不同鹽質量濃度四川泡菜腐敗前后微生物的分析比較研究
2015-10-29 02:13:40蔣云露楊建濤車振明
食品科學 2015年13期
關鍵詞:質量
王 猛,蔣云露,楊建濤,常 偉,車振明,陳 功,饒 瑜,*
(1.西華大學生物工程學院,四川 成都 610039;2.通威三文魚研究所,四川 都江堰 611800;3.四川省食品發酵工業研究設計院,四川 溫江 611130)
不同鹽質量濃度四川泡菜腐敗前后微生物的分析比較研究
王猛1,蔣云露1,楊建濤1,常偉2,車振明1,陳功3,饒瑜1,*
(1.西華大學生物工程學院,四川 成都 610039;2.通威三文魚研究所,四川 都江堰 611800;3.四川省食品發酵工業研究設計院,四川 溫江 611130)
研究不同鹽質量濃度四川泡菜正常發酵后與發生腐敗后鹽鹵中微生物的數量、分布及變化。利用不同的選擇培養基對3 個鹽質量濃度四川泡菜腐敗前后鹽鹵中的微生物進行分離純化,得到168 株菌株,并經16S rDNA和18S rDNA分子生物學鑒定其種屬。結果表明:在不同鹽質量濃度泡菜正常發酵過程中,以植物乳桿菌和戊糖乳桿菌為主的乳酸菌主導了泡菜發酵過程。泡菜腐敗時,不同鹽質量濃度泡菜中的腐敗微生物以芽孢桿菌和酵母為主,其中枯草芽孢桿菌和塔克斯假絲酵母在不同鹽質量濃度泡菜腐敗時均占一定比例;40 g/L鹽質量濃度泡菜鹽鹵以甲級營養型芽孢桿菌、地衣芽孢桿菌、庫德里阿茲威畢赤酵母和熱帶念珠酵母為主,60 g/L鹽質量濃度泡菜鹽鹵以短小芽孢桿菌、克魯維畢赤酵母和熱帶念珠酵母為主,80 g/L鹽質量濃度泡菜鹽鹵以奇異變形桿菌、解淀粉芽孢桿菌、白地霉、地衣芽孢桿菌、庫德里阿茲威畢赤酵母以及克魯維畢赤酵母為主。腐敗泡菜鹽鹵表層的膜璞由細菌和真菌共同形成,且鹽質量濃度越低,細菌所占的比例越高。
四川泡菜;鹽質量濃度;正常發酵;腐敗;優勢菌群
泡菜是一類以各種新鮮蔬菜為原料,在一定鹽濃度溶液中經乳酸菌自然發酵得到的一種蔬菜制品。四川泡菜歷史悠久,以味道咸酸、口感脆生、色澤鮮亮、香味撲鼻而著稱,被稱為“川菜之骨”[1],泡菜風味獨特,不僅是佐餐佳品,而且還具有刺激食欲,改善腸道環境,促進胃腸健康等功能,廣受消費者喜愛[2-5]。但是在泡菜的制作過程中,由于泡菜原料未經過滅菌處理,加之其微生物發酵的自發性,在泡菜正常發酵完成后鹽鹵中仍保留了大量微生物,在泡菜貯藏的過程中會繼續進行生長發酵,造成泡菜的過熟或腐敗。發酵或貯藏過程中條件控制不當也會造成大腐敗微生物的繁殖引起產品的腐敗變質。家庭自制泡菜因為時常加入新的蔬菜或添加白酒等方法,在一定程度上可以避免泡菜的過熟或腐敗問題。而工業化泡菜生產中如何控制腐敗微生物的生長,保證產品的品質及安全,進而推動四川傳統泡菜的產業化發展是一個亟待解決的問題[6]。研究四川泡菜的微生物區系,分離鑒定引起泡菜腐敗的微生物,將對泡菜腐敗控制起到一定指導作用。
本研究利用不同選擇培養基分離純化不同鹽質量濃度四川泡菜正常發酵后與貯藏腐敗后的各種微生物,并進行分子生物學菌種鑒定。對比泡菜正常發酵后與貯藏腐敗后的微生物變化,分析正常發酵的泡菜中存在的潛在腐敗微生物,探討不同鹽質量濃度對腐敗微生物及腐敗現象的影響。
1 材料與方法
1.1材料與試劑
