海州香薷Actin基因片段克隆及表達分析
2015-10-26 08:51:23蔡深文熊治廷劉晨徐仲瑞鄧松強
生物技術通報 2015年2期
蔡深文熊治廷劉晨徐仲瑞鄧松強
(1. 遵義師范學院資源與環境學院,遵義 563002;2. 武漢大學資源與環境科學學院,武漢 430079)
海州香薷Actin基因片段克隆及表達分析
蔡深文1熊治廷2劉晨2徐仲瑞2鄧松強2
(1. 遵義師范學院資源與環境學院,遵義 563002;2. 武漢大學資源與環境科學學院,武漢 430079)
通過克隆海州香薷Actin基因片段并分析其組織表達,為研究海州香薷重金屬抗性相關基因的表達調控奠定基礎。根據GenBank中其他植物Actin基因保守序列設計兼并引物,以海州香薷根總RNA為模板,利用RT-PCR技術分離得到Actin基因片段。序列分析結果表明,海州香薷Actin基因片段長576 bp,編碼192個氨基酸,與其他植物同源基因的氨基酸序列相似性為84%-97%,所克隆的序列為Actin基因的同源片段,將其命名為EhACT,在GenBank中提交序列,獲得登錄號AGT37260。半定量RT-PCR分析結果表明,EhACT在海州香薷的根、莖和葉中表達相對穩定,初步表明其可作為研究海州香薷基因表達的內參基因。
海州香薷;Actin基因;克隆;表達
海州香薷(Elsholtzia haichowensis Sun)是一年生草本植物,屬唇形科,廣泛分布于長江中下游銅礦區和銅污染的土壤中,是一種銅礦指示植物,俗稱“銅草”[1,2]。海州香薷在非銅礦區也有發現[3]。銅礦區的海州香薷由于長期生活在高濃度Cu污染的環境脅迫條件下,可能已經發生了與Cu污染相適應的抗性進化,形成了與非礦區種群不同的抗性生態型[4],通常來自礦區種群的海州香薷對Cu具有較高的吸收富積能力和耐受能力[5]。海州香薷是植物重金屬抗性機理研究領域的理想植物材料,從抗性相關基因表達角度能夠更深入地理解海州香薷的銅抗性機理,對于基因表達量的研究通常需要穩定的內參基因作為標準來衡量,選擇合適的內參基因至關重要。植物肌動蛋白(Actin)在植物細胞形成和物質運輸方面起著重要作用[6],是組成型表達蛋白,高度保守,在基因表達調控中被廣泛作為內參基因[7]。而關于海州香薷Actin基因的研究還未見報道。因此,本研究通過同源克隆方法分離海州香薷Actin基因片段并進行表達分析,以此基因為內參標準,旨在為其他功能基因在海州香薷中的表達調控研究奠定基礎。
1 材料與方法
1.1 材料
1.1.1 試驗材料 海州香薷(E. haichowensis)種子采自湖北大冶銅綠山古銅礦遺址(11453’E,30 05’N,MP)。海州香薷種子用0.3% NaClO溶液浸泡消毒20 min,去離子水沖洗5遍后用去離子水浸泡過夜,使種子充分吸收水分。第2天,將種子轉移至底部鋪有濾紙的塑料碗中,加去離子水,水面浸沒種子的一半左右,置于HSR025型人工氣候箱中黑暗萌發,溫度設定為25℃,濕度75%。約96 h后種子基本全部萌發。隨后采用水培方法培養海州香薷幼苗,待長至4對真葉時剪取新鮮的根尖(約2 cm)用于RNA提取。
1.1.2 試劑 RNA提取使用的Trizol試劑購自Invitrogen公司;PCR反應所用的Ex Taq,dNTPmixture,pMD18-T載體和RNase Inhibitor,均購自TaKaRa公司;反轉錄酶M-MLV RT購自Promega公司;DNA凝膠回收試劑盒為Axygen公司產品;大腸桿菌TOP10感受態細胞購自天根生化科技有限公司;其他生化試劑購自丁香園生物技術有限公司,引物由上海博道生物技術公司合成,序列測定由華大基因完成。
