灌漿前期低溫脅迫對秈粳稻產量和品質的影響
2015-09-10 11:58:26張榮萍
江蘇農業科學 2015年8期
關鍵詞:產量
張榮萍


摘要: 低溫冷害是我國攀西高原稻區生產中的一大限制因子。以3種秈稻和3種粳稻品種為試驗材料,利用人工氣候室研究灌漿前期低溫脅迫對秈粳稻產量和品質形成的影響。結果表明,灌漿前期低溫脅迫,稻谷產量呈不同程度的下降趨勢,主要是通過影響水稻結實率,尤其是降低二次枝梗結實率,從而導致產量降低,且秈稻結實率下降幅度高于粳稻,秕粒增加幅度大于粳稻。灌漿前期適當的低溫脅迫且天數較短時,有利于降低堊白度和堊白粒率,有助于常規秈稻和粳稻的蛋白質積累,但嚴重低溫脅迫不利于稻米品質的提高。
關鍵詞: 秈稻;粳稻;低溫脅迫;耐冷性;產量;品質;攀西高原
中圖分類號:S511.2+10.1 文獻標志碼:A
文章編號:1002-1302(2015)08-0063-05
水稻結實期是水稻重要的生育階段,結實期最適溫度為21 ℃左右 [1]。但生產上因氣溫劇烈波動,出現高溫熱害和低溫冷害,中國長江流域7月中旬至 8 月中旬常出現持續高溫天氣,是水稻花期高溫危害的重災區。而一些低緯度高海拔地區在水稻開花結實期常出現低溫冷害,影響水稻灌漿結實,導致減產。水稻開花結實期對溫度較敏感,前人關于灌漿期高溫對秈稻產量和品質的影響研究較多 [2-6],而對灌漿期低溫對秈稻和粳稻結實、產量和品質的影響研究相對較少。因此,本試驗主要探討灌漿前期低溫脅迫對攀西稻區秈稻、粳稻籽粒結實及兩者間關系的影響及低溫生理相應機制,旨在為攀西高原稻區水稻生產中減輕和防御灌漿前期低溫傷害提供理論參考和科學依據。
1 材料與方法
1.1 試驗材料
選用立體氣候明顯的攀西高原稻區的西川省西昌市廣泛推廣種植的秈稻、粳稻2種類型的品種(組合) 6個(表1),其開花持續時間基本一致,抽穗7~8 d后開花基本結束。
1.2 試驗設計
1.2.1 試驗方法
試驗于2009和2010年在西昌學院試驗農場和人工氣候室進行。種子消毒浸種2 d,于3月15日播種,采用濕潤育秧,4葉1心時移栽,小區面積6.0 m×2.6 m,小區間隔30 cm,3次重復。以每穴2苗移栽,常規粳稻為167 cm×13.3 cm (45萬穴/hm2),常規秈稻和雜交粳稻為20.0 cm×16.7 cm (30萬穴/hm2),雜交秈稻為20 cm×20 cm (25萬穴/hm2)。按當地常規栽培技術進行田間管理。
當生長進入孕穗期前時,每品種(組合)選取50穴生育進程和長勢基本一致的稻株,帶泥移栽入盆缽,盆缽高 35 cm,直徑 30 cm,每盆裝過篩沙壤土18 kg,施肥量為純氮0.02 g/kg、P2O5 0.008 g/kg、K2O 0.024 g/kg,每盆1穴,每品種(組合)50盆。移栽后及時補水以保持盆缽中有淺水層,擺放于露天農場空地待試驗用。
1.2.2 試驗處理 當生長進入灌漿前期(始穗后8 d)時,移入人工氣候室設定程序自動控制,白天相對濕度70%,晚上相對濕度75%,光照150 μmol/(m2·s),光照處理時間為08:00—20:00。設2個低溫處理,處理1于22 ℃/16 ℃(19 ℃)處理5 d,處理2于22 ℃/16 ℃(19 ℃)處理10 d。室外放置的對照處理7月28日至8月16日平均最低溫度、平均最高溫度和平均溫度分別為20.16、31.63、24.69 ℃,低溫處理結束后移至室外自然條件下至成熟。
