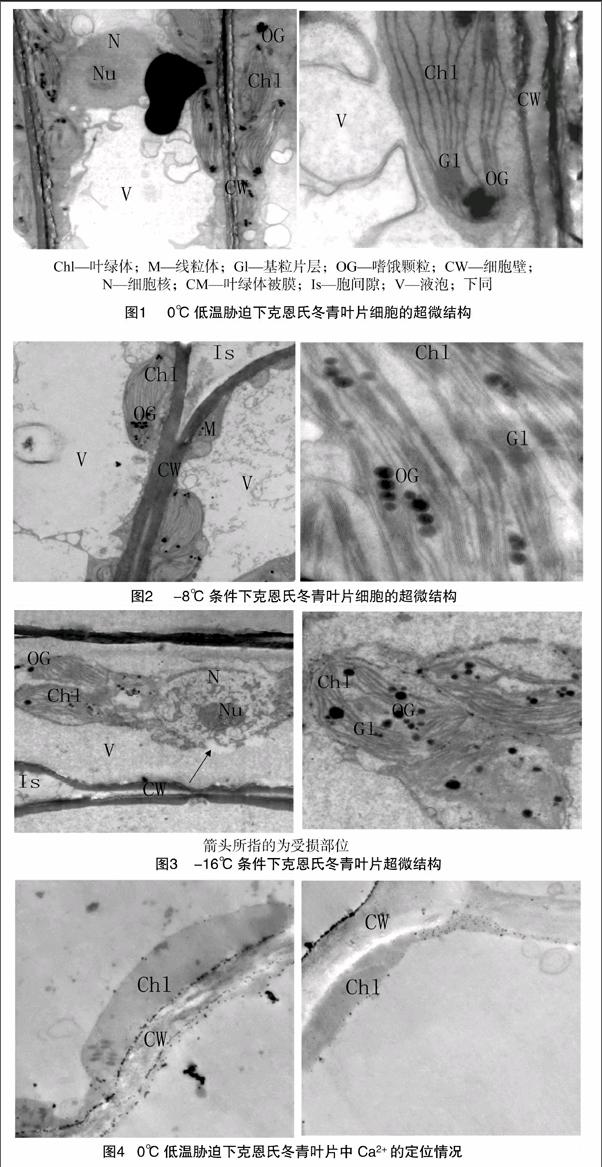
2.2.2 -8 ℃條件下克恩氏冬青葉片中Ca2+的分布情況 由圖5可知,在-8 ℃低溫脅迫下,克恩氏冬青葉片的細(xì)胞壁外側(cè)、細(xì)胞間隙上有Ca2+沉積,大多情況下葉綠體、細(xì)胞壁、液泡上有少量沉積。
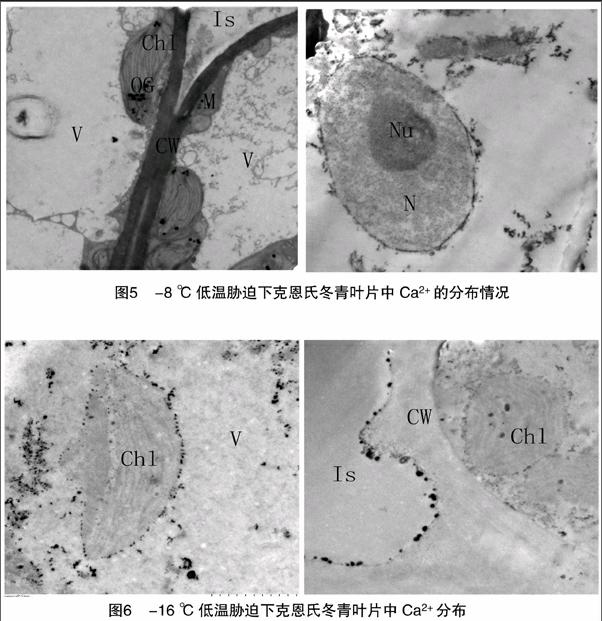
2.2.3 -16 ℃條件下克恩氏冬青葉片中Ca2+的分布情況
由圖5、圖6可以看出,在-16 ℃低溫處理下,Ca2+沉淀明顯多于-8 ℃低溫處理。類囊體片層或消失或模糊不清,葉綠體游離到細(xì)胞內(nèi),不再緊貼細(xì)胞壁,且多數(shù)嚴(yán)重腫脹,細(xì)胞中液泡內(nèi)、細(xì)胞質(zhì)和葉綠體中、細(xì)胞壁上有少量Ca2+沉積(圖6)。
3 結(jié)論與討論
隨著氣溫越來越低,克恩氏冬青葉肉細(xì)胞超微結(jié)構(gòu)出現(xiàn)明顯變化。在 0 ℃條件下,克恩氏冬青葉綠體呈長橢圓形且多數(shù)緊貼細(xì)胞壁分布,細(xì)胞核靠近一端,葉綠體片層清晰可見,液泡和葉綠體中有少量高電子密度的嗜鋨物質(zhì)分布,不含淀粉粒。總體來說,葉片細(xì)胞較正常,葉色較正常。在 -8 ℃ 條件下,細(xì)胞核結(jié)構(gòu)正常,葉綠體稍有膨脹,片層彎曲,嗜鋨顆粒增加,出現(xiàn)了較多的環(huán)狀片層。在-16 ℃條件下,葉片細(xì)胞結(jié)構(gòu)遭到嚴(yán)重破壞,嗜鋨顆粒增多且變大,細(xì)胞內(nèi)大部分細(xì)胞器散亂于細(xì)胞內(nèi)并變得模糊。可見,隨著低溫脅迫的增強(qiáng),細(xì)胞核、葉綠體等細(xì)胞器受損程度也增強(qiáng),細(xì)胞膜系統(tǒng)出現(xiàn)了嚴(yán)重的損傷,主要有葉綠體膜、細(xì)胞膜或解體或模糊不清。在0、-8 ℃脅迫下,克恩氏冬青葉片的基本結(jié)構(gòu)沒有改變,能適應(yīng)此低溫;而到-16 ℃時(shí),細(xì)胞內(nèi)線粒體受到破壞,說明此時(shí)植物體已發(fā)生凍害。克恩氏冬青細(xì)胞內(nèi)的超微結(jié)構(gòu)發(fā)生變化,說明與其抗寒性密切相關(guān)。試驗(yàn)中細(xì)胞內(nèi)的線粒體、葉綠體同樣是對低溫比較敏感的2個(gè)細(xì)胞器,而質(zhì)膜則是低溫傷害的首要部位[7],這說明植物與低溫的適應(yīng)性具有很重要的意義。
