康氏木霉固體發酵木聚糖酶條件的研究
2015-10-10 06:51:07易旭東林元山
中國釀造 2015年10期
馮 波,尹 翌,2,易旭東,林元山,2*
(1.湖南農業大學生物科學技術學院,湖南長沙410128;2.長沙水地沙生物科技有限公司,湖南長沙410125)
康氏木霉固體發酵木聚糖酶條件的研究
馮波1,尹翌1,2,易旭東1,林元山1,2*
(1.湖南農業大學生物科學技術學院,湖南長沙410128;2.長沙水地沙生物科技有限公司,湖南長沙410125)
通過固體發酵培養,經單因素及正交試驗分析,得出康氏木霉發酵產木聚糖酶最優條件組合為:麩皮與玉米芯質量比為2∶8,硫酸銨2.5%,MnSO40.50%,料水比1∶1.5(g∶mL),培養基初始pH自然,培養溫度30℃,發酵時間6 d。在此條件下,康氏木霉固體發酵木聚糖酶活力達11.98 IU/g。該木聚糖酶水解產物富含2~5個木糖分子的低聚木糖。
康氏木霉;木聚糖酶;固體發酵;優化
木聚糖(xylan)是自然界中含量非常豐富的一類多糖,僅次于纖維素。木聚糖結構十分復雜,包含有多種不同的側鏈基團,需要在一系列具有不同功能和作用方式的水解酶類相互協同作用下才能將其徹底降解[1-2]。木聚糖酶(xylanase)是將木聚糖水解成低聚木糖和木寡糖的一類酶的總稱,包含有多種內切酶和外切酶的一個復合酶系,主要分為內切β-1,4-D-木聚糖酶,β-D-木糖苷酶,α-L-呋喃阿拉伯糖苷酶,α-D-葡萄糖醛酸苷酶,乙酰木聚糖酯酶和酚酸酯酶等[3-4]。近年來,木聚糖酶已經在很多行業展露出潛在的優勢,可應用于功能性低聚糖的生產、面團改良劑、飼料添加劑,在造紙、醫藥、紡織等行業也有廣泛應用[5-6]。功能性低聚糖是雙歧桿菌增殖因子[7],能夠調節畜禽腸道益生菌群,減少抗生素的使用,實現環保養殖[8-9]。目前,功能性低聚木糖在飼料行業已成為熱點,市場前景良好,李敏康等[10]進行了木霉固態發酵麥麩制備低聚木糖的初步研究。本試驗旨在獲取康氏木霉內切木聚糖酶,以制備功能性低聚木糖,并通過優化產酶條件提高木聚糖酶活力。
1 材料與方法
1.1材料與試劑
1.1.1試劑
V900513櫸木木聚糖:美國Sigma公司;3,5-二硝基水楊酸(3,5-dinitrosalicylic acid,DNS)、苯酚、無水亞硫酸鈉、四水酒石酸鉀鈉、氫氧化鈉、正丁醇、冰醋酸、二苯胺、苯胺、木糖等均為國產分析純。