四川泡菜:原料為白蘿卜、胡蘿卜及空心菜。將原料洗干凈,晾干。白蘿卜、胡蘿卜切成片狀;空心菜切成條狀;將白蘿卜、胡蘿卜與白菜混在一起裝入1.7 L土陶壇子,將壇子中的蔬菜壓實,裝到土陶壇子總體積的2/3刻度處,白蘿卜、胡蘿卜與白菜的質量比為6∶2∶2。之后在壇子中加入鹽質量濃度分別為40、60、80 g/L的冷開水,加到土陶壇子總體積的4/5刻度處,料液比大概在2∶1。液封防止空氣進去壇子中,置于室溫發酵。每個鹽質量濃度平行2 組實驗。
1.2儀器與設備
臺式冷凍離心機 美國Beckman公司;PCR儀、凝膠成像系統 美國Bio-Rad公司;電泳儀 北京六一儀器廠;立式壓力蒸汽滅菌鍋 上海申安醫療器械廠。
1.3方法
1.3.1泡菜理化指標的測定
泡菜中亞硝酸鹽是參照GB 5009.33—2010《食品安全國家標準 食品中亞硝酸鹽與硝酸鹽的測定方法》進行測定,氨基態氮是參照GB/T 12143—2008《飲料通用分析方法》進行測定。
1.3.2微生物計數
參照GB 4789.2—2010 《食品安全國家標準 食品微生物學檢驗 菌落總數測定》的方法制備樣品稀釋液,選取適當的稀釋度涂布于培養基。NA瓊脂培養基用于細菌總數計數;MRS瓊脂培養基用于乳酸菌的篩選,培養基中添加終質量濃度為35 μg/mL的放線菌酮;VRBA瓊脂培養基用于分離大腸菌群;虎紅瓊脂培養基用于霉菌和酵母菌的分離和計數。NA、VRBA瓊脂培養基于37 ℃培養箱培養24~36 h,MRS、虎紅瓊脂培養基于30 ℃培養先培養24~48 h。
1.3 統計學分析 用SPSS22.0統計軟件,計數資料用%表示,采用χ2檢驗,P<0.05為差異有統計學意義。
1.3.3不同微生物的分離鑒定
從培養基上挑選不同形態的單菌落進行液體培養,使用細菌或真菌DNA提取試劑盒提取總DNA,于1.0%瓊脂糖凝膠電泳檢驗結果。以提取的總DNA為模板,細菌以Eu27F(5'-AGAGTTTGATCCTGGCTCAG-3')和1490R(5'-GGTTACCTTGTTACGACTT-3')為引物,于95 ℃ 5 min;95 ℃ 1 min,50 ℃ 1 min,72 ℃ 2 min,35 個循環;72 ℃ 10 min條件下進行擴增16S rDNA全長片段。真菌以NS1(5'-GCATATCAATAAGC GGAGGAAAAG-3')和NS4(5'-GGTCCGTGTTTC AAGACGG-3')為引物,于94 ℃ 3 min;94 ℃ 30 s,40 ℃ 1 min,72 ℃ 1 min,30個循環;72 ℃ 10 min條件下進行擴增18S rDNA部分片段。
擴增產物經膠回收純化,送上海華津生物科技有限公司測序,測得的序列與GenBank數據庫進行相似性分析,并通過Clustal X軟件多重比對后用MEGA6.0軟件中的Neighbor-Joining法進行構建系統發育樹。
2 結果與分析
2.1不同鹽質量濃度四川泡菜發酵過程中的理化指標及微生物變化
