抑癌基因PTEN非基因調控的研究進展
2015-08-01 00:01:45尹博煒
基礎醫學與臨床 2015年10期
尹博煒,張 謝,李 宏*
(1.寧波大學 醫學院,浙江 寧波 315211; 2.寧波市醫療中心李惠利醫院,浙江 寧波 315040)
?
短篇綜述
抑癌基因PTEN非基因調控的研究進展
尹博煒1,張 謝2,李 宏2*
(1.寧波大學 醫學院,浙江 寧波 315211; 2.寧波市醫療中心李惠利醫院,浙江 寧波 315040)
抑癌基因PTEN的缺失和突變常常和腫瘤的發生發展密切相關。而除了基因的缺失和突變,PTEN也受到許多非基因機制的調控,如轉錄調控、表觀沉默、非編碼RNA的轉錄后調控、翻譯后修飾。
PTEN;腫瘤抑制因子;PI3K/AKT信號通路;腫瘤
第10號染色體缺失的磷酸酶及張力蛋白同源基因(phosphatase and tensin homologue deleted on chromosome 10,PTEN),又稱晚期腫瘤突變基因,位于人類染色體10q23.3,在多種人類腫瘤中存在突變或缺失,缺失或突變頻率僅次于抑癌基因p53[1]。PTEN蛋白具有酪氨酸磷酸酶活性和脂質磷酸酶活性,其脂質磷酸酶活性能催化磷脂酰肌醇3,4,5-三磷酸(phosphatidylinositol 3,4,5-trisphosphate,PIP3)去磷酸化,使其逆轉為PIP2,從而阻斷PI3K/AKT信號通路。PTEN蛋白功能的缺失可導致PI3K下游信號通路過度激活,引起絲氨酸/蘇氨酸激酶和蛋白激酶B(Akt/PKB)堆積。而Akt/PKB調節許多靶蛋白調控細胞的生物學行為,如凋亡、增殖和細胞運動。因此PTEN表達下降或在細胞中異常分布與腫瘤的發生發展密切相關。近幾年研究發現,非基因型的調控也與PTEN基因的表達關系密切[2](圖1)。
1 轉錄調節
現已發現多種轉錄因子能直接結合PTEN啟動子區調節其表達。轉錄因子c-Jun屬于AP- 1家族。通過結合PTEN啟動子區的AP- 1位點抑制PTEN的表達。Ras/Raf/MEK/ERK通路即是通過上調c-Jun抑制PTEN基因表達,從而促進細胞增殖和抑制細胞凋亡。NF-κB(nuclear factor-kappa B,NF-κB)通過清除轉錄輔激活物CBP/p300負性調控PTEN的表達。抑制NF-κB/miR- 21/PTEN通路可以增加非小細胞肺癌對順鉑的敏感性[3]。有趣的是,Notch1靶向作用于MYC 和Hes1分別誘導和抑制PTEN的轉錄,但因對Hes1的作用較對MYC強,所以Notch1總的效應是下調PTEN的轉錄,從而引起PI3K/AKT通路的過度激活[4]。在多種腫瘤細胞系中誘導抑癌基因P53表達后,PTEN的mRNA水平增加。深入研究發現PTEN基因的上游區域含有P53蛋白的結合位點,P53通過與該位點的結合上調PTEN的轉錄[5]。然而p53的過表達能引起肝細胞性肝癌患者PTEN蛋白水平的下調,但它并非下調PTEN的轉錄而是下調了PTEN蛋白的穩定性。早期生長反應基因1(EGR1)可以與PTEN的啟動子區結合上調PTEN的表達。在許多人類腫瘤中,EGR1的缺失可引起PTEN基因表達下調導致腫瘤的耐輻射性。過氧化物酶體增殖因子活化受體γ(PPARγ) 被活化后與PTEN基因啟動子結合上調人體正常和腫瘤細胞中PTEN的表達,進而調控PI3K/AKT通路誘導細胞死亡。這為PPARγ激動劑治療腫瘤提供了一定的理論依據[6]。
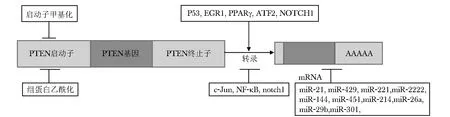
圖1 PTEN非基因調控機制Fig 1 Non-genetic regulation of PTEN
2 表觀沉默