1.1.2菌種
康氏木霉(Trichoderma koningiiLys-368):湖南農業大學生科院實驗室保存。
1.1.3培養基
麩皮斜面培養基:麩皮汁5.0%,葡萄糖1.0%,瓊脂2.0%,水100 mL,自然pH,121℃滅菌25 min。
固體發酵初始培養基:玉米芯粉(過10目篩)10.0 g,(NH4)2SO41.0%,蒸餾水20 mL,pH自然,攪勻于250 mL三角瓶中,121℃滅菌25 min。
1.2儀器與設備
MJX-250BⅢ霉菌培養箱:天津泰斯特儀器有限公司;V-5000紫外可見分光光度計:上海元析儀器有限公司;SX-500立式壓力蒸汽滅菌鍋:日本TOMY KOGYO公司;CAV812電子天平:日本OHAUS公司。
1.3方法
1.3.1木聚糖酶活力測定方法
酶活力單位定義為:在pH值5.0,溫度50℃條件下,每分鐘水解木聚糖底物溶液生成1 μmol木糖所需的酶量為一個酶活力單位(IU)。
木糖還原糖的測定:參照GHOSE T K[11]的方法,采用3,5-二硝基水楊酸(DNS)法測定。
酶活力的計算:酶活力按每克固體曲所產生的酶量進行計算,其中木糖含量由木糖標準曲線(Y=1.327 7X-0.033 8,X為OD540nm,R2=0.998)計算所得,酶活力計算公式如下:
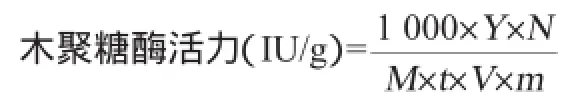
式中:Y為通過木糖標準曲線計算出的木糖含量,mg;N為酶液的總稀釋倍數;M為木糖相對分子質量150.13;t為反應時間30 min;V為反應體系中加入的酶液體積0.1 mL;m為固體發酵原料的質量10 g。
1.3.2康氏木霉固體發酵產木聚糖酶的條件研究
單因素試驗:以固體發酵基礎培養基,采用單一變量原則,以酶活力為評價指標,先后確定發酵時間,溫度,碳源配比,氮源種類,氮源濃度,培養基料水比,培養基起始pH值,金屬離子對康氏木霉固體發酵產木聚糖酶的影響。每組試驗分別做3組平行,取平均值。
正交試驗:選擇單因素試驗影響較大的金屬離子添加量,氮源,發酵時間,料水比4個因素,進行3水平正交試驗,確定最優發酵條件組合。試驗數據采用SPSS18.0軟件進行分析。
1.3.3木聚糖酶解產物分析