克恩氏冬青細(xì)胞內(nèi)Ca2+分布隨溫度的變化而變化。在 0 ℃ 低溫脅迫下,細(xì)胞質(zhì)中的Ca2+濃度很低;到-8 ℃低溫條件時(shí),細(xì)胞內(nèi)Ca2+濃度明顯增加;到-16 ℃時(shí),克恩氏冬青細(xì)胞內(nèi) Ca2+濃度增加,并沉淀成聚集狀態(tài)分布,此時(shí)細(xì)胞內(nèi)Ca2+濃度已經(jīng)超出植物的承受能力,Ca2+濃度平衡被打破,從而破壞和擾亂細(xì)胞正常的結(jié)構(gòu)與功能。細(xì)胞質(zhì)內(nèi)Ca2+變化通過啟動(dòng)細(xì)胞內(nèi)生理生化過程,導(dǎo)致植物的外部抗寒性反應(yīng),起著傳遞和放大信號的作用[8]。此外,Ca2+濃度過度增加會(huì)擾亂以無機(jī)磷為基礎(chǔ)的能量代謝系統(tǒng)[9]。由 Ca2+信使誘發(fā)的這一系列變化致使植物發(fā)生低溫傷害。
參考文獻(xiàn):
[1]張紀(jì)林,謝曉金,教忠意,等. 幾種冬青屬植物抗凍能力比較[J]. 園藝學(xué)報(bào),2005,32(3):477-481.
[2]曾愛松,嚴(yán)繼勇,宋立曉,等. 甘藍(lán)幼苗葉片超微結(jié)構(gòu)及細(xì)胞內(nèi)Ca2+分布對低溫的響應(yīng)[J]. 華北農(nóng)學(xué)報(bào),2011,26(6):129-135.
[3]楊鳳娟,魏 珉,蘇秀榮,等. 不同濃度NO3-脅迫下黃瓜幼苗根系分生區(qū)細(xì)胞內(nèi)Ca2+分布為化的差異[J]. 園藝學(xué)報(bào),2009,36(9):1291-1298.
[4]張銀志,孫秀蘭,劉興華,等. 低溫脅迫和變溫處理對李子生理特性的影響[J]. 食品科學(xué),2003,24(2):134-138.
[5]Blancaflor E B,Masson P H. Plant gravitropism. Unraveling the ups and downs of a complex process[J]. Plant Physiology,2003,133(4):1677-1690.
[6]雷江麗,杜永臣,朱德蔚,等. 低溫脅迫下不同耐冷性番茄品種幼葉細(xì)胞Ca2+分布變化的差異[J]. 園藝學(xué)報(bào),2000,27(4):269-275.
[7]Kratsch H A,Wise R R. The ultrastructure of chilling stress[J]. Plant Cell and Environment,2002,23:337-350.
[8]Bush D S. Calcium regulation in plant cells and its role in signaling[J]. Annu Rev plant Physiol Plant Mol Biol,1995,46:95-112.
[9]Hepler P K,Wayne R O. Calcium and plant development[J]. Ann Rev Plant Physiol,1985,36:397-439.