1.2 方法
1.2.1 海州香薷總RNA提取 采用Trizol法提取總RNA,結果用1.2%瓊脂糖凝膠電泳檢測,紫外分光光度計測定總RNA濃度及純度,-80℃超低溫冰箱保存備用。
1.2.2 RT-PCR擴增 首先以海州香薷根部總RNA為模板,反轉錄酶與引物Oligo(dT)18引導cDNA第一鏈的合成。0.2 mL離心管中依次加入12.5 μL總RNA,3 μL Oligo(dT)18,11 μL DEPC處理的水,70℃溫浴5 min,迅速置于冰上冷卻,微離心收集溶液至離心管底部,再依次加入5×M-MLV RT Buffer 10 μL,RNase Inhibitor 1.5 μL,dNTP Mixture(2.5 mmol/L each) 10 μL,M-MLV RT 2 μL,用移液槍吸打混勻,于PCR儀上42℃反應60 min,70℃溫浴10 min失活反轉錄酶,冰上分裝后-20℃保存備用。根據其他物種Actin基因序列,設計其上游引物PrimerF 5'-TGTGGTCCTTTTGTGTCATT-3'和下游引物PrimerR:5'-CTCCTTGCTCATCCTGTCAGC-3',以反轉錄所得的cDNA為模板,進行PCR反應,反應體系為20 μL:10× Ex Taq Buffer(Mg2+plus) 2 μL,dNTP Mixture(2.5 mmol/L each) 1 μL,PrimerF 1 μL,PrimerR 1 μL,cDNA 1 μL,Ex Taq 0.2 μL,ddH2O 13.8 μL。PCR反應程序為:94℃預變性3 min;94℃變性30 s,55℃退火30 s,72℃延伸45 s,32個循環;72℃延伸5 min。PCR產物用1.2%瓊脂糖凝膠電泳檢測。
1.2.3 陽性克隆鑒定 目的片段按照DNA凝膠回收試劑盒操作說明進行切膠回收純化,連接至pMD18-T載體,轉化TOP10感受態細胞,擴大培養后提取質粒DNA,進行PCR檢測,選取帶有目的片段的重組質粒測序。
1.2.4 序列分析 使用NCBI網站的BLAST在線軟件(http://www.ncbi.nlm.nih.gov/blast)進行海州香薷Actin基因片段的同源性比對分析與相似性搜索。使用ClustalX軟件將海州香薷和其他物種Actin基因的氨基酸序列進行比對[8]。
1.2.5 半定量RT-PCR分析Actin基因表達 根據已獲得的Actin基因片段序列,設計特異性引物Actin--F(5'-TTCTTTGGCAGGTATTGTTCTCG-3')和ActinR(5'-TTC-CCGCTCGGCTGTAGTTGT-3')用于半定量RT-PCR反應。分別提取海州香薷的根、莖和葉的總RNA,并反轉錄生成cDNA,以該cDNA為模板進行PCR反應,反應體系為20 μL:10×Ex Taq Buffer(Mg2+plus) 2 μL,dNTP Mixture(2.5 mmol/L each)1 μL,PrimerF 1 μL,PrimerR 1 μL,cDNA 1 μL,Ex Taq 0.2 μL,ddH2O 13.8 μL。PCR反應程序為:94℃預變性3 min;94℃變性30 s,55℃退火30 s,72℃延伸30 s,28個循環;72℃延伸5 min。PCR產物用1.2%瓊脂糖凝膠電泳檢測。
2 結果