將1.0%木聚糖于50℃水浴中酶解反應12 h,酶解產物薄層色譜定性檢測,參照LIN Y S等[12]的方案適當修改進行,薄層色譜檢測條件如下:
展開劑為正丁醇∶冰醋酸∶水(4∶1∶1,V/V);標樣為1%木糖,上樣0.5 μL;酶解產物上樣1.5 μL;顯色劑:2.0 g苯胺溶于2 mL二苯胺以及10 mL 85%H3PO4中;顯色條件:當展開劑上行至距硅膠板頂端0.5 cm處時,取出硅膠板,熱風風干,噴霧顯色劑,再將硅膠板置于80℃條件下顯色10 min,拍照記錄。
2 結果與分析
2.1發酵周期的測定
以固體發酵初始培養基為基礎,接種后置于30℃霉菌培養箱中進行培養,從第2天開始,每天取樣測定木聚糖酶活力,其木聚糖酶活力變化曲線見圖1。
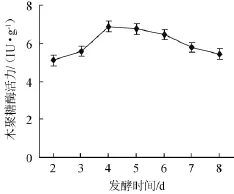
圖1 發酵時間對康氏木霉產木聚糖酶的影響Fig.1 Effect of fermentation time on xylanase production by Trichoderma koningii
從圖1可以看出,康氏木霉在前3 d生長較為緩慢,木聚糖酶活力較低,至第4天時酶活力達到高峰(6.89 IU/g),此后緩慢下降。第4天至第6天木聚糖酶活力無顯著差異(P<0.05,下同),因此,選擇康氏木霉固體發酵產木聚糖酶的周期為4~6 d。
2.2最適培養溫度的測定
以固體發酵初始培養基為基礎,接種后分別置于28℃,30℃,32℃,35℃,37℃的恒溫培養箱中培養4 d,木聚糖酶活力變化結果見圖2。
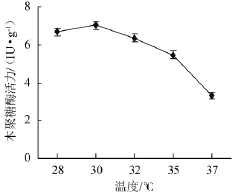
圖2 溫度對康氏木霉產木聚糖酶的影響Fig.2 Effect of temperature on xylanase production by Trichoderma koningii
由圖2可知,在28℃和30℃條件下所產木聚糖酶活力無顯著差異,與其他溫度條件差距明顯,>30℃后酶活力下降。康氏木霉固體發酵產木聚糖酶的最佳培養溫度為30℃,此條件下木聚糖酶活力達7.03 IU/g。
2.3碳源配比的確定
以初始培養基為基礎,碳源條件選擇不同麩皮和玉米芯粉質量比0∶10,1∶9,2∶8,3∶7,4∶6,5∶5,6∶4,7∶3,8∶2,9∶1,10∶0進行發酵試驗,木聚糖酶活力結果見圖3。
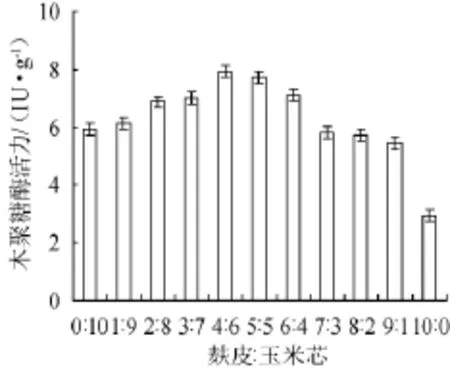
