高效耐海水型厭氧氨氧化污泥的馴化
2015-11-18 05:16:12李智行陳曉波大連海洋大學海洋科技與環(huán)境學院遼寧大連603大連市市政設計研究院有限責任公司遼寧大連60
中國環(huán)境科學 2015年3期
李智行,張 蕾*,陳曉波,李 航,李 旦(.大連海洋大學海洋科技與環(huán)境學院,遼寧 大連 603;.大連市市政設計研究院有限責任公司,遼寧 大連 60)
高效耐海水型厭氧氨氧化污泥的馴化
李智行1,張 蕾1*,陳曉波2,李 航1,李 旦1(1.大連海洋大學海洋科技與環(huán)境學院,遼寧 大連 116023;2.大連市市政設計研究院有限責任公司,遼寧 大連 116021)
針對部分含海水廢水生物脫氮效能較低的問題,研究了梯度鹽度海水對淡水厭氧氨氧化污泥的馴化過程.考察了不同海水鹽度對厭氧氨氧化反應動力學、厭氧氨氧化菌細胞形態(tài)和反應器中菌群變化的影響.結果表明,梯度鹽度廢水可以成功馴化淡水厭氧氨氧化污泥,通過145d的馴化,其總氮去除速率為2.80kgN/(m3·d).在海水鹽度由0提高至10‰、20‰和30‰的過程中厭氧氨氧化反應速率經(jīng)歷了升高、降低、再升高的過程,其中,海水鹽度20‰在淡水厭氧氨氧化污泥的馴化過程中是一個臨界點.馴化后,厭氧氨氧化菌細胞結構更加不規(guī)則,并在細胞壁上出現(xiàn)了類菌毛狀結構,經(jīng)16S rDNA PCR擴增測序鑒定該優(yōu)勢厭氧氨氧化菌為“Candidatus Kuenenia Stuttgartiensis”.馴化前后反應器中細菌菌群也發(fā)生改變.
厭氧氨氧化;海水;馴化;脫氮
近年來,一些沿海城市為緩解淡水資源日益緊缺的現(xiàn)狀,開始推行海水直接利用或混合利用的方法,將其作為市政用水進行沖廁、街道灑水等,導致城市污水鹽度增加,引發(fā)市政污水處理廠處理難題.此外,許多行業(yè)廢水,如水產(chǎn)養(yǎng)殖廢水、海產(chǎn)品加工廢水,其中也含有大量的海水,鹽度較高,難于處理[1-3].因此,開發(fā)能夠耐受海水的高效菌種尤為必要.這種耐受海水的菌種可來源于2種途徑,一是海洋生物資源的開發(fā),二是淡水污泥的海水馴化.第1種途徑由于對地理等特殊條件的要求往往受限,第2種途徑就顯得尤為重要.
厭氧氨氧化(anammox)工藝是一種近年來研究較多的新型生物脫氮工藝,該工藝依托自養(yǎng)型厭氧氨氧化菌,以氨為電子供體,亞硝酸鹽為電子受體,產(chǎn)生N2.由于能同時去除氨和亞硝酸鹽,無須外加有機碳源,且能夠改善硝化反應產(chǎn)酸、反硝化反應產(chǎn)堿均需中和的情況,其運行費用可比傳統(tǒng)生物脫氮工藝節(jié)省近40%[4].目前發(fā)現(xiàn)的厭氧氨氧化菌包括6個屬,其中淡水環(huán)境中的厭氧氨氧化菌有5個屬,海水環(huán)境中的厭氧氨氧化菌有1個屬[5-6].
