長白豬TAS2R1基因克隆及序列分析
2015-10-31 08:32:18魏成曉
豬業科學 2015年12期
魏成曉
(青島農業大學,山東 青島 266109)
長白豬TAS2R1基因克隆及序列分析
魏成曉
(青島農業大學,山東 青島 266109)
本研究通過引物合成、PCR擴增、膠回收和測序對長白豬苦味受體基因1(TAS2R1基因)進行序列分析。將測序得到的結果同GenBank中的TAS2R1基因序列進行比對,共發現3處突變位點。分別位于該基因的228 bp、583 bp和712 bp處。其中,在228 bp處由T突變成C,屬于同義突變,其編碼的氨基酸無變化;在583 bp處由C突變成G,屬于錯義突變,編碼的氨基酸由亮氨酸突變為纈氨酸;在712 bp處由A突變成G,屬于錯義突變,編碼的氨基酸由亮氨酸突變為纈氨酸。通過生物學信息軟件DNAMAN分析,發現其與GenBank中的TAS2R1基因序列一致性達99.67%。
基因克隆;長白豬;TAS2R1;序列分析
生物對苦味物質的感知主要是通過苦味受體轉導的,由許多苦味受體基因家族編碼所決定該受體的功能。由于苦味受體基因所表達的苦味物質在自然界中的豐富性,以及膜蛋白在此種實驗中在細胞膜上難以表達[1],因此必須利用分子技術進行苦味受體基因的研究。現在新興的分子層面的技術包括比較基因組學和分子進化理論以及進化基因學手段,可以使我們了解苦味基因家族的演化歷史和演化驅動力,從而使我們在苦味基因的研究上取得一定的進展。科學家對脫氧核糖核酸數據庫進行分析,通過比對遺傳草圖上脊椎動物對苦味敏感的基因座位的具體方位,發現了動物體內的2種相似的苦味受體基因[2],將其定名為TRB與TAS2R。通過對人和小鼠的苦味受體基因探究,又重新發現了人具有10個TAS2R基因,而小鼠則具有30個TAS2R基因。至此,苦味基因在脊椎動物中的所有成員全部被找到。
本試驗成功克隆并分析了長白豬的TAS2R1基因序列,具體揭示了長白豬的這一段基因組成及發生突變的具體位置,為進一步研究TAS2R1基因在其生長發育過程中的作用及分子進化奠定了基礎。
1 試驗材料與方法
1.1試驗材料和主要儀器設備
長白豬耳樣取自青島農業大學試驗基地,放入無水乙醇中保存帶回,于-20 ℃貯存。
所需10×PCR Buffer、dNTP、rTaq DNA聚 合 酶;PCR擴 增 儀(BIO RAD,美國);凝膠成像系統(Alpha Innotech,美國);低溫高速離心機(Heraeus,德國);-80 ℃超低溫冰箱(海爾,中國);移液器(Eppendorf,德國);超純水系統(Milli-Q);高壓滅菌鍋(Hirayama,日本)等。
1.1.1主要試劑的配制
1)50×TAE 的配制。先精確稱取C4H11NO3 242 g,EDTA-2Na 37.2 g放在燒杯內,其次添加900 mL的去離子水,經過充分攪拌后使其溶解。最后添加57.2 mL的CH3COOH,使其完全混勻,最后用去離子水定容達到1 L,放置在室溫下儲存。
2)1×TAE 的配制。精確量取20 mL 50×TAE 濃儲液,向其中添入蒸餾水定容,使之達到1 000 m L,上下顛倒充分混勻待用。
3)1MTris·Cl(pH8.0)。把24.22 gTris放置于80 mL雙蒸餾水中使其溶解,向其中加入濃HCl,將其pH調節到8.0,定容到200 mL,進行高壓滅菌后,于4 ℃保存。
4)10%SDS。在60 ℃的條件下,將20 gSDS加入超純水中溶解,通過滴加稀HCl,將其pH調節到7.2,定容到200 mL,放置于室溫保存。
5)0.5MEDTA(pH8.0)。稱出93 gNa2EDTA·2H2O在160 mL超純水中溶解,將其混合均勻后加入2 g左右NaOH,將其pH調節到8.0,使其定容到200 mL,于4 ℃高壓滅菌后保存。
6)蛋白酶K(20 mg/mL):將200 mg蛋白酶在10 mL超純水中溶解,混合均勻后,定容至500 μL分裝,放置在-20 ℃冷凍儲藏。
1.2耳樣的采取
將耳號鉗用75%的酒精棉消毒,剪下剛出生仔豬的耳樣1~2塊,放入1.5 mL盛有75%酒精的離心管內,帶回實驗室,于-20 ℃保存。