CpG島的高甲基化可引起抑癌基因失活,因此與腫瘤關系密切。48%散發性乳腺癌、16%肝細胞肝癌和>50%甲狀腺癌存在PTEN基因啟動子的高甲基化[7]。PTEN基因啟動子的高甲基化還與藥物抵抗有關。22%費城染色體陽性(Ph+)急性淋巴細胞白血病患者發生PTEN啟動子區高甲基化和PTEN基因下調。Ph+急性淋巴細胞白血病患者更易發生伊馬替尼抵抗,這顯然與PI3K/Akt通路的過度激活有關。而去甲基化能增加PTEN的表達促進細胞凋亡。這些發現提示伊馬替尼抵抗是由或至少部分是由PTEN基因下調引起。相似的發現在慢性骨髓性白血病中也有報道。組蛋白乙酰化作用也有相類似的機制[8]。這些發現提示DNA轉甲基酶和組蛋白脫乙酰基酶對一些惡性腫瘤可能具有抗腫瘤的效應。
3 轉錄后調控
微小RNA(miRNA)是由20~24核苷酸組成的短的RNA分子, 其作用是綁定到目標mRNA上的互補序列,通常引起翻譯抑制或目標退化和基因沉默。miRNA參與PTEN基因的調控,減少多種腫瘤細胞PTEN的表達。例如, MiR- 429、miR- 221和miR- 222在肝細胞癌中直接負性調控PTEN基因,引起肝細胞肝癌增殖、轉移和侵襲能力的增強[9]。在胰島素瘤細胞中,MiR- 144/451能負性調控PTEN表達,引起PI3K/AKT通路過度激活促進細胞的增殖[10]。在非小細胞性肺癌中,miR- 21下調PTEN的表達促進腫瘤生長、侵襲和順鉑抵抗。通過抑制NF-κB降低miR- 21的表達,可以增加非小細胞性肺癌對順鉑的敏感性[3]。miR- 214與3′-UTR結合后,抑制PTEN mRNA的翻譯激活Akt通路,引起人胃癌細胞增殖和順鉑抵抗[11]。在小鼠膠質瘤模型中,MiR- 26a抑制PTEN的表達促進新腫瘤的形成。MiR- 26a也可通過調節腫瘤轉移相關基因和抑制PTEN基因表達促進肺癌的轉移[12]。在乳腺癌中,miR- 29b下調PTEN控制細胞的侵襲,而miR- 301下調PTEN促進細胞增殖、集落生成、轉移、侵襲、他莫昔芬三苯氧胺耐藥和腫瘤生長[13]。
4 翻譯后修飾
4.1 磷酸化
PTEN蛋白的磷酸化能調控蛋白的活性、穩定性及其亞細胞分布。 PTEN蛋白在N端有兩個功能區域:PIP2結合區域(參與PTEN蛋白與膜的相互作用)和發揮磷酸酶作用的區域。此外,PTEN蛋白C端區域包含一個與膜結合所必需的結構C2區域,及一條含PEST區域和PDZ 結合區域的C端尾巴[14]。PTEN蛋白可被多種激酶磷酸化,如CK2、GSK3、RAK、PICT- 1和ROCK[15]。但是,這些激酶如何在腫瘤中,常常發現PTEN啟動子高甲基化。組蛋白乙酰化作用也可能負性調控PTEN的表達。轉錄因子C-Jun和NF-κB抑制PTEN的轉錄,而 P53,EGR1,PPARγ,ATF2作用恰恰相反, Notch1既能抑制也能上調PTEN轉錄。不同微小RNA引起不同的腫瘤PTEN表達的下降。影響PTEN蛋白功能和腫瘤細胞的生物學行為仍不是很清楚。
蛋白激酶CK2是一種真核細胞中普遍存在的信使非依賴性絲/蘇氨酸蛋白激酶。CK2能將PTEN蛋白C端的殘基Ser/Thr磷酸化[16]。由其介導的磷酸化可阻止PTEN因泛素化而降解從而使蛋白更加穩定,然而其磷酸化后的“關閉”的構象阻止了其與配體結合,從而減少了與質膜結合的PTEN蛋白及其脂質磷酸酶活性。總之,CK2通過對PTEN蛋白C端的磷酸化使之更穩定,但是這最終會導致PTEN蛋白的功能失活[15]。
PTEN蛋白C端也能被GSK3β磷酸化。在惡性膠質瘤細胞系中,其殘基Thr366的磷酸化降低了PTEN蛋白的穩定性。該殘基也能被RAK蛋白磷酸化使PTEN蛋白更穩定,在功能方面其作用恰恰和CK2介導的C端磷酸化相反。敲除移植了乳腺癌的小鼠模型的RAK基因,出現腫瘤轉移和轉移瘤的生長,并伴隨PTEN蛋白水平的下降[17]。PICT- 1可通過與C端(與CK2作用區域相同)相互作用直接增加PTEN蛋白的磷酸化作用和穩定性。與CK2相比,PICT- 1同樣能增加PTEN蛋白的穩定性,然而對下游PI3K信號通路的影響恰恰相反,引起這種差異的機制仍不清楚[18]。ROCK可以磷酸化PTEN蛋白C2區域的Ser229和Thr321。在胰腺癌中發現PTEN蛋白磷酸酶活性依賴于ROCK1活化作用[19]。