眾多學者采用添加NaCl模擬鹽度廢水進行了厭氧氨氧化工藝影響的研究.金仁村等[7]、Ma等[8]、Chen等[9]對淡水厭氧氨氧化污泥進行短期的鹽度沖擊,發(fā)現(xiàn)低濃度的鹽(0,5,10g/L NaCl)可使厭氧氨氧化活性提高,而高濃度的鹽(20,30,50g/L NaCl)則明顯抑制厭氧氨氧化活性.Liu等[10]、Kartal等[11]、Windey等[12]對淡水厭氧氨氧化污泥進行長期的鹽度馴化,發(fā)現(xiàn)通過梯度提高鹽度淡水厭氧氨氧化污泥最終可適應高達30g/L的鹽度環(huán)境,然而當鹽度繼續(xù)升高至30g/L以上,淡水厭氧氨氧化污泥受到可逆抑制.經(jīng)鹽度馴化后微生物群落發(fā)生演變,但優(yōu)勢厭氧氨氧化菌依然是淡水型厭氧氨氧化菌“Candidatus Kuenenia Stuttgartiensis”.然而由于海水本身成分復雜,包括多種有機和無機的、溶解態(tài)和懸浮態(tài)的物質(zhì),其含量約為 30‰ ,其中各組分的含量相差懸殊,難以人工模擬[13].研究表明,不同類型的無機離子對厭氧氨氧化的抑制程度不一[7],而有機物同樣會對厭氧氨氧化產(chǎn)生影響[14-16],但截止到目前,關于有機物對厭氧氨氧化作用機理的研究還比較少,無機物和有機物協(xié)同作用的研究更是未有報道,以往僅采用添加NaCl模擬鹽度條件并據(jù)此展開的鹽度對厭氧氨氧化影響的研究事實上并不完善,不能直接應用于含海水廢水的實際處理.因此本研究針對含海水廢水生物脫氮效率低的問題,期望通過梯度鹽度海水馴化淡水厭氧氨氧化污泥,使其耐受海水鹽度,并具有較好的脫氮效果.在研究過程中考察了不同海水鹽度對厭氧氨氧化反應動力學、厭氧氨氧化菌細胞形態(tài)和反應器內(nèi)菌群變化的影響.
1 材料與方法
1.1 試驗裝置及其運行
有機玻璃制作的上流式厭氧反應器構型及流程如圖1所示.該反應器有效容積9L(內(nèi)徑為95mm、高度為400mm),主體反應區(qū)添加無紡濾布為載體,反應器上方設三相分離系統(tǒng),保證出水、產(chǎn)氣和污泥有效分離.反應器主體外部利用水浴夾層保證反應溫度為37℃,控制水力停留時間(HRT)為0.45d.反應器在本試驗前已采用低鹽模擬廢水進行厭氧氨氧化菌富集,該反應器的總氮去除速率維持在1.35kgN/(m3·d)左右,通過優(yōu)勢菌群鑒定,反應器中的厭氧氨氧化菌為“Candidatus Kuenenia”,為典型的淡水型厭氧氨氧化菌.
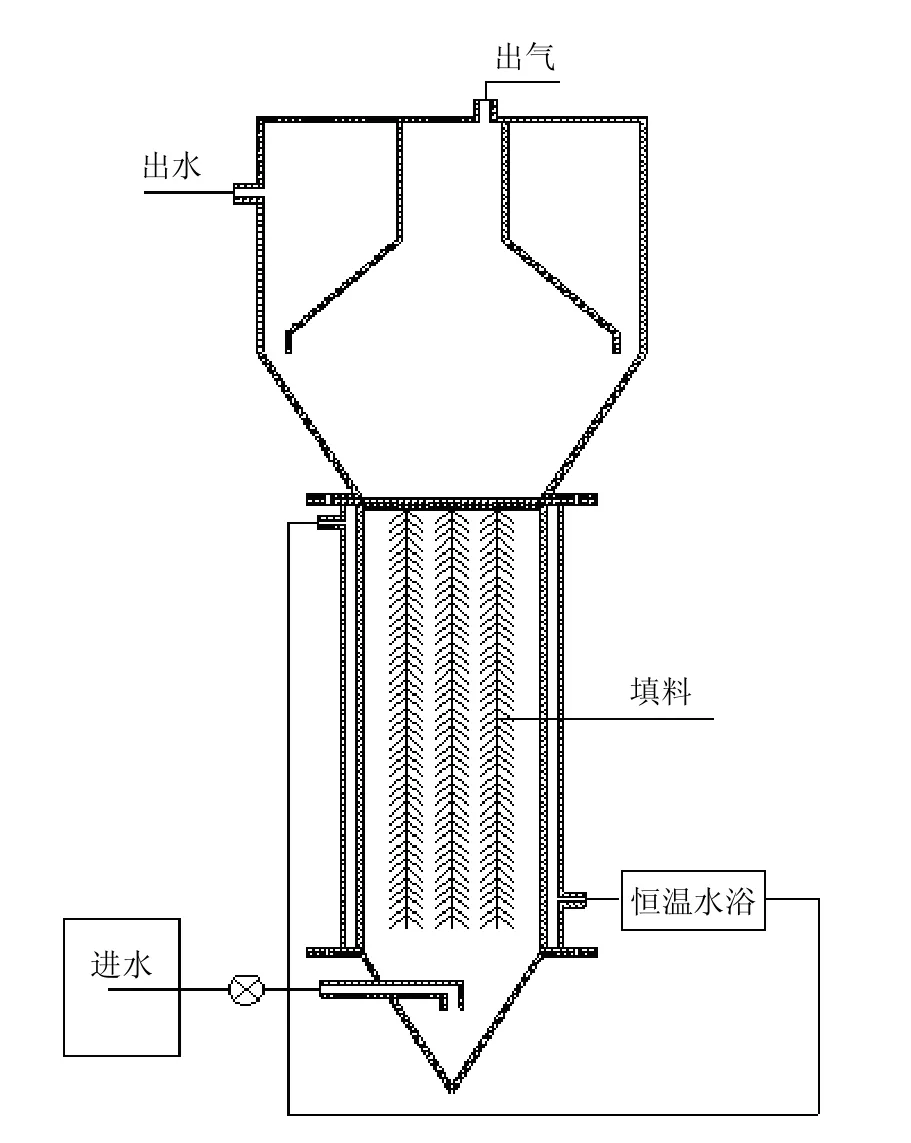
圖1 反應裝置流程示意Fig. 1 Schematic diagram of the reactor