圖3 碳源配比對康氏木霉產木聚糖酶的影響Fig.3 Effect of carbon sources ratio on xylanase production by Trichoderma koningii
由圖3可知,木聚糖酶活力隨麩皮含量的增加呈先上升后下降趨勢,麩皮與玉米芯的質量比為4∶6時木聚糖酶活力達到最高(7.95 IU/g),隨著麩皮含量的增大,培養基易結塊,透氣性較差,酶活力逐漸下降。從酶解產物薄層色譜分析(圖4)來看,麩皮/玉米芯粉質量比為2∶8時,雖然酶活力只有6.89 IU/g,但所含低聚木糖成分較多,木二糖至木五糖的斑點較深,說明低聚木糖濃度較高。因此選擇麩皮與玉米芯粉質量比為2∶8作為最佳碳源配比。
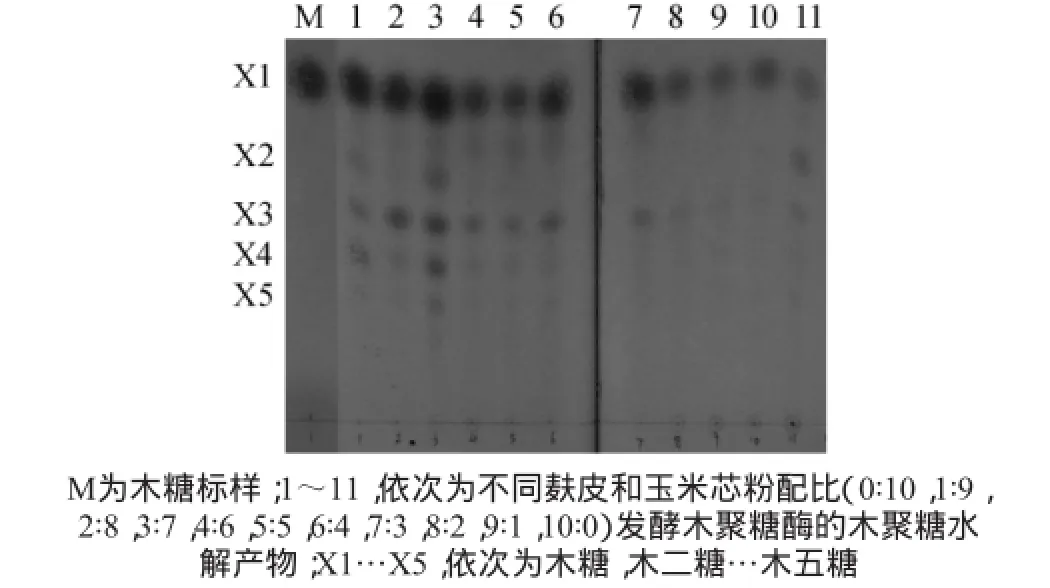
圖4 木聚糖酶反應樣品的薄層層析結果Fig.4 Results of thin layer chromatography of samples by xylanase reaction
2.4氮源條件的確定
2.4.1不同氮源對康氏木霉固體發酵木聚糖酶的影響
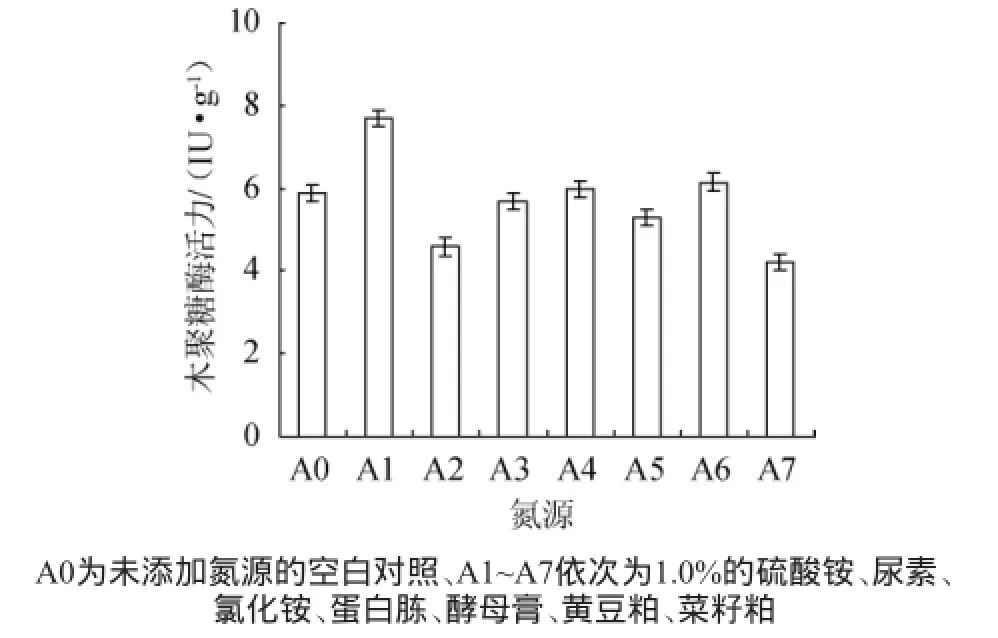
圖5 不同氮源對康氏木霉產木聚糖酶的影響Fig.5 Effect of different nitrogen sources on xylanase productionbyTrichoderma koningii
在碳源為麩皮/玉米芯粉比例為2∶8(g∶g)的條件下,分別添加1.0%的硫酸銨、尿素、氯化銨、蛋白胨、酵母膏、黃豆粕、菜籽粕,考察氮源對木聚糖酶的影響,結果見圖5。從圖5可以看出,康氏木霉以硫酸銨為氮源進行固體發酵時,木聚糖酶活力達到7.68 IU/g,明顯高于其他氮源和對照。
2.4.2最適氮源添加量的優化
以麩皮和玉米芯(2∶8)為培養基碳源,考察質量分數為0、0.25%、0.50%、1.00%、1.50%、2.00%、2.50%、3.00%、4.00%、5.00%的硫酸銨對木聚糖酶的影響,結果見圖6。從圖6可以看出,當硫酸銨添加量達到2.50%時,木聚糖酶活力為9.04 IU/g,之后隨著硫酸銨添加量增加,木聚糖酶活力明顯下降。因此,選擇添加量硫酸銨添加量1.50%~2.50%進行正交試驗。