如圖1所示,發酵至第12天時,40、60、80 g/L鹽質量濃度泡菜pH值都降至4.0甚至4.0以下,一般認為,pH≤4.0時,泡菜正常發酵完成[7]。在此發酵過程中,40、60、80 g/L這3 種鹽質量濃度泡菜鹽鹵中的氨基態氮、亞硝酸鹽的變化情況與余文華[8]、楊瑞[9]等研究一致,氨基態氮含量提高了約2 g/100 g,亞硝酸鹽含量在發酵的第4~5天時達到最大值,在第12天時已下降到初始狀態。圖2所示為12 d發酵過程中,不同鹽質量濃度泡菜鹽鹵中不同微生物的生長變化情況。可以看出,不同鹽質量濃度對四川泡菜的微生物生長產生一定影響。四川泡菜的發酵主要是乳酸菌的發酵過程,也是引起泡菜pH值降低的主要原因。40、60、80 g/L鹽質量濃度泡菜中的乳酸菌數量分別在第10、9、10天達到了最大值5.8×106、6.8×106、6.7×106CFU/mL,而60 g/L鹽更適于乳酸菌的生長(圖2B),這與Suzanne等[10]的研究一致。真菌和大腸菌群在該過程中也有一定生長,在發酵第12天時,3 種鹽質量濃度泡菜的真菌總數都達到了104CFU/mL,但不同鹽質量濃度中真菌生長速率有一定差異(圖2D)。大腸菌群在正常發酵后期達到了104CFU/mL,鹽質量濃度越高,生長速率越快(圖2C)。梭菌在該期間數量很少并無明顯變化(結果未顯示)。
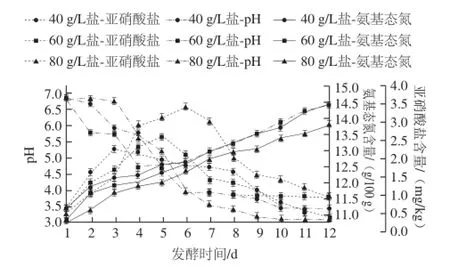
圖1 不同鹽質量濃度泡菜中理化指標動態變化Fig.1 Dynamic changes of physical and chemical indicators in pickles with different salt concentrations
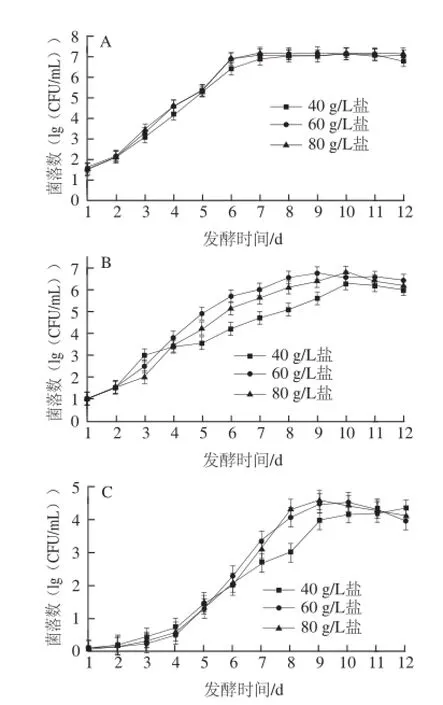

圖2 不同鹽質量濃度四川泡菜發酵過程中不同微生物的生長變化Fig.2 Growth curves of different microorganisms in Sichuan pickles with different salt concentrations
2.2不同鹽質量濃度四川泡菜腐敗后理化及微生物指標
當泡菜表面出現白色膜璞[11-13],鹽鹵出現混濁時,認為泡菜已發生腐敗。40、60、80 g/L鹽質量濃度泡菜分別于第35、47、37天出現腐敗,在鹽鹵表面出現白色生物膜。40 g/L和80 g/L鹽質量濃度泡菜鹵水較60 g/L鹽質量濃度泡菜混濁,已不具有泡菜特有香味,有一定惡臭氣味[7]。泡菜腐敗后理化以及微生物指標如表1、2所示。
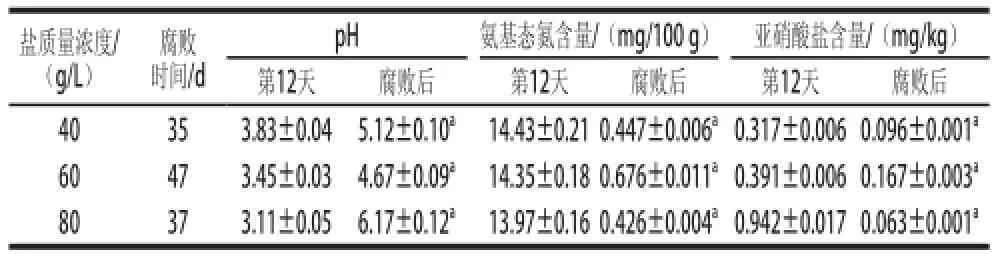
表1 不同鹽質量濃度泡菜正常發酵與腐敗后理化指標Table1 Comparison of physical and chemical indicators in pickles with different salt concentrations before and after spoilage
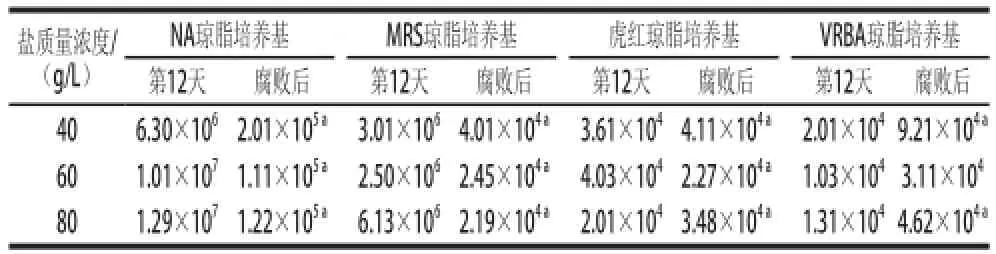
表2 不同鹽質量濃度泡菜正常發酵與腐敗后的微生物數量Table2 Comparison of microbiological indicators in pickles with different salt concentrations before and after spoilage CFU/mL