試驗采用梯度鹽度海水對淡水厭氧氨氧化污泥進行馴化,在馴化過程中,保證不同鹽度海水馴化初期進水基質(zhì)濃度相近,且每個鹽度下馴化時間相同,通過逐步提高進水基質(zhì)濃度的方法提高總氮去除速率.同時通過對每個海水鹽度下基質(zhì)代謝動力學參數(shù)的比較考察不同鹽度海水對厭氧氨氧化反應的影響.
1.2 模擬廢水
模擬廢水中的溶劑是比例不同的自來水和鹽度為30‰的砂濾海水混合液,保證混合液中海水鹽度分別為0、10‰、20‰和30‰.基質(zhì)和由NH4Cl和NaNO2按照一定比例提供.此外,該模擬廢水中(g/L)還含有KH2PO40.01,CaCl2·2H2O 0.00565, MgSO4·7H2O 0.3, NaHCO31和微量元素濃縮液I、II各1.25mL[17].微量元素濃縮液I、II組成見表1[18].以NaOH溶液或鹽酸溶液控制進水pH值在7.0~7.5.

表1 微量元素濃縮液的組成Table 1 Composition of trace elements
1.3 厭氧氨氧化污泥比活性測定
1.4 樣品透射電鏡觀察
從反應器中取污泥樣,置于2.5%的戊二醛溶液中,4℃固定過夜,經(jīng)0.1mol/L、pH 7.0磷酸緩沖液漂洗后,再用1%鋨酸溶液固定1~2h,繼續(xù)用磷酸緩沖液漂洗.經(jīng)過50%、70%、80%、90%、95%和100%五種濃度的乙醇溶液脫水處理后,用純丙酮處理20min.接著,分別用體積比為1:1和3:1的包埋劑與丙酮的混合液處理樣品1h和3h,最后用包埋劑處理樣品過夜.將滲透處理的樣品包埋起來,70℃加熱過夜,即得到包埋好的樣品.樣品在Reichert超薄切片機中切片,獲得70~90nm的切片,用檸檬酸鉛溶液和醋酸雙氧鈾50%乙醇飽和溶液各染色15min,再用JEM-2000EX型透射電鏡(日本JEOL公司)觀察結果.
1.5 DNA提取、聚合酶鏈式反應(PCR)、變性梯度凝膠電泳(DGGE)及測序
將反應器在海水鹽度為0和30‰階段后期的污泥樣品進行DNA提取及細菌通用引物擴增.采用上海生能博彩公司環(huán)境樣品DNA提取試劑盒v2.2進行DNA提取,然后以細菌通用引物PRBA338f和PRUN518通過PCR擴增樣品中細菌的DNA片段.反應條件為:94℃預變性10min;94℃變性30s,55℃退火30s,72℃延伸1min,進行30個循環(huán);最后在72℃下延伸7min[17].將PCR產(chǎn)物在35%~65%的變性液梯度條件下進行DGGE,電泳條件為先在120V電壓下預電泳20min,然后在200V電壓下電泳3.5h,在Bio-Rad凝膠成像系統(tǒng)中對DGGE結果進行觀察.對優(yōu)勢菌條帶進行割膠,然后進行測序(寶生物工程有限公司).對測序結果進行BLAST比對后,在MEGA4.0軟件中采用最大簡約法建立系統(tǒng)發(fā)育樹.