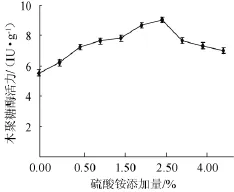
圖6 不同硫酸銨添加量對康氏木霉產木聚糖酶的影響Fig.6 Effect of different(NH4)2SO4addition on xylanase production byTrichoderma koningii
2.5培養基料水比的優化
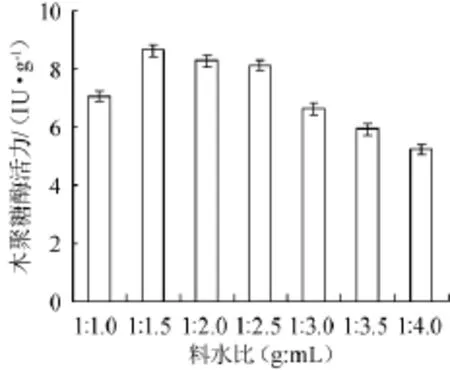
圖7 料水比對康氏木霉產木聚糖酶的影響Fig.7 Effect of material to water ratio on xylanase production byTrichoderma koningii
以麩皮和玉米芯(2∶8)為碳源,2.5%硫酸銨為氮源,將培養基固料與水配制成1∶1.0,1∶1.5,1∶2.0,1∶2.5,1∶3.0,1∶3.5,1∶4.0(g∶mL)不同質量比進行固體發酵,結果見圖7。從圖7可知,料水比為1∶1.0(g∶mL)時,木聚糖酶活力降低,原因可能為含水量過低,無法滿足霉菌生長所需的濕度,造成生物量較低,從而影響其產酶量;料水比為1∶1.5,1∶2.0,1∶2.5(g∶mL)這三個比例條件下酶活力較高,結果較為接近,料水比為1∶1.5(g∶mL)時酶活力最高(8.62 IU/g);隨著含水量的繼續增加,酶活力下降明顯。因此選擇固料與水質量比1∶1.5(g∶mL)為康氏木霉固體發酵產木聚糖酶的最適料水比,由于在料水比1∶1.0(g∶mL)條件下酶活力較低,因此正交試驗選擇料水比為1∶1.5~1∶2.5(g∶mL)進行正交試驗。
2.6培養基初始pH值的確定
將培養基起始pH值分別調至3.0,4.0,5.0,6.0,7.0,8.0,接種康氏木霉發酵木聚糖酶,結果見圖8。從圖8可知,培養基起始pH值對康氏木霉固體發酵產木聚糖酶的影響并不顯著,木聚糖酶活力處于7.5~8.5 IU/g,發酵曲最終pH值維持在5.0~5.5,說明菌株康氏木霉具有一定的調節pH的能力。由于培養基自然pH值為5.5~6.2,因此,康氏木霉固體發酵產木聚糖酶的最適初始pH值無需調整,選擇自然狀態。
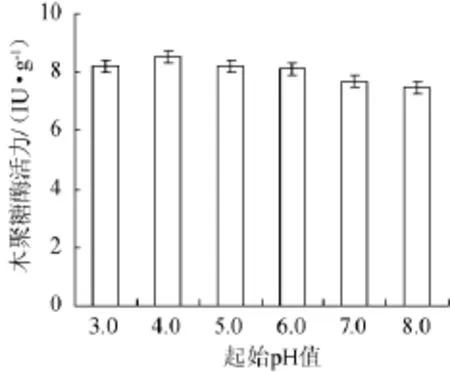
圖8 培養基初始pH值對康氏木霉產木聚糖酶的影響Fig.8 Effect of initial pH on xylanase production by Trichoderma koningii