由表1可知,泡菜腐敗時,鹽鹵的pH值會有所升高,其中80 g/L鹽質量濃度泡菜的pH值變化最大,在腐敗時(第37天)pH值已升至6.17。如表2所示,在泡菜發酵腐敗后,40、60、80 g/L鹽質量濃度泡菜中微生物數量均變化顯著,總數下降了一個數量級,乳酸菌數量均下降了兩個數量級。其中以80 g/L鹽質量濃度泡菜中乳酸菌數量下降最為明顯,由發酵第12天的6.1×106CFU/mL下降至2.2×104CFU/mL。40、60、80 g/L鹽質量濃度泡菜中真菌的數量有一定波動,40 g/L和80 g/L鹽質量濃度泡菜在腐敗后真菌數量分別增加了0.5×104CFU/mL和1.5×104CFU/mL,而60 g/L鹽質量濃度泡菜腐敗后真菌數量下降了1.7× 104CFU/mL。大腸菌群的數量在不同鹽質量濃度泡菜腐敗后均提高了3~4倍,其中40 g/L鹽質量濃度泡菜腐敗后(第35天)大腸菌群的數量達到了9.2×104CFU/mL,為3 個鹽質量濃度泡菜腐敗后大腸菌群數量中最高。
一般認為,在泡菜發酵后期,由于pH值降至4.0左右,乳酸菌的生長在該pH值條件下會受到抑制[14-17],在圖2B中可看出乳酸菌數量在發酵第9~12天時已有所下降,并在泡菜貯藏 的過程中一直受到抑制并出現數量大幅降低(表2)。泡菜發酵后,基質中殘存的糖將被以酵母為主的真菌所利用,也可能利用由乳酸菌發酵產生的乳酸、醋酸等有機酸引起基質pH值的上升[18-19]。本實驗中,真菌在泡菜發酵過程中數量持續增長(圖2D),在貯藏過程中數量也有一定增加(表3),但60 g/L鹽質量濃度泡菜中真菌數量有小幅度下降。Franco等[18]報道在酸黃瓜的腐敗過程中,大腸菌群也是重要的腐敗菌之一,腐敗真菌生長引起的pH值上升和基質的還原為大腸菌群的生長提供了條件。本實驗中,泡菜發酵后期(第9~12天)低pH值條件下,大腸菌群受到抑制,數量有所下降,但在貯藏過程中卻大量生長,腐敗后泡菜中大腸菌群的數量已超過了乳酸菌和真菌,這應該與真菌在腐敗過程中改變泡菜基質條件有關。
pH值主要是來自原料在發酵過程中乳酸菌代謝乳酸和在腐敗后腐敗微生物消耗有機酸。氨基態氮主要來自于原料中蛋白質的水解和微生物的自溶[20]。在泡菜腐敗過程中,pH值的升高的原因是腐敗微生物利用泡菜中的乳酸、醋酸等有機酸以及殘糖來代謝生長[18-19]。在本實驗中,40 g/L和80 g/L鹽質量濃度的泡菜腐敗微生物生長較厲害,有機酸被大量利用,造成pH值上升,所以在腐敗后40 g/L和80 g/L鹽質量濃度泡菜的pH值較60 g/L鹽質量濃度泡菜要高。在泡菜腐敗的過程,氨基態氮的變化是原料中蛋白的水解和微生物的自溶。40 g/L和80 g/L鹽質量濃度泡菜中的腐敗微生物的大量快速生長,造成氨基態氮的破壞,所以氨基態氮的含量減少。
2.3四川泡菜正常發酵后與腐敗后腐敗細菌的分離與比較

表3 不同鹽質量濃度泡菜正常發酵與腐敗后優勢微生物的挑選Table3 Dominant microorganisms in normal and spoiled pickles with different salt concentrations
將3 種鹽質量濃度泡菜在發酵第12天和泡菜腐敗后,分別取樣稀釋并涂布于NA、MRS、VRBA、虎紅4 種瓊脂培養基上并培養,分離培養數量較多的優勢菌株進行分子生物學鑒定,菌株挑選數量與分布如表3所示,共挑選菌株168 株。圖3、4分別為部分菌株的基因組和16S rDNA/18S rDNA瓊脂糖凝膠電泳圖。