2.1 總RNA提取
海州香薷根中的總RNA經1.2%瓊脂糖凝膠電泳檢測(圖1)顯示,呈現3條清晰的條帶,其中28S rRNA的亮度約為18S rRNA的兩倍,5S rRNA亮度較低,無明顯降解,紫外分光光度計測定RNA樣品OD260/OD280的平均值為1.91,表明所提取的總RNA質量較好,可用于反轉錄反應。
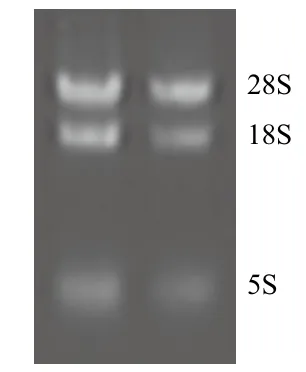
圖1 海州香薷根部總RNA提取
2.2 RT-PCR擴增和克隆
以第一鏈cDNA為模板,用引物PrimerF和PrimerR進行PCR擴增,獲得一條約550 bp的條帶,與預期大小相似,將目的條帶進行凝膠回收,隨后連接至pMD18-T載體上,轉化大腸桿菌TOP10。從轉化平板上隨機挑取5個白斑進行菌液PCR檢測,結果(圖2)表明5個均為陽性克隆。
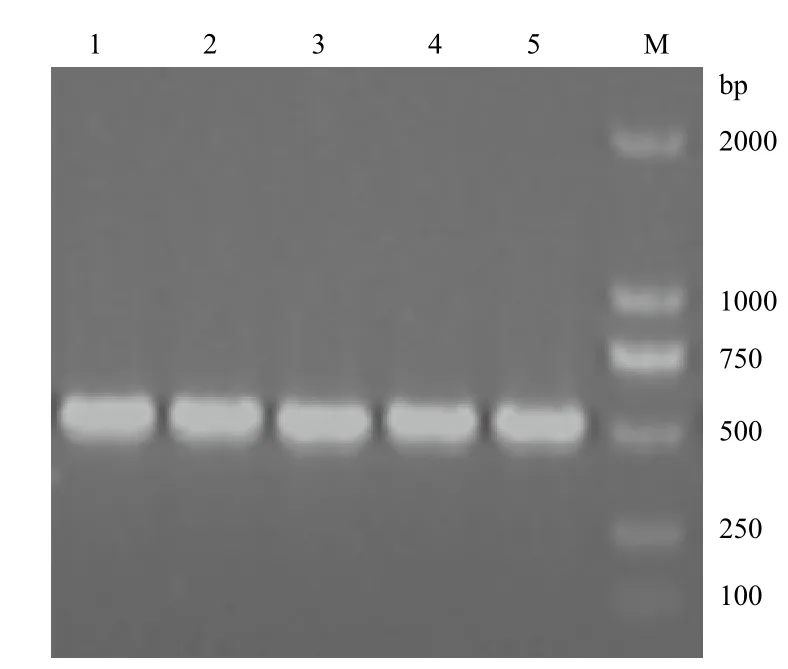
圖2 陽性克隆的PCR鑒定
2.3 測序結果與序列分析
將5個克隆進行測序,其測序結果完全一致,得到576 bp的序列,編碼192個氨基酸(圖3)。Blast結果表明,海州香薷與其他植物Actin基因的核苷酸序列相似性為78%-91%,氨基酸序列相似性為84%-97%,其中與唇形科植物丹參(Salvia miltiorrhiza)(GenBank:ADK11998)的相似性最高,達97%。將EhACT與其他8種植物Actin基因的氨基酸序列進行多重比對(圖4),EhACT具有153個保守氨基酸,非保守氨基酸為38個,表明所克隆的序列片段為Actin基因片段,將其命名為EhACT,在GenBank中提交序列,獲得登錄號AGT37260。

圖3 海州香薷Actin基因片段的核苷酸序列及推導的氨基酸序列
2.4 Actin基因的組織表達分析
通過特異性引物ActinF和ActinR克隆海州香薷不同組織中的Actin基因片段,將目的片段進行測序驗證,結果顯示其長度為184 bp,屬于EhACT基因序列。半定量RT-PCR分析結果(圖5)顯示,海州香薷Actin基因在根、莖和葉組織中的表達相對穩定,不具有組織表達特異性。
3 討論
本研究通過同源克隆方法分離得到海州香薷的Actin基因片段序列,序列分析結果表明EhACT與其他植物Actin基因的序列相似性較高,氨基酸序列高度保守,表明所克隆的序列為Actin基因片段序列。大量研究表明,Actin基因在核苷酸和氨基酸水平都具有高度的保守性和同源性[9-11],這種高度保守性是由于植物肌動蛋白起源于一個共同祖先,在進化過程中通過復制和變異而被保留下來[12,13]。因此,Actin基因在植物生命活動過程中起著十分重要的作用,克隆海州香薷的Actin基因對于認識海州香薷在銅脅迫環境下的進化過程也具有重要意義。Actin基因的重要性不僅表現在生命活動過程中,且由于該基因的表達屬于組成型表達,在研究其他基因的表達調控過程中也起著重要作用。

圖4 海州香薷Actin基因氨基酸序列片段與其他植物Actin基因的氨基酸序列多重比對
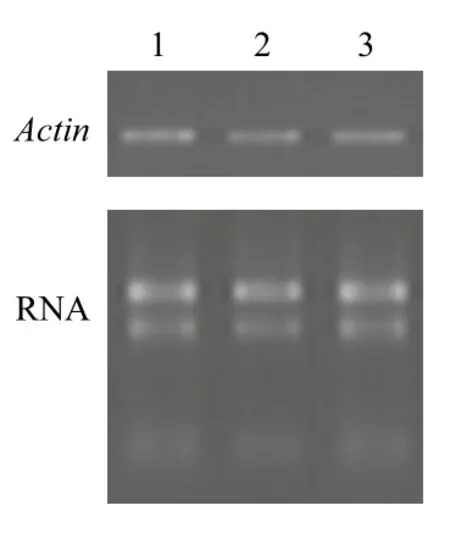
圖5 海州香薷Actin基因的組織表達