4.2 泛素化
泛素化修飾也能影響PTEN的表達及亞細胞分布。PTEN主要分布于細胞質,少量分布于細胞核。在細胞質中PTEN蛋白主要通過調節PIP3的水平下調Akt的活性。在細胞核中PTEN蛋白能穩定存在,其對染色體的穩定性、DNA修復和阻斷細胞周期起到更直接的作用。PTEN蛋白的單泛素化是其進入細胞核的機制之一[20]。Lys13 和 Lys289殘基是PTEN蛋白泛素化的位點,可介導其在細胞間穿梭和進入細胞核。而PTEN蛋白的多泛素化導致其在細胞質中滯留及降解,所以其多泛素化具有致瘤性,而其單泛素化因能促進PTEN蛋白向細胞核轉運而具抑癌作用。然而,PTEN蛋白泛素化的機制目前仍存在爭議[21]。NEDD4- 1是第一個被提出能導致PTEN蛋白多泛素化引起蛋白酶體降解的E3泛素連接酶。在PTEN基因未損傷的多種人腫瘤組織和患癌小鼠模型中發現NEDD4- 1高量表達而PTEN蛋白表達水平下降。NEDD4- 1也能單泛素化PTEN蛋白從而調節其進入細胞核。這些結果提示NEDD4- 1可能同時具有致瘤性和潛在的抑瘤作用[22]。除了NEDD4- 1,可能還有其他E3-泛素連接酶參與了PTEN蛋白的泛素化過程,比如Cdh1和WWP2[23]。
4.3 氧化還原作用的調節
活性氧簇(ROS)在癌細胞中常呈現高表達水平,包括H2O2和O2-,因此認為ROS具有促進正常細胞向癌細胞演變的作用。H2O2能氧化PTEN蛋白Cys124殘基的活性位點使之與Cys71殘基形成二硫鍵,引起PTEN蛋白可逆性失活。前列腺癌細胞的ROS高表達水平降低了PTEN蛋白的活性。清除ROS能恢復PTEN蛋白的活性和減弱PI3K介導的信號通路,從而引起前列腺癌細胞發生凋亡[24]。有兩種蛋白能抑制PTEN蛋白的氧化:過氧化物酶Prdx1和硫氧還蛋白相互作用蛋白(Txnip)。Prdx1阻止了由ROS介導的PTEN蛋白的失活,從而抑制了由H-RasV12和ErbB- 2/neu介導的正常細胞向癌細胞的演變過程。Txnip通過調節還原型輔酶II(NAPDH)減少PTEN蛋白的二硫鍵形成,重新激活PTEN蛋白。另外,ROS能將必需脂肪酸非酶促過氧化,如花生四烯酸。過氧化后的必需脂肪酸通過對PTEN蛋白的烷化作用增強Akt信號通路,促進細胞增殖[25]。
5 問題及展望
抑癌基因PTEN編碼的蛋白是具有磷酸酶活性的抑癌蛋白,其在眾多惡性腫瘤中存在突變、缺失或低表達,導致其抑癌功能減弱或喪失。PTEN通過抑制多種信號傳導通路,抑制腫瘤細胞的增殖、侵襲及轉移。其中研究最多的是其抑制PI3K/AKT通路,從而介導細胞凋亡。然而,目前人們對PTEN作用與功能調節機制的認識還處于初級階段,還有許多問題需要深人探索:如PTEN功能失活的基礎還未完全明了,正常的生理調節方式還未見報道,蛋白激酶如何磷酸化PTEN蛋白仍未研究透徹,PTEN蛋白泛素化的機制目前仍存在爭議,其他信號通路之間是否存在相互作用還不清楚,等等。隨著PTEN基因及相關信號傳導通路在腫瘤侵襲及轉移中作用機制的深入研究,相信不遠的將來,PTEN基因的生物治療可能用于臨床治療晚期腫瘤轉移患者,為惡性腫瘤的臨床監測、基因靶向治療、預后評估、藥物療效及新藥研發等提供科學的理論依據,可能成為抑制腫瘤細胞侵襲及轉移的新靶點。
[1] Leslie NR, Maccario H, Spinelli L,etal. The significance of PTEN’s protein phosphatase activity [J]. Adv Enzyme Regul, 2009, 49: 190- 196. doi:10.1016/j.advenzreg.2008.12.002.