1.3 測定項目與方法
1.3.1 酶的活性 處理剛結束時、處理結束后7 d時取籽粒鮮樣液氮處理后于-70 ℃低溫保存用于測定淀粉合成酶(SSS)、淀粉Q酶、ADPG焦磷酸化酶活性,測定參照金正勛等的方法 [6]進行。
1.3.2 考種 成熟時各品種(組合)取5株考種,[JP+1]穗子分為上部(穗最上面的3個一次枝梗)、中部(上部和下部之間的枝梗)和下部(穗子最下面3個一次枝梗),各部按一次枝梗(空粒、秕粒、飽粒)、二次枝梗(空粒、秕粒、飽粒)考種,測定每穗粒數、[JP+1]結實率、秕粒率、特定穎花結實率(所謂“特定穎花”是指穗上最頂部的3個一次枝梗上從上往下數的第3、第4和第5朵穎花,每個穗上共有特定穎花計9朵)、千粒質量。
1.3.3 稻米品質 參照GB/T 17891—1999《優質稻谷》測定糙米率、整精米率、糙米粒形、粒長、粒寬、長寬比、堊白粒率、堊白度、堿消值,采用瑞典FOSS公司生產的1241近紅外快速品質分析儀測定精米蛋白質和直鏈淀粉含量。
1.4 統計分析方法
試驗數據用Excel、DPS、SPSS 11.0進行處理分析。
2 結果與分析
2009年和2010年2年試驗結果基本一致,本文主要對2010年試驗結果進行分析。
2.1 灌漿前期低溫脅迫對秈粳稻結實和產量的影響
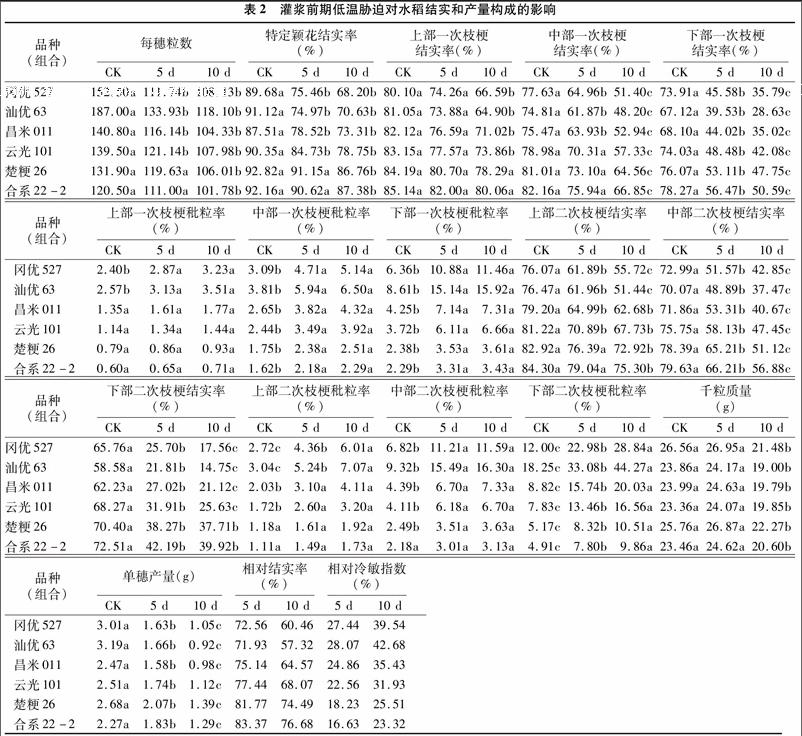
灌漿前期低溫脅迫下,各處理間單穗產量差異極顯著(F=53.955 * *,P=0.000),低溫處理5、10 d的單穗產量較對照顯著降低,降低幅度為汕優63(47.96%、71.16%)>岡優527(45.85%、65.12%)>昌米011(36.03%、60.32%)> 云光101(30.68%、55.38%)>楚粳26(22.76%、48.13%)>合 系22-2(19.38%、43.17%)(表2)。
從產量構成因素看,灌漿前期低溫脅迫對常規粳稻的每穗粒數影響較小,但對秈稻尤其是雜交秈稻每穗粒數影響較大(降幅為17.51%~36.84%)。灌漿前期低溫處理使各品種特定穎花結實率和穗各部一次枝梗和二次枝梗結實率均有不同程度降低,秕粒率增加,且隨脅迫溫度的降低變化幅度增加。處理5、10 d下特定穎花結實率降幅為雜交秈稻(1586%~23.96%)>常規秈稻(10.27%、16.22%)>雜交粳稻(8.86%、14.73%)>常規粳稻(4.77%~9.46%),10 d 處理與對照差異均顯著。低溫脅迫下穗各部一次枝梗結實率降低幅度為上部(3.69%~19.93%)<中部(6.02%~33.17%)<下部(27.44%~45.42%),除5 d處理的穗上部和粳稻穗中部及10 d處理的常規粳稻穗上部與對照差異不顯著外,其余處理與對照差異均顯著;二次枝梗結實率降低幅度分別為上部(6.24%~32.74%)<中部(16.82%~4653%) <下部(4182%~74.83%),除5 d處理的穗上部常規粳稻外,其余處理均與對照差異顯著。低溫脅迫下穗部一次 枝[CM(25]梗秕粒率和二次枝梗秕粒率較對照有明顯增加,增加幅度一次枝梗(882%~84.94%)<二次枝梗(34.29%~14258%),穗上部<穗中部<穗下部,汕優63>岡優527>昌米011>云光101>楚粳26>合系22-2。可見灌漿初期低溫脅迫主要造成穗中部、下部和二次枝梗結實率下降,秕粒率明顯增加,秈稻結實率下降幅度高于粳稻,秕粒增加幅度大于粳稻,說明灌漿前期低溫處理間表現不同,不同類型品種間存在明顯的差異。endprint