植物基因表達分析研究中比較常用的內參對照基因即管家基因主要有肌動蛋白基因(ACT)、甘油醛-3-磷酸脫氫酶基因(G3PDH/GAPDH)、18S和 28S核糖體RNA基因(18S rRNA、28S rRNA)及肽鏈延伸因子(ef1-α)基因等[14-16]。其中肌動蛋白是真核細胞中主要的胞質微絲骨架組分之一,是植物基因表達研究中廣泛使用的內參基因[17-19]。雖然上述管家基因一直被作為內參基因來標準化目的基因,但最近許多研究表明,不同管家基因在不同細胞類型和不同生理狀態下的表達并不是恒定不變的,如Xu等[20]分析了GAPDH、ACT、TUA、TUB、18SrRNA、RPII、EF-1b和TEF2基因在蘿卜不同組織、品種和發育時期的表達穩定性,其中只有ACT、RPII 和TEF2基因的表達相對穩定,可作為內參基因。Podevin等[21]通過實時熒光定量PCR分析了芭蕉(Musa)GAPDH等9個基因的表達,結果表明只有EF1、TUB 和ACT可以在不同組織穩定表達。因此,在基因表達研究中需選擇合適的管家基因作為內參基因,本試驗克隆了一條576 bp長的海州香薷Actin基因片段,并通過半定量RT-PCR分析發現其在不同組織部位的表達相對穩定,初步表明其可作為內參基因,為研究其他基因特別是重金屬抗性相關基因在海州香薷體內的表達和調控奠定了基礎。本試驗僅研究了EhACT基因的組織表達特異性,其他管家基因在不同條件下是否能夠穩定表達,以及與EhACT基因相比是否更加穩定,還有待進一步研究證實。
4 結論
本研究通過同源克隆得到海州香薷Actin基因片段(EhACT,GenBank登錄號AGT37260),片段長576 bp,編碼192個氨基酸,與其他植物同源基因的氨基酸序列相似性為84%-97%。半定量RT-PCR分析結果表明EhACT在海州香薷的根、莖和葉中表達相對穩定,可作為研究海州香薷基因表達的內參基因。
[1] 柯文山, 席紅安, 楊毅, 等. 大冶銅綠山礦區海州香薷(Elsholtzia haichowensis)植物地球化學特征分析[J]. 生態學報, 2001,21(6):907-912.
[2]謝學錦, 徐邦梁. 銅礦指示植物——海州香薷[J]. 地質學報,1952, 32(4):360-368.
[3]Liu J, Xiong ZT. Differences in accumulation and physiological response to copper stress in three populations of Elsholtzia haichowensis S. [J]. Water Air Soil Pollut, 2005, 168(1-4):5-16.
[4] 熊治廷. 植物抗污染進化及其遺傳生態學代價[J]. 生態學雜志,1997, 16(1):53-57.
[5]Yang MJ, Yang XE, R?mheld V. Growth and nutrient composition of Elsholtzia splendens Nakai under copper toxicity [J]. J Plant Nutr,2002, 25(7):1359-1375.
[6] 陳穎, 王剛, 趙俊霞. 高等植物體內的肌動蛋白[J]. 生物學通報, 2003, 38(2):13-15.
[7]Meagher RB, McKinney EC, Kandasamy MK. Isovariant dynamics expand and buffer the responses of complex systems:the diverse plant actin gene family [J]. Plant Cell, 1999, 11(6):995-1005.