圖3 部分真菌(A)、細菌(B、C)基因組瓊脂糖凝膠電泳圖Fig.3 Agarose gel electrophoresis of genomic DNAs from fungi (A) and bacteria (B, C)
圖4 部分菌株PCR擴增18S rDNA(A)、16S rDNA(B)瓊脂糖凝膠電泳圖Fig.4 Agarose gel electrophoresis of PCR-amplified products from 18S rDNA (A) and 16S rDNA (B)
PCR擴增的16S rDNA或18S rDNA片段經測序并BLAST分析及多重比對后,構建發育樹。Devereux等[21]認為當16S rDNA的序列同源性≥97%時可以認為是一個屬,同源性≥98%時則可以認為是同一個種。進行鑒定的168 株菌株分別屬于16 個菌種,包括11 種細菌,分別是枯草芽孢桿菌(Bacillus subtilis,21 株)、地衣芽孢桿菌(Bacillus licheniformis,6 株)、甲級營養型芽孢桿菌(Bacillus methylotrophicus,4 株)、短小芽孢桿菌(Bacillus pumilus,5 株)、蘇金云芽孢桿菌(Bacillus thuringiensis,5 株)、解淀粉芽孢桿菌(Bacillus amyloliquefaciens,7 株)、戊糖乳桿菌(Lactobacillus pentosus,4 株)、植物乳桿菌(Lactobacillus plantarum,5 株)、路德腸桿菌(Enterobacter ludwigii,15 株)、陰溝腸桿菌(Enterobacter cloacae,19 株)和奇異變形桿菌(Proteus mirabilis,7 株);5 種真菌,分別是庫德里阿茲威畢赤酵母(Pichia kudriavzevii,16 株)、白地霉(Galactomyces geotrichum,17 株)、塔克斯假絲酵母(Candida tanzawaensis,13 株)、熱帶念珠酵母(Candida tropicalis,10 株)和克魯維畢赤酵母(Pichia kluyveri var. cephalocereana,14 株)。這些微生物在不同鹽質量濃度泡菜正常發酵第12天和腐敗后樣品中的分布如圖5、6所示。
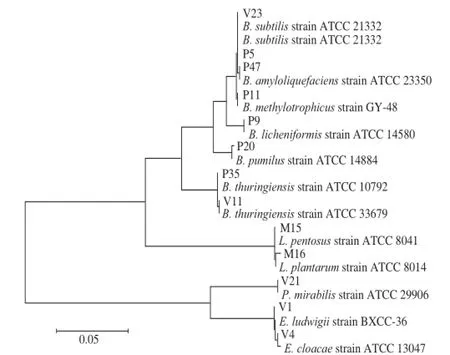
圖5 基于16S rDNA構建的發酵第12天與腐敗后不同鹽質量濃度泡菜鹵水中細菌的系統發育樹Fig.5 Phylogenetic tree based on 16S rDNA sequences of strains from 12-day fermented and spoiled pickle brines with different salt concentrations
由圖5可知,發酵后(第12天)的不同鹽質量濃度泡菜中微生物以乳酸菌為主,主要為植物乳桿菌和戊糖乳桿菌,共占微生物總數的48%左右,這與Yan Pingmei等[22-24]的研究一致。中國泡菜發酵過程中的主要微生物包括植物乳桿菌(43.6%)、戊糖乳桿菌(19.1%)、短乳桿菌(11.0%)和腸系膜明串珠球菌(7.3%)等乳酸菌。在泡菜發生腐敗后,不同鹽質量濃度泡菜中乳酸菌數量下降明顯,由發酵第12天的106CFU/mL下降至104CFU/mL。