1.3DNA的提取
取豬耳樣一小塊,洗凈后,用剪刀剪碎放在于1.5 mL的滅菌離心管內,按照以下步驟進行操作:
1)加入600 μL組織提取液(10mM EDTA(pH=8.0)+10 mM Tris-Hcl(pH=8.0)),其 次 向 其 中 加 入10%SDS10 mL/mL蛋白酶K使最終濃度達到1%、100 μg/mL,55%消化過夜;
2)于第2天將溶液冷卻使其達到室溫后,導入入體積相等的三羥甲基氨基甲烷飽和酚,將管蓋蓋緊,使離心管緩慢顛倒,顛倒10 min以上即可,12 000 r/min下離心15 min;
3)將體積相等的飽和酚加入上層清液中:氯仿(1:1),將管蓋蓋緊,將離心管緩慢顛倒,顛倒最少10 min以上,12 000 r/min下離心15 min;
4)取上層清液,向其中加入體積相等的三氯甲烷:異戊醇(24:1),將管蓋蓋緊,使離心管緩慢顛倒,最少持續10 min以上,12 000 r/min下離心15 min;
5)取上層清液,加入1/10體積的2 mol/L CH2CH3NaO2和2倍體積的C2H5OH,管蓋蓋緊后,使離心管緩慢顛倒,出現白色絮狀DNA為止,12 000 r/min下離心4~6 min;
6)將上層清液倒掉,并加入400 μL70%無水乙醇經過2次洗滌后,管蓋蓋緊后,將離心管緩慢顛倒,12 000 r/min下離心4~6 min,棄掉管內乙醇,把離心管顛倒在吸水紙上,經干燥處理后,使其溶解,放置于-20 ℃下用來保存以及以后使用。
1.4引物的設計與合成
依據目的基因TAS2R1設計一對引物,通過上海生工合成:
上游引物:5′AGCAAATGTCT GCCATCTTCC 3′,
下游引物:5′GGACATCTTTG CTGGGCTGG 3′。
PCR的 反 應 體 系:10xBuffer 2.5 μL,18.3 μL滅菌蒸餾水,dNTPMix1 μL,上游引物1 μL,下游引物1 μL,TapDNApolymerase(5U/ μL)0.2 μL,DNA模板1 μL。
PCR的反應條件:95 ℃,將其預變性3 min;94 ℃,變性30 s;58 ℃,退火40 s;72 ℃,延伸50 s;返回到變性過程并運用30個循環;72 ℃,延伸10 min;然后于4 ℃保存。PCR反應完成時選取5 μL產物,采用1.5%瓊脂糖凝膠電泳進行檢驗。
1.5基因片段的回收、純化、和測序
PCR的產物經由1%瓊脂糖凝膠電泳后,對切下的目的基因(1189 bp)片段,選取膠回收純化試劑盒進行回收。具體的方法如下:
1)將每100 mg凝膠加入約400 μL的BindingBuffer,放置于50 ℃水浴10 min,每2 min顛倒混勻1次。
2)用2.0 mL收集管中的吸附柱收集融化的膠溶液,在室溫下靜置2 min后,室溫下8 000 r/min離心2 min。
3)將吸附柱拿下,收集管中的廢液全部倒掉,把UNIQ-10柱插入收集管內,將600 μLWash Solution加入其中,10 000 r/min室溫離心40 s。重新洗滌1次。
4)將吸附柱拿下,收集管中的廢液全部倒掉,把UNIQ-10柱插入收集管內,室溫下10 000r/min離心15 s。
5)將吸附柱放進新管中,將20μL Elution Buffer,55 ℃水浴2 min。25 ℃ 10 000 r/min離心3 min,將DNA片段進行回收,就是離心管內的液體。
2 試驗結果
2.1PCR產物瓊脂糖凝膠電泳結果
通過1%瓊脂糖凝膠電泳顯示在1 189 bp處獲得一條清晰的條帶(圖1),與目的條帶大小基本一致。
2.2序列測定結果
測序結果顯示和GenBank里的TAS2R1基因序列一致性達99.67%。DNAMAN序列比對結果如下:
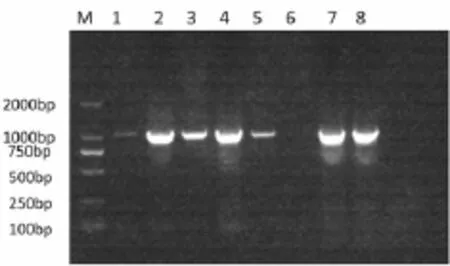
圖1 TAS2R1基因PCR產物瓊脂糖凝膠電泳結果圖
2.3SNPs分析