2.7金屬離子及添加量對康氏木霉產木聚糖酶的影響
在上述試驗優化后的培養基基礎之上,以不添加任何金屬離子為空白對照(CK),其他試驗組分別添加0.30%的MnSO4、CuSO4、MgSO4、KH2PO4、Fe2(SO4)3、CaCl2,考察不同金屬離子對康氏木霉產木聚糖酶的影響,結果見圖9。從圖9可知,Mn2+具有明顯的促進作用,Mg2+,K+,Ca+的促進作用并不明顯,而Cu2+則具有明顯的抑制作用。
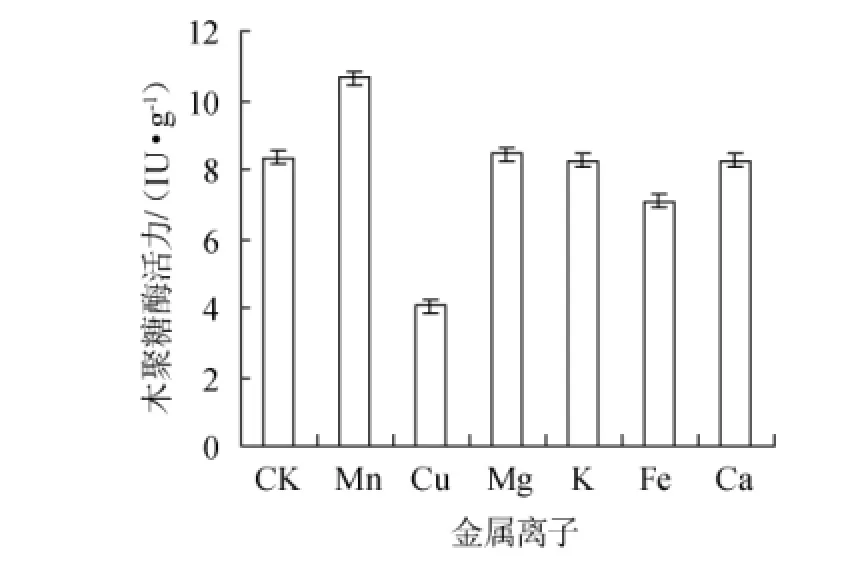
圖9 金屬離子對康氏木霉產木聚糖酶的影響Fig.9 Effect of metal ions on xylanase production by Trichoderma koningii
根據上述試驗結果,以麩皮和玉米芯(2∶8)為培養基干料,添加2.5%硫酸銨,料水比為1∶1.5(g∶mL),再分別添加0,0.10%,0.30%,0.50%,0.70%,0.90%的MnSO4,考察Mn2+添加量對康氏木霉固態發酵產木聚糖酶的影響,結果見圖10。從圖10結果可知,隨著Mn2+添加量的增加,木聚糖酶活力也逐漸升高,添加量為0.50%時酶活力達到最大(11.72 IU/g),繼續添加后,酶活力迅速下降。因此,添加0.50%的Mn2+對康氏木霉固體發酵產木聚糖酶的促進作用最強。
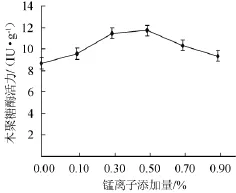
圖10 錳離子添加量對康氏木霉產木聚糖酶的影響Fig.10 Effect of different Mn2+concentration on xylanase production byTrichoderma koningii
2.8正交試驗分析
根據單因素試驗結果,選擇對康氏木霉固體發酵產木聚糖酶影響較為顯著的4個因素:Mn2+添加量,硫酸銨添加量,發酵時間,料水比為評價因素,以木聚糖酶活力為評價指標進行L9(34)的正交試驗,結果見表1,方差分析見表2。
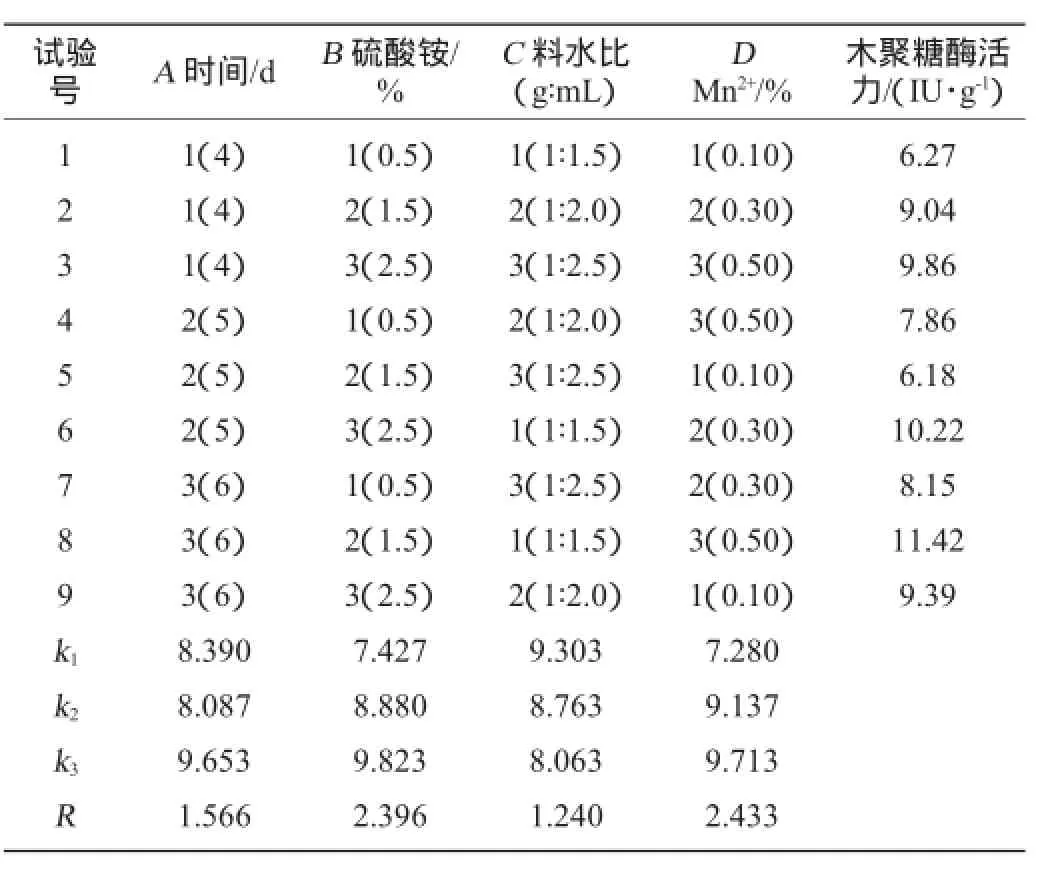
表1 發酵條件正交試驗結果與分析Table 1 Results and analysis of orthogonal experiments for fermentation conditions optimization