[2] Barata JT. The impact of PTEN regulation by CK2 on PI3K-dependent signaling and leukemia cell survival [J]. Adv Enzyme Regul, 2011, 51: 37- 49. doi:10.1016/j.advenzreg.2010.09.012.
[3] Yang Z, Fang S, Di Y,etal. Modulation of NF-kappaB/miR- 21/PTEN pathway sensitizes non-small cell lung cancer to cisplatin [J]. PLoS One, 2015, 10: e0121547. doi:10.1371/journal.pone.0121547.
[4] Yu L, Li F, Zhao G,etal. Protective effect of berberine against myocardial ischemia reperfusion injury: role of Notch1/Hes1-PTEN/Akt signaling [J]. Apoptosis, 2015, 20: 796- 810. doi:10.1007/s10495- 015- 1122- 4.
[5] Stankovic T, Milinkovic V, Bankovic J,etal. Comparative analyses of individual and multiple alterations of p53, PTEN and p16 in non-small cell lung carcinoma, glioma and breast carcinoma samples [J]. Biomed Pharmacother, 2014, 68: 521- 526. doi:10.1016/j.biopha.2014.03.014.
[6] Correia NC, Girio A, Antunes I,etal. The multiple layers of non-genetic regulation of PTEN tumour suppressor activity [J]. Eur J Cancer, 2014, 50: 216- 225. doi:10.1016/j.ejca.2013.08.017.
[7] Nakakido M, Deng Z, Suzuki T,etal. Dysregulation of AKT pathway by SMYD2-mediated lysine methylation on PTEN [J]. Neoplasia, 2015, 17: 367- 373. doi:10.1016/j.neo.2015.03.002.
[8] Nishioka C, Ikezoe T, Yang J,etal. Imatinib causes epigenetic alterations of PTEN gene via upregulation of DNA methyltransferases and polycomb group proteins [J]. Blood Cancer J, 2011, 1: e48. doi:10.1038/bcj.2011.33.
[9] Tang J, Li L, Huang W,etal. MiR- 429 increases the metastatic capability of HCC via regulating classic Wnt pathway rather than epithelial-mesenchymal transition [J]. Cancer Lett, 2015, 364: 33- 43. doi:10.1016/j.canlet.2015.04.023.
[10] Jiang X, Shan A, Su Y,etal. MiR- 144/451 promote cell proliferation via targeting PTEN/AKT pathway in insulinomas [J]. Endocrinology, 2015: en20141966. doi:10.1210/en.2014- 1966.
[11] Yang TS, Yang XH, Wang XD,etal. MiR- 214 regulate gastric cancer cell proliferation, migration and invasion by targeting PTEN [J]. Cancer Cell Int, 2013, 13: 68. doi:10.1186/1475- 2867- 13- 68.
[12] Liu B, Wu X, Liu B,etal. MiR- 26a enhances metastasis potential of lung cancer cells via AKT pathway by targeting PTEN [J]. Biochim Biophys Acta, 2012, 1822: 1692- 1704. doi:10.1016/j.bbadis.2012.07.019.
[13] Shi W, Gerster K, Alajez NM,etal. MicroRNA- 301 mediates proliferation and invasion in human breast cancer [J]. Cancer Res, 2011, 71: 2926- 2937. doi:10.1158/0008- 5472.CAN- 10- 3369.
[14] Song MS, Salmena L, Pandolfi PP. The functions and regulation of the PTEN tumour suppressor [J]. Nat Rev Mol Cell Biol, 2012, 13: 283- 296. doi:10.1038/nrm3330.
[15] Fragoso R, Barata JT. Kinases, tails and more: regulation of PTEN function by phosphorylation [J]. Methods, 2015, 77- 78: 75- 81. doi:10.1016/j.ymeth.2014.10.015.
[16] Shi Y, Paluch BE, Wang X,etal. PTEN at a glance [J]. J Cell Sci, 2012, 125: 4687- 4692. doi:10.1242/jcs.093765.