與對照相比,灌漿前期低溫處理5 d的千粒質量有所增加,但影響不明顯,而處理10 d的千粒質量顯著降低,降幅由大到小順序為汕優63(20.37%)>岡優527(19.13%)>昌 米011(17.51%)>云光101(15.03%)>楚粳26(13.55%)>合系 22-2(12.19%)。
經相關分析可知,穗粒數、特定穎花結實率、一次枝梗結實率、二次枝梗結實率、千粒質量與產量均呈極顯著正相關,相關系數r分別為0.844 0 * *、0.766 7 * *、0.876 7 * *、0.864 1 * *、0.664 4 * *;而二次枝梗秕粒率與產量呈顯著負相關,相關系數r為-0.483 2*。另外經灰度關聯度分析可知,水稻主要產量要素與產量的關聯度為二次枝梗結實率 (關 聯系數=0.639 4) >一次枝梗結實率 (關聯系數=0588 1) > 特定穎花結實率 (關聯系數=0.546 2) > 穗粒數(關聯系數=0.543 1)>千粒質量 (關聯系數=0.486 4)>二次枝梗秕粒率 (關聯系數=0.323 3)>一次枝梗秕粒率 (關聯系數=0.311 9),說明低溫脅迫主要是通過影響其結實率降低,尤其是二次枝梗結實率降低,從而導致產量降低。
根據相對結實率和相對冷敏感指數的大小(表2)可知,灌漿前期各品種相對耐冷性為合系22-2>楚粳26>云光101>昌米011>岡優527>汕優63。從圖1也可看出,不同類型水稻品種(組合)間的結實率對低溫的耐性是有差異的。為了更好地評價灌漿前期不同類型品種(組合)對低溫脅迫天數的反應特性,用19 ℃處理下空殼率達50%時的天數來表示,岡優527為6.2 d,汕優63為5.1 d,昌米011為7.4 d,云光101為105 d,楚粳26和合系22-2均大于13 d。說明灌漿前期低溫脅迫的天數對結實率的影響與品種耐冷性密切相關,但同一類型品種耐冷性也不同。
[FK(W10][TPZRP1.tif]
2.2 灌漿前期低溫脅迫對秈粳稻品質的影響
2.2.1 外觀品質
從表3可以看出,灌漿前期低溫脅迫對秈粳稻外觀品質性狀的影響不同,影響大小順序為堊白度>堊白粒率>粒寬>長寬比>粒長,且隨低溫脅迫天數的增加,外觀品質性狀變幅增加。與對照相比,低溫處理5 d下堊白度、堊白粒率有所降低,但差異不顯著,而低溫處理10 d下兩者均顯著增加。其中堊白度和堊白粒率增加幅度分別為汕優63(62.46%、25.36%)>岡優527(58.75%、23.80%)>昌米011(47.32%、19.29%)>云光101(38.29%、14.37%)>楚粳26(29.57%、10.16%)>合系22-2(29.21%、10.07%)。低溫脅迫下糙米粒長和粒寬較對照降低,長寬比較對照增加。