從表1可知,通過R值的大小(RD>RB>RA>RC)分析,康氏木霉固體發酵產木聚糖酶的影響因素由強到弱依次為Mn2+添加量,硫酸銨添加量,發酵時間,料水比;從k值分析可以得出最佳發酵條件組合為A3B3C1D3,即發酵時間為6 d,硫酸銨添加量為2.5%,料水比為1∶1.5(g∶mL),Mn2+添加量為0.50%。因此,得到最佳固體發酵條件為:麩皮與玉米芯質量比為2∶8,硫酸銨2.5%,MnSO40.50%,料水比1∶1.5(g∶mL),pH自然,培養溫度30℃,發酵時間6 d。在此條件下,進行發酵重復驗證試驗,木聚糖酶活力可達11.98 IU/g。
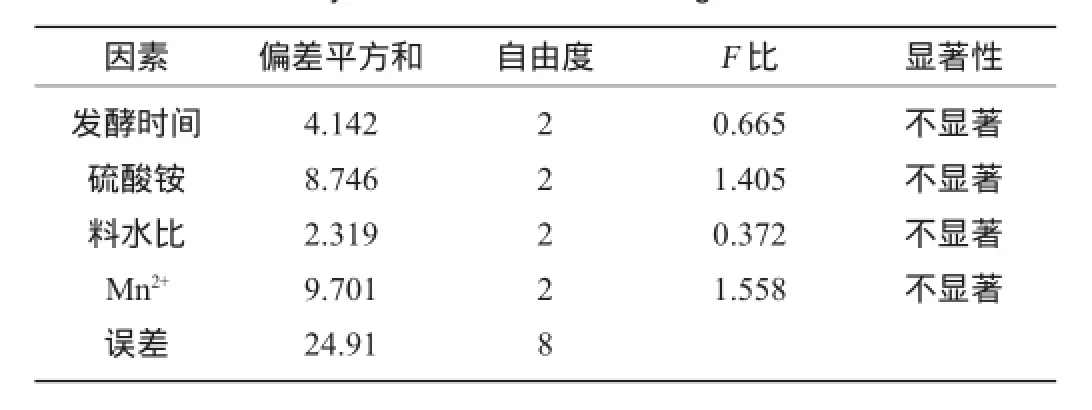
表2 正交試驗結果方差分析Table 2 Analysis of variance for orthogonal tests result
由表2可知,各因素對結果影響均不顯著。
3 結論
該試驗通過優化康氏木霉實驗室固體發酵產木聚糖酶的工藝,有效提高了木聚糖酶活力,所產木聚糖酶能夠將木聚糖底物水解生成含有2~5個木糖分子的低聚木糖。試驗過程中發現,僅有碳源因素對木聚糖酶水解產物種類存在影響,而在其他因素(氮源、培養基初始pH值、料水比、金屬離子)變化條件下,薄層色譜圖均相似,能夠產生5種左右的低聚木糖,原因可能為麩皮和玉米芯中含有木聚糖成分,對木聚糖酶存在一定的誘導作用[13-14]。目前,固態發酵產木聚糖酶的菌種研究多集中在黑曲霉方面,對康氏木霉的研究很少。前期研究發現康氏木霉有復雜的纖維素酶系,可以通過誘導調控獲得高產量的纖維素酶[15]。康氏木霉的木聚糖酶酶系及組分發酵有望通過調控獲得,但其精細調控和每個組分酶學特性有待深入研究。該實驗使用的玉米芯,既是發酵原料,也是潛在的木聚糖水解底物,這對農業秸稈廢棄物的深度資源化,發酵渣的肥料化,減少焚燒引起的環境污染,具有很好的現實意義和市場前景。
[1]COLLINS T,GERDAY C,FELLER G.Xylanases,xylanases families and extremophilic xylanases[J].FEMS Microbiol Rev,2005,29(1):3-23.