由圖5可知,在泡菜發酵(第12天)和泡菜腐敗后,泡菜鹽鹵中均篩選出枯草芽孢桿菌,數量級基本都在105CFU/mL,說明不同鹽質量濃度的泡菜在發酵前(第12天)和腐敗后枯草芽孢桿菌的數量差別不大。在發酵后第12天的泡菜鹽鹵中,大腸菌群以路德腸桿菌、陰溝腸桿菌為主,但泡菜腐敗后沒有發現這兩株菌株。雖然表2中,泡菜腐敗后樣品在VRBA上生長的微生物數量達到104CFU/mL,但卻不是大腸菌群而主要是枯草芽孢桿菌。
地衣芽孢桿菌、甲級營養型芽孢桿菌、短小芽孢桿菌、蘇金云芽孢桿菌、解淀粉芽孢桿菌、路德腸桿菌、陰溝腸桿菌和奇異變形桿菌在腐敗泡菜中出現,這應該是隨著pH值的升高,低pH值對腐敗菌抑制解除的結果。例如Franco等[18]在對黃瓜的二次發酵研究中發現,隨著酸黃瓜pH值的升高,陰溝腸桿菌等腐敗微生物逐漸富集。
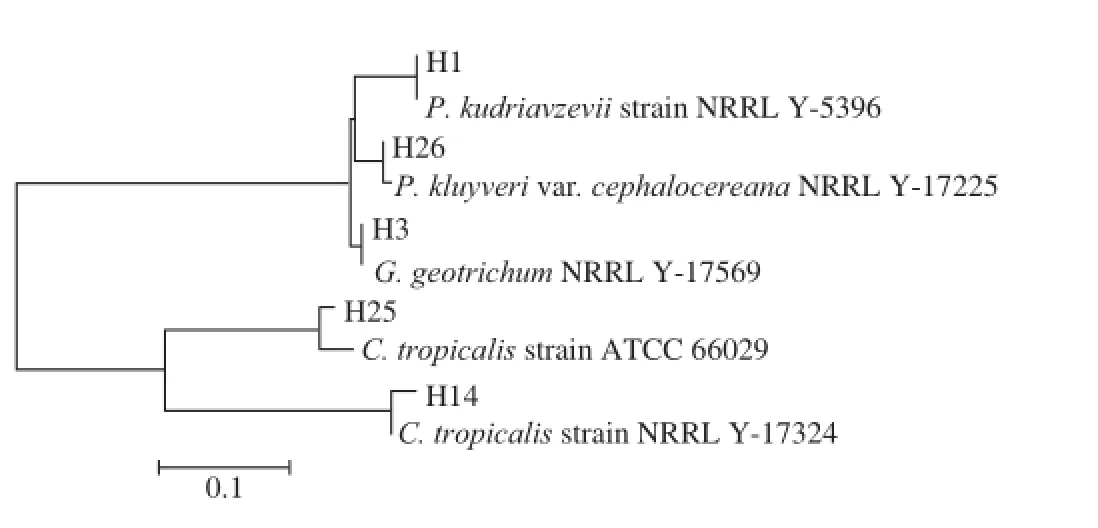
圖6 部分基于18S rDNA構建的發酵第12天與腐敗后不同鹽質量濃度泡菜鹵水中真菌系統發育樹Fig.6 Phylogenetic tree based on 18S rDNA sequences of 12-day fermented and spoiled pickle brines with different salt concentrations
由圖6可知,發酵后(第12天)的泡菜鹽鹵真菌以庫德里阿茲威畢赤酵母、白地霉、塔克斯假絲酵母、熱帶念珠酵母和克魯 維畢赤酵母為主。泡菜腐敗時5 株真菌的數量有所增加,數量是發酵后(第12天)的1~3 倍。泡菜腐敗時,40 g/L鹽質量濃度泡菜鹽鹵以庫德里阿茲威畢赤酵母、塔克斯假絲酵母和熱帶念珠酵母為主,60 g/L鹽質量濃度泡菜鹽鹵以克魯維畢赤酵母、熱帶念珠酵母以及塔克斯假絲酵母為主,80 g/L鹽質量濃度泡菜鹽鹵以庫德里阿茲威畢赤酵母、白地霉、塔克斯假絲酵母以及克魯維畢赤酵母為主。有文獻報道這5 株真菌均為腐敗菌[7,11,25]。
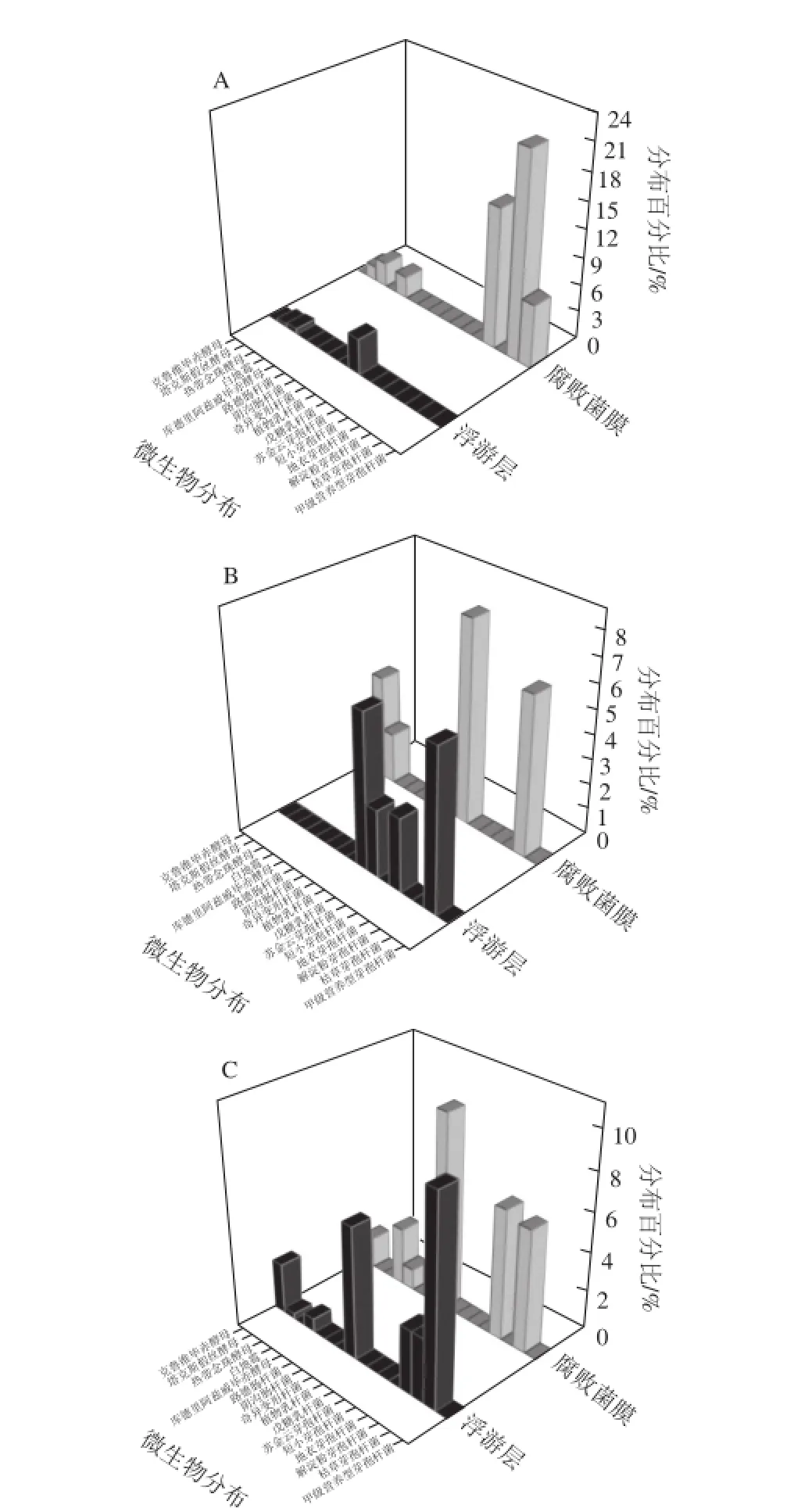
圖7 不同鹽質量濃度泡菜樣品中細菌及真菌的分布Fig.7 Bacterial and fungal distribution in pickle brines with different salt concentrations
一般認為,腐敗泡菜表面形成的膜醭(俗稱生花)主要由真菌組成[3,7],但本實驗中,泡菜腐敗時鹽鹵所形成生物膜是由真菌和細菌共同組成(圖7),且鹽質量濃度越低,膜醭中細菌所占微生物總數的比例越高(圖7A)。
3 結 論
在不同鹽質量濃度四川泡菜的正常發酵過程中,以植物乳桿菌和戊糖乳桿菌為主要優勢菌群。當泡菜發生腐敗后,不同鹽質量濃度泡菜中的腐敗微生物都以芽孢桿菌和酵母為主,但不同鹽質量濃度腐敗泡菜鹽鹵中芽孢桿菌和酵母種類有所差異;腐敗泡菜鹽鹵表層的膜醭是由細菌和真菌共同形成的。該研究結果為四川泡菜的腐敗防控提供了理論參考。