其中粒寬減少幅度為汕優63(7.29%、11.74%)>岡優527(6.54%、10.77%)>昌米011(4.74%、9.00%)>云光101(4.49%、8.33%)>楚粳26(3.40%、5.76%)>合系22-2(3.12%、5.61%),差異均顯著;粒長較對照降低幅度為1.57%~5.68%,除10 d處理下的秈稻與對照差異顯著,其余處理和品種差異不顯著;長寬比增幅為1.71%~6.81%,與對照相比差異均達到了顯著水平。可見,灌漿初期一定的低溫脅迫天數較短時,有利于降低堊白度和堊白粒率;外觀品質中的堊白度、堊白粒率、粒寬對低溫脅迫較敏感,粒長對低溫脅迫較遲鈍;秈稻尤其是雜交秈稻對低溫脅迫較粳稻敏感。表明低溫脅迫對外觀品質性狀的影響在性狀間和品種(組合)間都存在較大差異。
2.2.2 碾磨品質和理化特性 通過對低溫脅迫和對照相應性狀進行比較發現,低溫脅迫下處理天數不同、品種(組合)不同,其碾磨品質和理化特性表現不同(表4)。低溫脅迫處理5 d下,糙米率較對照略降低,整精米率較對照有所增加,但差異均不顯著,而處理10 d下糙米率、整精米率均顯著降低。與對照相比,處理10 d下, 糙米率、整精米率降低幅度為汕優63(21.62%、28.21%)>岡優527(19.77%、26.75%)> 昌米011(17.19%、22.41%)>云光101(15.31%、2083%)> 楚粳26(12.86%、16.53%)>合系22-2(1134%、1447%)。低溫脅迫使堿消值增加,處理5 d下除雜交秈稻外增幅不顯著,處理10 d下顯著增加(13.17%~26.25%)。與對照相比,低溫脅迫5 d下直鏈淀粉含量略增加,而處理 10 d 下有所降低, 但均變化不明顯。低溫脅迫5 d的蛋白質含量除雜交秈稻較對照降低外,常規秈
稻和粳稻均比對照有所增加,但差異不顯著,在10 d處理下岡優527、汕優63、昌米011、云光101、楚粳26和合系22-2 蛋白質含量分別降低了2408%、25.75%、17.25%、1692%、12.21%、1167%,與對照差異達顯著水平。可見,糙米率、整精米率、堿消值、蛋白質含量對低溫脅迫較敏感,直鏈淀粉含量對低溫脅迫較遲鈍;不同類型品種中秈稻尤其是雜交秈稻對低溫較粳稻敏感。表明在灌漿前期適當低溫處理有利于整精米率、堿消值、直鏈淀粉含量的提高,而對于蛋白質含量來說,灌漿初期適當低溫有助于常規秈稻和粳稻的蛋白質積累,但低溫脅迫會阻礙雜交秈稻蛋白質積累。說明外觀品質性狀的影響在性狀間和品種間都存在較大差異。
2.3 灌漿前期低溫脅迫對秈粳稻籽粒淀粉合成關鍵酶的影響
由圖2可以看出,對照和低溫處理5 d的ADPG焦磷酸化酶活性隨抽穗后生育進程的推進呈單峰曲線變化,到達峰值的時間是在抽穗后13 d,之后均迅速下降,而處理10 d 的ADPG焦磷酸化酶活性隨生育進程推進逐漸降低。低溫處理5 d與對照差異不明顯;低溫處理[JP3]10 d時,粳稻的ADPG焦磷酸化酶活性略高于對照,秈稻的顯著低于對照(降幅16.32%~ 2095%);處理后恢復時ADPG焦磷酸化酶活性迅速降低,降低幅度汕優63(32.35%)>岡優527(30.69%)>昌米011(27.17%)>云光101(11.89%)>楚粳26(8.37%)>合系22-2(7.76%),與對照差異顯著。endprint