同時對海水鹽度30‰階段后期的污泥樣品采用厭氧氨氧化菌特異性引物AMX368f和AMX1480r進行擴增,并對PCR產(chǎn)物直接測序,然后進行BLAST比對.
1.6 測定項目與方法
2 結果與討論
2.1 海水馴化期厭氧氨氧化污泥脫氮性能
圖2為淡水厭氧氨氧化污泥在馴化過程中的脫氮性能.在海水鹽度為0時(1~15d),淡水污泥正處于脫氮性能提高期,在該期最末階段(13~15d)進水和濃度分別為290mg/L和390mg/L,總氮去除速率穩(wěn)定于1.35kgN/(m3·d),總氮去除效率保持在90%左右.
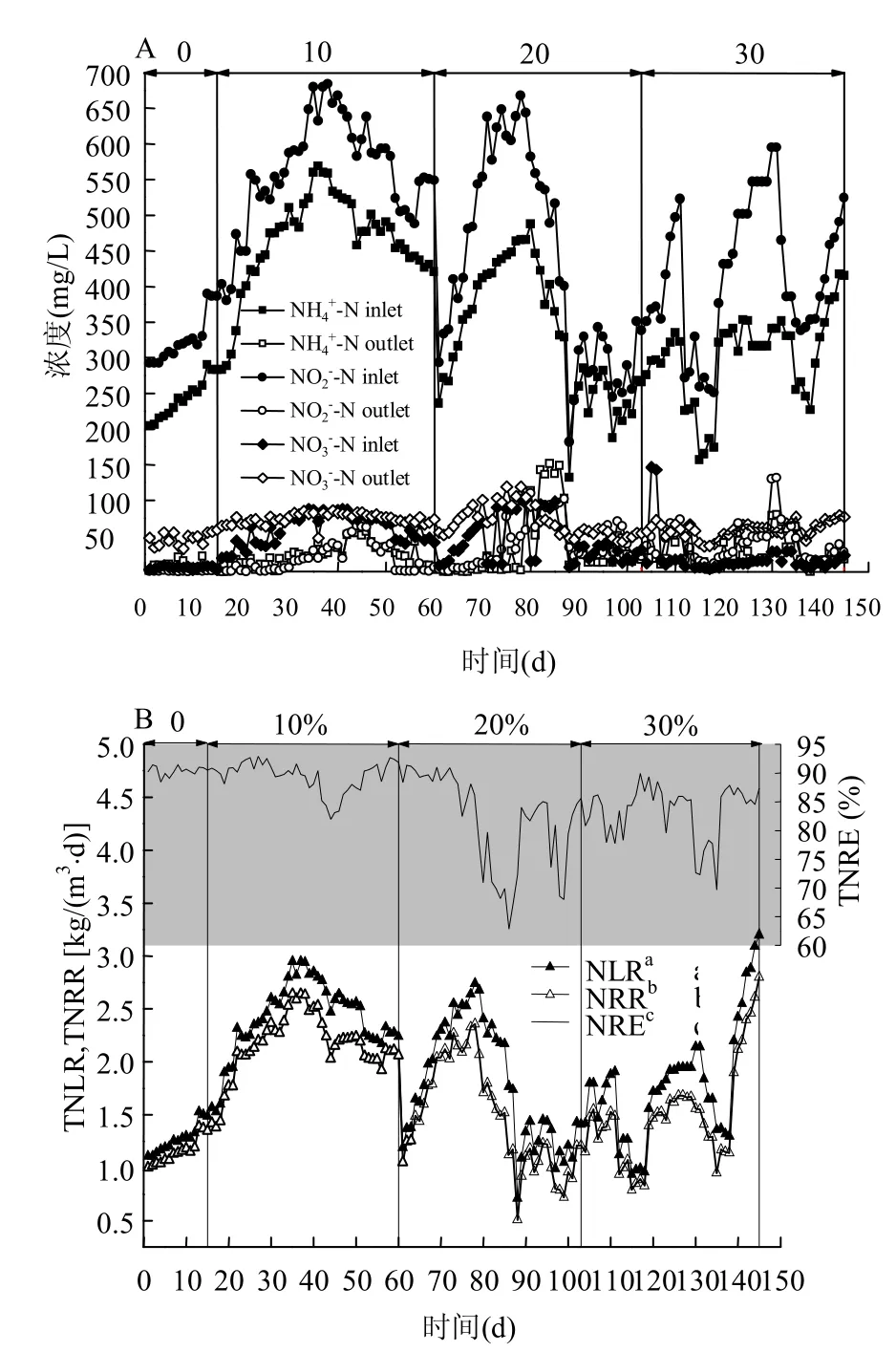
圖2 厭氧氨氧化菌馴化過程中脫氮性能Fig.2 Performance of nitrogen removal during the acclimation of anammox bacteria
繼續(xù)提高海水鹽度至20‰ (61~103d),并保持進水和濃度與海水鹽度10‰的初期濃度一致,總氮去除速率為1.25kgN/(m3·d)左右,比海水鹽度10‰初期略有下降.提高進水和濃度至480mg/L和580mg/L后,出水出現(xiàn)積累抑制現(xiàn)象,總氮去除速率最低降至0.51kgN/(m3·d),總氮去除效率僅為72%.經(jīng)過15d恢復后,總氮去除速率為1.21kgN/(m3·d),去除效率恢復至85%.
由此可見,梯度鹽度海水馴化可使淡水厭氧氨氧化污泥在海水鹽度30‰條件下具有較高的脫氮性能.