通過查看TAS2R1基因擴增產物序列測定結果,共顯示3個雙峰位置。將所檢測出的SNPs與GenBank中的TAS2R1進行比對,發現3個突變位。上述結果表明,第228 bp處發生了突變,由T變成了C,該突變為同義突變;第583bp處發生了突變,由C變成了G,該突變為錯義突變,編碼的亮氨酸變成了纈氨酸;第712 bp處發生了突變,由A變成了G,該突變為錯義突變,編碼的氨基酸由異亮氨酸變成了纈氨酸。通過DNAman比對發現一致性達99.67%。將所檢測出的SNPs與GenBank比對,發現3個突變位點中有3個在外顯子區域,其中導致錯義突變的有2個。
3 結論與討論
本研究對長白豬TAS2R1基因的研究結果表明,長白豬該基因的結構與人、牛、馬、羊和小鼠等其他哺乳動物種類的一樣,基因內部無內含子,即整個閱讀框架只有1個外顯子組成,其起始密碼子ATG上游的第3個核苷酸是腺嘌呤,緊跟在ATG后面的核苷酸是鳥嘌呤,這與“Kozak序列”特征一致。符合大多數真核生物起始密碼子所具有的結構特征,說明本研究得到的TAS2R1基因結構是正確的。苦味受體基因家族,具有3個細胞外環和3個細胞內環,并且相互對應。處于TAS2R1細胞外的N端非常短,也許作為配體結合區域,多態性特別明顯,可與多種結構的苦味物質相結合。苦味受體細胞中則具有3個環狀結構,其保守性很高,作為細胞中G蛋白偶聯片段。TAS2R1受體能夠識別苦味物質,主要依賴于基因家族中不同的受體基因能夠與不同的苦味受體物質結合。若是長白豬屢次和具有特殊苦味的物質接觸,也許會增強它們在日后辨別該特種有毒物質的能力。經過對有限的基因序列資料進行整合,可以打破TAS2R1基因序列數目有限的缺陷,使得到的研究結果能夠為我們提供些許TAS2R1基因的演化過程。盡管不同豬物種的食性不同,但是通過研究長白豬的食性選擇,或許能夠揭示食性選擇在基因進化中的動力作用。kim等(2003)對與苯硫脲(phenyl thiocarbamide,PTC)苦味感知相關的苦味受體T2R38的研究結果則表明,PTC味感知者和PTC味非感知者間的差異僅僅是由于T2R38氨基酸序列上3個氨基酸的替換所致。根據Kim等的研究結果,我們可以推測本研究中發現的3處SNPs可能會對苦味物質識別產生一定的影響。通過這個研究,我們或許能夠深入的探討物種形成機制的差異[3]。
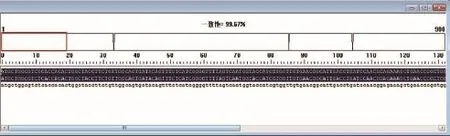
圖2 DNAMAN序列比對結果
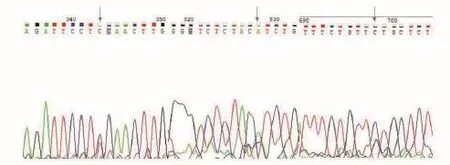
圖3 TAS2R1基因測序結果峰圖
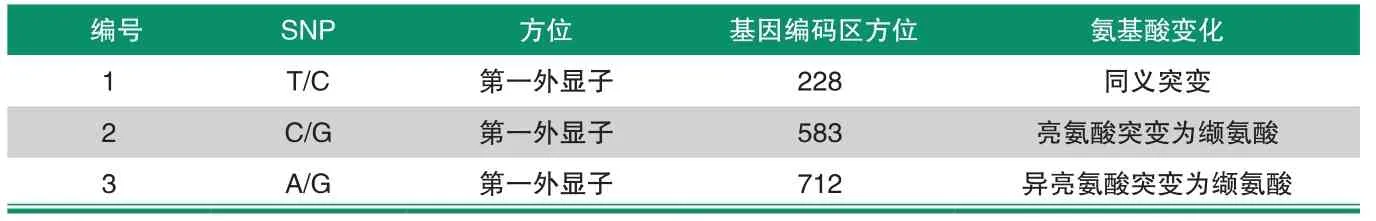
表1 TAS2R1基因的SNP分布
本研究通過對長白豬TAS2R1基因的nucleotide和它的脫氧核苷酸所編碼蛋白的序列進行分析,揭示了其序列特征。對TAS2R1基因的多樣性、基因表達調節和長白豬苦味受體的分子演化順應等方向的研究具有一定意義,同時為長白豬苦味基因遺傳資源的開發和保護應用提供了有力支持。然而,多個基因成員組成了苦味受體基因家族,TAS2R1基因也只是在其中占據了一小部分。為了深入透徹的揭曉長白豬苦味系統的遺傳特征,需要我們進行更深層次的探究。
[1] Nowark R M. Walker's Mammals of the Wodd[M]. Baltimore:Johns Hopkins Univ Press, 1999,9(3):364-417.
[2] Flynn J J,Nedbal MA.Phylogeny of the Carnivora(Mammalia):congruence vs incompatibility among multiple data sets[J]. MolPhylogenet Evol,1998, 9(3):414-426.
[3] Moon SJ,Kttgen M,Jiao Y,et al. A taste receptorrequired for the caffeine response in vivo[J].Curr. Biol,2006(16):1812-817.
2015-10-22)