抽穗后對照處理的SSS活性逐漸降低,處理5 d的SSS活性呈單峰曲線變化,到達峰值的時間是在抽穗后13 d,之后迅速下降。峰值時岡優527、汕優63、昌米011、云光101、楚粳26、合系22-2分別比對照高32.68、29.21、36.00、44.99、54.98、55.55百分點;處理后恢復5 d時迅速下降。低溫處理10 d和處理后恢復時,粳稻的SSS活性略高于對照,雜交秈稻顯著低于對照,降幅汕優63(16.66%、15.48%)>岡優527(1495%、12.72%),常規秈稻昌米011變化介于兩者之間。
對照處理中Q酶活性隨抽穗后生育進程的推進呈單峰曲線變化,到達峰值的時間是在抽穗后13 d,之后均迅速下降(圖2)。低溫脅迫處理5 d時,雜交秈稻的Q酶活性較對照降低,但差異不顯著,粳稻的Q酶活性均顯著高于對照(增幅16.93%~18.60%)。低溫處理10 d和處理后恢復時Q酶活 性均比對照顯著降低,降低幅度汕優63(15.29%、12.42%)> 岡優527(16.02%、13.54%)>昌米011(20.89%、16.22%)>云光101(25.62%、18.26%)>楚粳26(2895%、22.18%)>合系22-2(30.77%、24.72%)。可見灌漿前期一定的低溫處理有利于Q酶和SSS活性的提高,但隨低溫脅迫天數增加會降低SSS、Q酶和ADPG焦磷酸化酶活性,且對照和低溫處理下耐冷性相對較強的粳稻品種比耐冷性弱的雜交秈稻品種3種酶活性高,低溫脅迫下變化幅度小。
3 討論
3.1 灌漿前期低溫脅迫對秈粳稻結實和產量的影響
水稻灌漿期間溫度過高或過低均會影響水稻籽粒灌
[FK(W31][TPZRP2.tif]
漿 [8],而水稻開花后籽粒灌漿狀況決定結實率和千粒質量,是奪取高產的關鍵 [9-10]。本研究結果表明,灌漿前期低溫脅迫下,單穗產量較對照顯著降低,降低幅度以雜交秈稻最大,常規秈稻次之,后者高于雜交粳稻,常規粳稻最小。從產量構成因素來看,低溫處理間表現不同,不同類型品種間存在明顯的差異。灌漿前期低溫脅迫對常規粳稻的每穗粒數影響較 小,但對秈稻每穗粒數影響較大。處理5 d下千粒質量比對照略增加,而處理10 d的千粒質量顯著降低。低溫處理主要造成穗中、下部和二次枝梗結實率下降,秕粒率明顯增加,秈稻結實率下降幅度高于粳稻,秕粒增加幅度大于粳稻。說明灌漿前期低溫脅迫的天數對結實率的影響與品種耐冷性密切相關。經灰度關聯度分析可知,灌漿前期低溫脅迫主要是通過影響其結實率,尤其是降低二次枝梗結實率,從而導致產量降低。
3.2 灌漿前期低溫脅迫對秈粳稻品質的影響
稻米品質主要受遺傳因子控制和栽培因素的影響 [11-26],但在諸多因子中,溫度對稻米品質影響最大 [14-17]。呂文彥等研究發現,堊白粒率、堊白面積變化與日均溫<21 ℃天數呈正相關,且堊白的形成隨著時間的推移逐漸增加達到高峰以后又有一定的下降 [18,26]。這與Resurrecion等的研究結果 [19-20]趨勢基本一致。本研究表明,灌漿前期低溫脅迫對秈粳稻外觀品質性狀的影響不同,影響大小順序為堊白度>堊白粒率>粒寬>長寬比>粒長,且隨著低溫脅迫天數增加,外觀品質性狀變幅增加。與對照相比,低溫處理5 d下堊白度、堊白粒率有所降低,但差異不顯著,而低溫處理10 d下兩者均顯著增加。其中堊白度和堊白粒率增幅以雜交秈稻最大,常規秈稻次之,后者高于雜交粳稻,常規粳稻最小。低溫脅迫下糙米粒寬和秈稻粒長較對照顯著降低,長寬比除5 d處理下的粳稻外均較對照顯著增加。說明灌漿前期一定的低溫脅迫天數較短時,有利于降低堊白度和堊白粒率;外觀品質中的堊白度、堊白粒率、粒寬對低溫脅迫較敏感 [26],粒長對低溫脅迫較遲鈍;秈稻尤其是雜交秈稻對低溫脅迫較敏感,粳稻外觀品質性狀在低溫脅迫5 d處理下影響不明顯。表明低溫脅迫對外觀品質性狀的影響在性狀間和品種間都存在較大差異。