[17] Yim EK, Peng G, Dai H,etal. Rak functions as a tumor suppressor by regulating PTEN protein stability and function [J]. Cancer Cell, 2009, 15: 304- 314. doi:10.1016/j.ccr.2009.02.012.
[18] Kalt I, Levy A, Borodianskiy-Shteinberg T,etal. Nucleolar localization of GLTSCR2/PICT- 1 is mediated by multiple unique nucleolar localization sequences [J]. PLoS One, 2012, 7: e30825. doi:10.1371/journal.pone.0030825.
[19] Vo K, Amarasinghe B, Washington K,etal. Targeting notch pathway enhances rapamycin antitumor activity in pancreas cancers through PTEN phosphorylation [J]. Mol Cancer, 2011, 10: 138. doi:10.1186/1476- 4598- 10- 138.
[20] Li Y, Low LH, Putz U,etal. Rab5 and Ndfip1 are involved in Pten ubiquitination and nuclear trafficking [J]. Traffic, 2014, 15: 749- 761. doi:10.1111/tra.12175.
[21] Hsia HE, Kumar R, Luca R,etal. Ubiquitin E3 ligase Nedd4- 1 acts as a downstream target of PI3K/PTEN-mTORC1 signaling to promote neurite growth [J]. Proc Natl Acad Sci U S A, 2014, 111: 13205- 13210. doi:10.1073/pnas.1400737111.
[22] Hong SW, Moon JH, Kim JS,etal. p34 is a novel regulator of the oncogenic behavior of NEDD4- 1 and PTEN [J]. Cell Death Differ, 2014, 21: 146- 160. doi:10.1038/cdd.2013.141.
[23] Choi BH, Pagano M, Huang C,etal. Cdh1, a substrate-recruiting component of anaphase-promoting complex/cyclosome (APC/C) ubiquitin E3 ligase, specifically interacts with phosphatase and tensin homolog (PTEN) and promotes its removal from chromatin [J]. J Biol Chem, 2014, 289: 17951- 17959. doi:10.1074/jbc.M114.559005.
[24] Chetram MA, Don-Salu-Hewage AS, Hinton CV. ROS enhances CXCR4-mediated functions through inactivation of PTEN in prostate cancer cells [J]. Biochem Biophys Res Commun, 2011, 410: 195- 200. doi:10.1016/j.bbrc.2011.05.074.
[25] Covey TM, Edes K, Coombs GS,etal. Alkylation of the tumor suppressor PTEN activates Akt and β-catenin signaling: a mechanism linking inflammation and oxidative stress with cancer [J]. PLoS One, 2010, 5. doi:10.1371/journal.pone.0013545.
新聞點擊
精液質量對后代健康影響巨大
據英國《BBC新聞》(BBC NEWS)2014年1月28日報道,澳大利亞的一項研究表明,父親對后代的健康有很重大的影響,在母親受孕的那一個時刻,父親精液的質量或許就決定了胎兒未來成人后的健康狀況。
阿得雷德大學的研究人員發現,精液對后代的一系列發育影響巨大,男孩尤甚。從長遠看,這項研究可確定兒童是否會患上肥胖癥或2型糖尿病。
研究主管羅伯遜(Sarah Robertson)認為,“我們開始了解精液所傳達的復雜信號。”
眾所周知,精液對懷孕影響至關重要,但關于精液對后代健康的影響,還是新發現。這一新的認識還可為不孕夫婦帶來更好的建議和新的選擇。
在低質的精液中受孕勉強生存下來的胎兒出世之后易患上肥胖、高血壓和葡萄糖耐受不良等癥狀。
Advances in the non-genetic regulation ofPTENtumour suppressor activity
YIN Bo-wei1, ZHANG Xie2, LI Hong2*
(1.Medical School of Ningbo University, Ningbo 315211; 2.Ningbo Medical Treatment Center Lihuili Hospital, Ningbo 315040, China)
Mutations and deletions of the tumour suppressor PTEN are frequently involved in the development of cancer. However, PTEN is also tightly controlled by various non-genomic mechanisms,such as the epigenetic silencing of PTEN, post-transcriptional regulation by non-coding RNAs and post-translational modification.
PTEN;tumour suppressor;PI3K-Akt/PKB pathway;cancer
2015- 05- 06
2015- 07- 16
1001-6325(2015)10-1414-05
R34
A
*通信作者(corresponding author):lancet2010@aliyun.com