2.2 鹽度馴化期厭氧氨氧化反應基質(zhì)代謝動力學
研究表明,亞硝酸為厭氧氨氧化菌的限制性基質(zhì),且對厭氧氨氧化菌會產(chǎn)生基質(zhì)自抑制作用,當濃度為70mg/L時,即可抑制厭氧氨氧化菌的代謝[20].因此本試驗通過控制出水濃度,采用Haldane模型[式(1)]對不同海水鹽度下反應器出水濃度與總氮去除速率建立代謝動力學關系(圖3)[21].通過擬合得到各海水鹽度下的最大總氮去除速率(即厭氧氨氧化反應速率),比較不同海水鹽度下最大總氮去除速率與海水鹽度為0條件下最大總氮去除速率之間的差異率i[式(2)],考察不同海水鹽度對厭氧氨氧化反應基質(zhì)代謝的影響(表2).
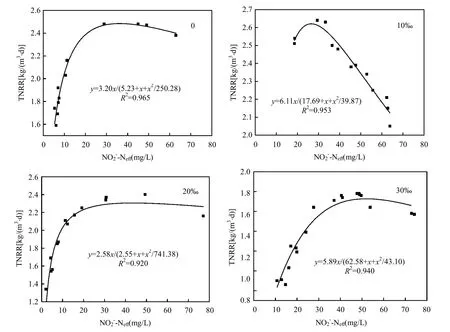
圖3 不同海水鹽度Haldane模型中基質(zhì)代謝動力學Fig.3 Metabolic kinetics of substratewith Haldane model under different seawater contents TNRR:總氮去除速率;:出水亞硝酸氮濃度
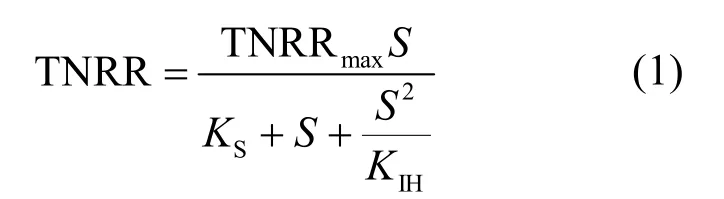
式中: TNRR為總氮去除速率, kg/(m3·d); TNRRmax為最大總氮去除速率, kg/(m3·d); S為出水濃度, mg/L; KS為半飽和常數(shù), mg/L; KIH為基質(zhì)抑制動力學常數(shù), mg/L.