[1] 田偉, 張琦, 鄧珍珍, 等. 利用16S rRNA分析傳統四川發酵泡菜中的細菌多樣性[J]. 食品科學, 2013, 34(17): 215-218. doi: 10.7506/ spkx1002-6630-201317046.
[2] 林燕文, 黃君紅, 黃建杏, 等. 泡菜營養強化食品的研制初探[J]. 食品科技, 2001, 26(2): 28-29.
[3] 羅冬英, 尹傳武. 乳酸菌制劑對人體保健功效的機理探討[J]. 鄂州大學學報, 2002(4): 53-54.
[4] CHOI S M, JEON Y S, RHEE S H, et al. Red pepper powder and kimichi reduce body weight and blood and tissue lipids in rats fed a high fat diet[J]. Nutraceuticals and Food, 2002, 7(2): 162-167.
[5] 蔣和體, 盧新軍. 泡菜對大鼠血脂的調節作用研究[J]. 食品科學,2008, 29(1): 314-316.
[6] 鐘少樞, 吳克剛, 柴向華, 等. 七種單離食用香料對食品腐敗菌抑菌活性的研究[J]. 食品工業科技, 2009, 30(5): 68-71.
[7] 饒瑜, 常偉, 龔麗, 等. 四川泡菜生花酵母的分離與鑒定[J]. 食品與發酵科技, 2013, 49(3): 19-22.
[8] 余文華, 張蜀艷, 陳功, 等. 直投式功能菌對青菜鹽漬發酵過程中各指標變化研究[J]. 食品與發酵科技, 2011, 47(6): 22-25.
[9] 楊瑞, 張偉, 徐小會. 泡菜發酵過程中主要化學成分變化規律的研究[J]. 食品工業科技, 2005, 26(2): 95-98.
[10] SUZANNE D J, FRANCO W, PEREZ-DIAZ I M, et al. Influence of Sodium chloride, pH, and lactic acid bacteria on anaerobic lactic acid utilization during fermented cucumber spoilage[J]. Food Microbiology,2012, 77(7): 394-404.
[11] 敖曉琳, 蔡義民, 夏姣, 等. 引起泡菜“生花”腐敗微生物的分離鑒定[J]. 食品科學, 2013, 34(21): 95-98. doi: 10.7506/spkx1002-6630-201321042.
[12] 鄯晉曉. 四川泡菜菌系分離、篩選及發酵劑的研究[D]. 重慶: 西南大學, 2008: 5-9.
[13] 何玲, 李勤振. 漿水芹菜發酵過程中優勢菌群的分離、鑒定及變化[J].食品科技, 2010, 35(5): 36-40.
[14] FRANCO W, PEREZ-DIAZ I M, JOHANNINGSMEIER S, et al. Characteristics of spoilage-associated secondary cucumber fermentation[J]. Applied and Environmental Microbiology, 2013,78(4): 1273-1284.