[8]Thompson JD, Gibson TJ, Plewniak F, et al. The CLUSTAL_ X windows interface:flexible strategies for multiple sequence alignment aided by quality qnalysis tools [J]. Nucl Acid Res, 1997,25(24):4876-4882.
[9]Pollard TD, Cooper JA. Actin, a central player in cell shape and movement [J]. Science, 2009, 326(5957):1208-1212.
[10]Erickson HP. Evolution of the cytoskeleton [J]. Bioessays, 2007,29(7):668-677.
[11]Carlon E, Dkhissi A, Malki ML, et al. Stability domains of actin genes and genomic evolution [J]. Phys Rev E, 2007, 76(5):051916.
[12]Skaloud P, Peksa O. Evolutionary inferences based on ITS rDNA and actin sequences reveal extensive diversity of the common lichen alga Asterochloris(Trebouxiophyceae, Chlorophyta) [J]. Mol Phylogenet Evol, 2010, 54(1):36-46.
[13] Baruaa B, Pamulab MC, Hitchcock-DeGregori SE. Evolutionarily conserved surface residues constitute actin binding sites of tropomyosin [J]. Proc Nat Acad Sci USA, 2011, 108(25):10150-10155.
[14]牙庫甫江·阿西木, 關波, 張富春. 植物基因表達轉錄分析中內參基因的選擇與應用 [J]. 生物技術通報, 2011(7):7-11.
[15] Lopez-Pardo R, de Galarreta JIR, Ritter E. Selection of housekeeping genes for qRT-PCR analysis in potato tubers under cold stress[J]. Mol Breeding, 2013, 23(1):39-45.
[16] Eisenberg E, Levanon EY. Human housekeeping genes, revisited[J]. Trends Genet, 2013, 29(10):569-574.
[17]Reid K, Olsson N, Schlosser J, et al. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development [J]. BMC Plant Biol, 2006, 6(1):27.
[18]Hruz T, Wyss M, Docquier M, et al. RefGenes:identification of reliable and condition specific reference genes for RT-qPCR data normalization [J]. BMC Genomics, 2011, 12(1):156.
[19]Yan J, Yuan F, Long G, et al. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus [J]. Mol Biol Rep, 2012, 39(2):1831-1838.
[20]Xu Y, Zhu X, Gong Y, et al. Evaluation of reference genes for gene expression studies in radish(Raphanus sativus L.) using quantitative real-time PCR [J]. Biochem Biophys Res Commun,2012, 424(3):398-403.
[21]Podevin N, Krauss A, Henry I, et al. Selection and validation of reference genes for quantitative RT-PCR expression studies of the non-model crop Musa [J]. Mol Breeding, 2012, 30(3):1237-1252.
(責任編輯 李楠)
Cloning and Expression Analysis of Actin Gene Gragment from Elsholtzia haichowensis
Cai Shenwen1Xiong Zhiting2Liu Chen2Xu Zhongrui2Deng Songqiang2
(1. School of Resource and Environment,Zunyi Normal College,Zunyi 563002;2. School of Resource and Environmental Sciences,Wuhan University,Wuhan 430079)
Cloning and expression analysis of Actin gene fragment from Elsholtzia haichowensis would provide foundation for the study of gene expression and regulation of heavy metal resistance related genes. Degenerate primers were designed based on the conserved sequences of the Actin genes from other plants. Total RNA was extracted from the root of E. haichowensis. An Actin gene fragment was separated by reverse transcription polymerase chain reaction(RT-PCR). The sequence analysis results revealed that Actin gene fragment from E. haichowensi contains 576 bp, encoding a protein of 192 amino acids. Homology comparison with other plants Actin gene sequences in the GenBank showed that it shared 84%-97% amino acid sequence homology with other plants. The cloned sequence was Actin gene fragment. It was named as EhACT and was registered into GenBank(accession number:AGT37260). Semi-quantitative PCR assays indicated that the expression of EhACT in root, stem and leaf of E. haichowensis was relatively stable, suggesting that EhACT can be used as the reference to analyze the gene expression in E. haichowensis.
Elsholtzia haichowensis;Actin gene;cloning;expression
10.13560/j.cnki.biotech.bull.1985.2015.02.016
2014-07-17
國家自然科學基金項目(30870365,31270432)
蔡深文,男,博士,講師,研究方向:環境生物學;E-mail:caishenwen@163.com
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