[2]高艷秀,陳復生,丁長河.微生物木聚糖酶及其應用[J].中國釀造,2012,31(3):10-12.
[3]倪大偉,李瑞云,白愛枝,等.黑曲霉木聚糖酶的同源表達及其高產木聚糖酶發酵條件研究[J].食品與發酵工業,2015,41(4):48-53.
[4]萬紅貴,武振軍,蔡恒,等.微生物發酵產木聚糖酶研究進展[J].中國生物工程雜志,2010,30(2):141-146.
[5]葉世超,薛婷,何文錦,等.木聚糖酶的應用及其研究進展[J].中國釀造,2013,32(7):8-10.
[6]BEG Q K,KAPOOR M,MAHAJAN L.Microbial xylanases and their industrial applications:a review[J].Appl Microbiol Biot,2001,56(3): 326-338.
[7]YUAN X,WANG J,YAO H.Feruloyl oligosaccharides stimulate the growth of bifidobacterium[J].Bifidum Anaerobe,2005,11:225-229.
[8]丁強.飼用木聚糖酶的研究進展及應用[J].制漿造紙工藝,2010,41(4):18-21.
[9]KIM J C,SIMMINS P H,MULLAN B P,et al.The digestible energy value of wheat for pigs,with special reference to the post-weaned animal[J]. Anim Feed Sci Tech,2005,122(3-4):257-287.
[10]李敏康,張帆,宋宏新.木霉固態發酵麥麩制備低聚木糖的初步研究[J].中國釀造,2012,31(12):60-63.
[11]GHOSE T K.Measurement of cellulase activities[J].Pure Appl Chem,1987,59(2):257-268.
[12]LIN Y S,CHEN G G,LING M,et al.A method of purification,identification and characterization of β-glucosidase fromTrichoderma koningii AS3.2774[J].J Microbiol Method,2010,83(1):74-81.
[13]SINGH S,MADLALA A M,PRIOR B A.Thermomyces lanuginosus: properties of strains and their hemicellulases[J].FEMS Microbiol Rev,2003,27(1):3-16.
[14]張姝倩.黑曲霉產木聚糖酶固體發酵條件研究[J].農產品加工·學刊,2013(8):5-7,11.
[15]林元山.康氏木霉AS3.2774纖維素酶系的誘導、阻遏、純化及鑒定方法研究[D].南寧:廣西大學博士論文,2010.
Solid-state fermentation conditions for xylanase production byTrichoderma koningii
FENG Bo1,YIN Yi1,2,YI Xudong1,LIN Yuanshan1,2*
(1.College of Bioscience and Biotechnology,Hunan Agricultural University,Changsha 410128,China;2.Changsha Shuidisha Biotechnology Co.,Ltd.,Changsha 410125,China)
By solid-state fermentation,the optimal fermentation process for xylanase production byTrichoderma koningiiwas investigated by single factor experiments and orthogonal experiments as follows∶bran to corncob mass ratio 2∶8 as carbon sources,(NH4)2SO42.5%as nitrogen source,Mn-SO40.25%,solid to liquid ratio 1∶1.5(g∶ml),initial pH in natural state,temperature 30℃and time 6 d.Under this condition,the activity of xylanase byT.koningiireached 11.98 IU/g.The products of xylanase hydrolysis were rich in functional polymeric xylose bonded with 2-5 xylose molecular.
Trichoderma koningii;xylanase;solid-state fermentation;optimization
Q939.9
A
0254-5071(2015)10-0073-05
10.11882/j.issn.0254-5071.2015.10.016
2015-09-07
長沙市科技成果轉化資金項目資助(K1501194-31)
馮波(1990-),女,碩士研究生,研究方向為生物技術與工程。
林元山(1969-),男,副教授,博士,研究方向為發酵工程。