[15] XIONG Tao, GUAN Qiangqian, SONG Suhua, et al. Dynamic changes of lactic acid bacteria flora during Chinese sauerkraut fermentation[J]. Food Control, 2012, 26(1): 178-181.
[16] PLENGVIDHYA V, BREIDT F, LU Zhongjing, et al. DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations[J]. Applied and Environmental Microbiology, 2007, 73(23): 7697-7702.
[17] YU Jie, GAO Wa, SUN Zhihong, et al. Identification and characterization of lactic acid bacteria isolated from traditional pickles in Sichuan, China[J]. Journal of General and Applied Microbiology,2012, 58(3): 163-172.
[18] FRANCO W, PEREZ-DIAZ I M. Role of selected oxidative yeasts and bacteria in cucumber secondary fermentation associated with spoilage of the fermented fruit[J]. Food Microbiology, 2012, 32(2): 338-344.
[19] FRANCO W, PEREZ-DIAZ I M. Development of a model system for the study of spoilage associated secondary cucumber fermentation during long-term storage[J]. Journal of Food Science, 2012, 77(10):586-592.
[20] 李由. 提高郫縣豆瓣氨基碳氮含量關鍵技術的研究[D]. 成都: 西華大學, 2010: 3-4.
[21] DEVEREUX R, HE S H, DOYLE C L, et al. Diversity and origin of Desulfovibrio species: phylogenetic definition of a family[J]. Journal of Bacteriology, 1990, 172(7): 3609-3619.
[22] YAN Pingmei, XUE Wentong, TAN S S, et al. Effect of inoculating lactic acid bacteria starter cultures on the nitrite concentration of fermenting Chinese paocai[J]. Food Control, 2008, 19(1): 50-55.
[23] OH C K, OH M C, KIM S H. The depletion of sodium nitrite by lactic acid bacteria isolated from kimchi[J]. Journal of Medicinal Food,2004, 7(1): 38-44.
[24] OH M C, OH C K, KIM S H. Depletion of nitrite by lactic acid bacteria isolated from commercial kimchi[J]. Journal of the Korean Society of Food Science and Nutrition, 2009, 38(7): 892-901.
[25] MOON S H, CHANG Mi, KIM H Y. Pichia kudriavzevii is the major yeast involved in film-formation, off-odor production, and texturesoftening in over-ripened kimchi[J]. Food Science and Biotechnology,2014, 23(2): 489-497.
Comparative Analysis of Microflora Profile in Spoilage Sichuan Pickles with Different Salt Concentrations before and after Spoilage
WANG Meng1, JIANG Yunlu1, YANG Jiantao1, CHANG Wei2, CHE Zhenming1, CHEN Gong3, RAO Yu1,*
(1. College of Bioengineering, Xihua University, Chengdu 610039, China; 2. Institute of Salmon, Tongwei Group Co. Ltd.,Dujiangyan 611800, China; 3. Sichuan Academy of Food and Fermentation Industries, Wenjiang 611130, China)
The microbial population, diversity and dynamics of Sichuan pickles with different salt concentrations were assessed. Using different selective media, 168 microbes were isolated from pickle brines with three different salt concentrations and identified by 16S rDNA or 18S rDNA analysis. The results showed the lactic acid bacteria Lactobacillus plantarum and Lactobacillus pentosus dominated the normal process of Sichuan pickle fermentation. When Sichuan pickle was deteriorated, Bacillus and yeast especially Bacillus subtilis and Candida tanzawaensis were the dominant flora in the pickle samples. Furthermore, Bacillus methylotrophicus, Bacillus licheniformis, Pichia kudriavzevii and Candida tropicalis were isolated from pickle brine with 40 g/L salt. Bacillus pumilus, Pichia kluyveri var. cephalocereana and Candida tropicalis were the most abundant isolates from pickle brine with 60 g/L salt. The major representatives of spoilage-causing microbes involved in pickle brine with 80 g/L salt were Proteus mirabilis, Bacillus amyloliquefaciens, Galactomyces geotrichum, Bacillus licheniformis, Pichia kudriavzevii and Pichia kluyveri var. cephalocereana. The biofilms formed on the surface of spoiled pickle brines consisted of bacteria and fungi. When the salt concentration was higher in the pickle brine,the more proportion of bacteria existed in the biofilm.
Sichuan pickles; salt concentration; normal fermentation; spoilage; dominant flora
TS201.3
A
1002-6630(2015)13-0184-06
10.7506/spkx1002-6630-201513034
2014-09-15
四川省教育廳科研重點項目(13ZA0026);西華大學開放基金項目(szjj2013-047);西華大學“西華杯”大學生科技創新項目(2014134);西華大學研究生創新基金項目(ycjj2014105)
王猛(1988—),男,碩士,研究方向為食品工程。E-mail:164651862@qq.com
饒瑜(1982—),女,副教授,博士,研究方向為食品微生物。E-mail:ryfish@163.com
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