李林等研究認為灌漿前期日均溫在26 ℃以上或21 ℃以下都會使稻米加工品質下降 [12,26]。本研究表明,低溫脅迫下處理天數不同、品種不同,其碾磨品質和理化特性表現不同。低溫脅迫處理5 d下,糙米率較對照略降低,整精米率和堿消值較對照有所增加,但差異均不顯著,而處理10 d下,糙米率、整精米率、蛋白質含量均顯著降低,堿消值顯著增加。這與殷延勃等提出糙米率和精米率與灌漿結實期溫度呈負相關的研究結果 [21,26]有一定的差異性。稻米的直鏈淀粉含量受遺傳和環境條件的共同影響 [2,23-25],而環境對直鏈淀粉含量影響最重要的因子是結實期的溫度 [25-26]。關于直鏈淀粉含量同結實期溫度的關系,目前尚無定論 [26]。本試驗結果可知,與對照相比,低溫脅迫5 d下直鏈淀粉含量略增加,而處理 10 d 下有所降低,但變化不明顯。蔣李健等也證實了高溫可促進籽粒蛋白質含量的增加,低溫則在一定程度上抑制了蛋白質含量增加 [26-27]。本研究結果還表明,低溫脅迫5 d的蛋白質含量除雜交秈稻較對照降低外比對照有所增加,而 10 d 處理下顯著降低。說明糙米率、整精米率、堿消值、蛋白質含量對灌漿前期低溫脅迫較敏感,直鏈淀粉含量對低溫脅迫較遲鈍;不同類型品種中秈稻尤其是雜交秈稻對低溫較秈稻敏感。在灌漿前期適當低溫處理有利于整精米率、堿消值、直鏈淀粉含量的提高,而對于蛋白質含量來說,灌漿前期適當低溫有助于常規秈稻和粳稻的蛋白質的積累,但低溫脅迫會阻礙雜交秈稻蛋白質的積累。說明外觀品質性狀的影響在性狀間和品種間都存在較大差異。
3.3 灌漿前期低溫脅迫對秈粳稻籽粒淀粉合成關鍵酶的影響
本研究結果得出,低溫處理5 d的ADPG焦磷酸化酶和SSS酶隨抽穗后生育進程的推進呈單峰曲線變化,到達峰值的時間是在抽穗后13 d,之后均迅速下降;處理10 d 的ADPG焦磷酸化酶隨生育進程推進逐漸降低,而粳稻的SSS酶活性略高于對照,雜交秈稻顯著低于對照。 低溫脅迫處理5 d時,粳稻的Q酶均顯著高于對照;低溫處理10 d和脅迫解除后Q酶活性均比對照顯著降低。說明灌漿初期一定的低溫處理有利于Q酶和SSS酶活性的提高,但隨低溫脅迫天數增加會降低SSS、Q酶和ADPG焦磷酸化酶活性,且對照和低溫處理下耐冷性相對較強的粳稻比耐冷性弱的雜交秈稻3種酶活性高,低溫脅迫下變化幅度小。程方民等研究也表明,Q酶活性可能是溫度對稻米淀粉品質變化產生影響作用的酶調節位點,其不同溫度下的活性表現可能存在一定的品種類型特征 [5,26]。endprint
[HS2*2][HT8.5H]參考文獻:
[1] 凌啟鴻. 水稻精確定量栽培理論與技術[M]. 北京:中國農業出版社,2007:125-140.
[2]陳留根,鄭建初,張 彬,等. 抽穗期高溫對水稻產量構成要素和稻米品質的影響及基因型差異[J]. 江蘇農業學報,2005,21(4):249-254.