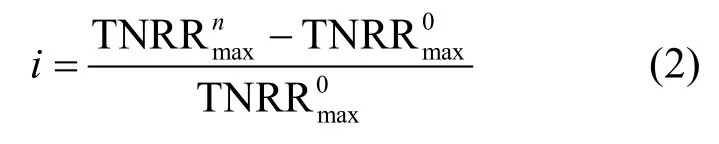
式中: i為不同海水鹽度下最大總氮去除速率與0海水鹽度下最大總氮去除速率之間的差異率;為10‰、20‰、30‰海水鹽度下最大總氮去除速率為0海水鹽度下最大總氮去除速率.

表2 不同海水鹽度下最大總氮去除速率的差異Table 2 Discrepancy of the maximum total nitrogen removal rate under different seawater contents
比較不同海水鹽度下的i值(表2)可以看出海水鹽度僅在海水鹽度20‰階段對厭氧氨氧化反應產(chǎn)生不利影響,其TNRRmax比0海水鹽度階段降低了19.4%;而在海水鹽度10‰和30‰階段,其TNRRmax反而比0海水鹽度階段分別提高了90.9%、84.1%.因此,在本試驗中,厭氧氨氧化反應速率在低鹽度(10‰)下升高,在中鹽度(20‰)下降低,而在高鹽度(30‰)下又再次升高.
以往研究曾發(fā)現(xiàn)厭氧氨氧化反應速率在低鹽度時升高、高鹽度時降低.例如,金仁村等[7]試驗中觀察到了類似的現(xiàn)象,0, 5(5‰), 10 (10‰),20g/L (20‰)不同濃度NaC1的作用下,最大去除速率分別為5.04, 7.30, 6.88,4.24mg/(g VSS·h);最大去除率分別為6.99, 10.55, 9.60, 5.15mg/(g VSS·h).關于低鹽度時厭氧氨氧化反應速率升高的具體原因,有待進一步研究.對于高鹽度時厭氧氨氧化反應速率的降低,一般認為可能是由于在面對外界較高滲透壓的情況下,微生物的代謝受阻所致[22].在此次試驗中,當海水鹽度由20‰升高至30‰后,反應速率提高.產(chǎn)生這一現(xiàn)象的原因可能包括:①經(jīng)過馴化后,污泥的比活性增加;②長時間馴化厭氧氨氧化菌量增長,導致表觀的總氮去除速率增加.本試驗通過血清瓶批次試驗測定了馴化前后厭氧氨氧化污泥的比活性.結果表明,海水鹽度30‰和20‰條件下厭氧氨氧化污泥的比基質(zhì)轉(zhuǎn)化速率分別為0.70和1.02mg N/(g VSS·h).生物量在經(jīng)過30‰海水鹽度馴化后由0.78g VSS/g載體增加至0.82g VSS/g載體.由此可見,海水鹽度由20‰增至30‰階段,污泥比活性降低,而生物量增加,所以由菌量增長引發(fā)的表觀總氮去除速率增加可能是反應器厭氧氨氧化速率提高的主要原因.需要注意的是,活性的降低幅度尚不能由生物量增加所彌補.這是因為血清瓶內(nèi)的微觀環(huán)境并不能完全等價于反應器內(nèi)的宏觀環(huán)境,二者運行模式的不同導致了其內(nèi)部環(huán)境間的差異,包括pH值、溶解氧以及基質(zhì)濃度等均對反應器的脫氮性能至關重要.另外,生物量也與反應器脫氮效能密切相關.研究表明,高菌體濃度可降低單位污泥負荷,增加生長因子,從而可減輕厭氧氨氧化過程中可能發(fā)生的抑制作用,提高反應器運行的穩(wěn)定性[23-25].因此,反應器脫氮效能不能籠統(tǒng)地等價于比基質(zhì)轉(zhuǎn)化速率與生物量的簡單乘法運算.此外,金仁村等[7]發(fā)現(xiàn),在30g/L NaCl作用下,淡水厭氧氨氧化菌活性相對20g/L時降低了37.2%;而Ma等[8]則發(fā)現(xiàn),鹽度沖擊會對淡水厭氧氨氧化菌的生長形成抑制作用,當鹽度沖擊負荷高于10g/L (20g/L和30g/L)時,反應器內(nèi)生物量急劇減少.本試驗中雖然也觀察到了類似的活性降低現(xiàn)象,但其降低幅度僅為31.4%;反應器內(nèi)的生物量不僅沒有減少,在經(jīng)過30‰海水鹽度馴化后反而增長了5.1%. Ma等[8]、Kartal等[11]均在其研究過程中發(fā)現(xiàn)過類似現(xiàn)象.這表明,原有淡水厭氧氨氧化菌本身并不具有對高海水鹽度的固有抗性,但通過梯度提高海水鹽度對淡水污泥進行馴化后,厭氧氨氧化菌的耐海水鹽度性能增強,代謝和繁殖的受抑制程度降低,最終可以適應高達30‰的海水鹽度環(huán)境.海水鹽度20‰很可能是淡水厭氧氨氧化污泥馴化過程中的一個臨界點,這是因為淡水厭氧氨氧化菌為非嗜鹽菌,而該鹽度則是整個馴化過程中第一次超出非嗜鹽菌最佳生長鹽濃度(<11.7‰)的分界點[11,26].