[3]程方民,胡東維,丁元樹. 人工控溫條件下稻米堊白形成變化及胚乳掃描結構觀察[J]. 中國水稻科學,2000,14(2):83-87.
[4]湯日圣,鄭建初,陳留根,等. 高溫對雜交水稻籽粒灌漿和劍葉某些生理特性的影響[J]. 植物生理與分子生物學學報,2005(6):22-24.
[5]程方民,蔣德安,吳 平,等. 早秈稻籽粒灌漿過程中淀粉合成酶的變化及溫度效應特征[J]. 作物學報,2001,27(2):202-204.
[6]金正勛,楊 靜,錢春榮,等. 灌漿成熟期溫度對水稻籽粒淀粉合成關鍵酶活性和品質的影響[J]. 中國水稻科學,2005,19(4):377-379.
[8]李健陵,張曉艷,杜堯東,等. 高溫對抽穗開花期至灌漿結實期水稻源庫特性的影響[J]. 中國農業氣象,2013,34(1):23-29.
[9]王 華,杜堯東,陳新光,等. 廣東早稻高產的適宜氣溫指標[J]. 生態學雜志2012,31(3):594-599.
[10] 矯 江,祖世亨. 黑龍省主要氣象災害對糧食產量的影響及減災對策[J]. 自然災害學報,2002,(3):18-21.
[11]孫義偉. 水稻灌漿成熟期氣溫對稻米品質的影響[J]. 水稻文摘,1993,12(2):6-8.
[12]李 林,沙國棟,陸景淮.水稻灌漿期溫光因子對稻米品質的影響[J]. 中國農業氣象,1989,10(3):33-38.
[13]唐建軍,陳 欣. 環境條件和稻米品質綜述[J]. 耕作與栽培,1985(5):39-44.
[14]Toshio T. Relation between mean air temperature during ripening period of rice and amylographiccharacteristics or cooking quality[J]. Japanese Journal of Crop Science,1999,68(1):45-49.
[15]孟亞利,高如嵩,張嵩午. 影響稻米品質的主要氣候生態因子研究[J]. 西北農業大學學報,1994,22 (1):40-43.
[16]趙同華,張志猛,馮會中. 生態條件與農藝措施對稻米品質影響的研究進展[J]. 河北農業大學學報,1992,15(1):93-96.
[17]趙式英. 灌漿期氣溫對稻米食用品質的影響[J]. 浙江農業科學,1983(4):178-181.
[LM]
[18] 呂文彥,邵國軍,曹萍,等. 灌漿結實期日均溫度對稻米品質影響的研究進展[J]. 遼寧農業科學,1998(4):1-6.
[19]Resurrecion A P.Effect of temperature during ripening on grain quality on rice [J]. Soil Science and Plant Nutrition,1977,23(1):109-112.
[20]張 磊,吳冬云,朱碧巖. 灌漿期不同溫光對水稻葉、籽粒可溶性蛋白質及可溶性糖動態變化的影響[J]. 華南師范大學學報,2002(2):98-101.
[21]殷延勃,朱美靜,馬洪文,等. 環境因子對寧夏水稻品質性狀的影響——水稻主要品質性狀對環境因子的逐步回歸分析[J]. 寧夏農林科技,2002(2):17-19.
[22]程方民,朱碧巖,吳永常,等. 結實期環境溫度對稻米糊化溫度影響的時段效應分析[J]. 陜西農業科學,1996(1):57-59.
[23]賈志寬,朱碧巖. 灌漿期氣溫的分布對稻米直鏈淀粉累積效應的研究[J]. 陜西農業科學,1990(4):5-7.
[24]李 欣,顧銘洪. 稻米品質研究.灌漿期間環境條件對稻米品質的影響[J]. 江蘇農學院學報,1989,10(1):7-12.
[25]沈 鵬,金正勛,羅秋香,等. 水稻灌漿過程中籽粒淀粉合成關鍵酶活性與蒸煮食味品質的關系[J]. 中國水稻科學,2006(1):23-24.
[26]蔣李健. 結實期溫度脅迫對水稻產量和品質的影響[D]. 揚州:揚州大學,2009:25-33.endprint
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