2.3 海水鹽度馴化對厭氧氨氧化菌形態(tài)的影響
采用梯度鹽度海水對淡水厭氧氨氧化污泥進行馴化后,反應器內(nèi)顆粒污泥呈厭氧氨氧化菌所特有的磚紅色[17].采用透射電鏡對原有淡水厭氧氨氧化菌及海水鹽度30‰階段獲得的耐海水型厭氧氨氧化菌的細胞形態(tài)進行觀察.結果表明,二者的形態(tài)發(fā)生顯著變化(圖4).原有淡水厭氧氨氧化菌細胞大小1 μm左右,細胞呈月牙形或鐮刀形,表面有火山口形狀,細胞內(nèi)部大部分被厭氧氨氧化體占據(jù),具有典型的厭氧氨氧化菌結構[16].海水鹽度30‰階段獲得的耐海水型厭氧氨氧化菌細胞大小無明顯變化,細胞形狀更加不規(guī)則,細胞壁外出現(xiàn)類菌毛狀結構,這種結構曾在“Candidatus Scalindua spp.”這種海洋環(huán)境中的厭氧氨氧化菌有過報道[27].
2.4 海水鹽度馴化對菌群變化的影響
利用PCR-DGGE對馴化前后反應器中的細菌菌群變化進行分析,可以看出馴化前后細菌優(yōu)勢菌群發(fā)生明顯變化(圖5).對馴化前后樣品中優(yōu)勢菌條帶(1#、2#、3#、4#、5#、6#)進行割膠測序,后在BLAST數(shù)據(jù)庫進行比對,并建立系統(tǒng)發(fā)育樹(圖6).
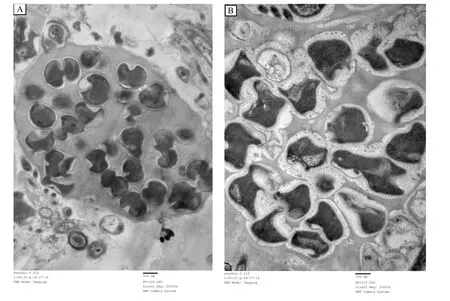
圖4 鹽度馴化前后厭氧氨氧化菌形態(tài)變化Fig.4 Morphology of the non-acclimated and acclimated anammox bacteria

圖5 鹽度馴化前后菌群16S rDNA基因序列變性梯度凝膠電泳圖譜變化Fig.5 Denaturing gradient gel electrophoresis (DGGE)profiles for the 16S rDNA gene fragments of the nonacclimated and acclimated bacterial community
馴化前樣品中的優(yōu)勢菌種主要為1#和2#條帶,其中1#與Candidatus Kuenenia sp. clone(JN182853)的相似度為99%, 2#與Denitratisoma Oestradiolicum clone (KF810120)的相似度為100%.因此在淡水反應器中主要存在厭氧氨氧化菌和反硝化細菌.經(jīng)梯度鹽度海水馴化后的樣品中的優(yōu)勢菌種主要為3#、4#、5#、6#條帶,其中3#與一種鹽堿土壤中獲取的uncultured bacterium clone (JQ427623)的相似度為99%, 4#、5#與自養(yǎng)型脫氮除硫反應器中的uncultured Rhodocyclaceae bacterium clone (GQ324225)的相似度為96%, 6#與鹽沼地中的uncultured bacterium clone (JN684694)的相似度為99%.由此可見,馴化后反應器中細菌主要為不可培養(yǎng)細菌,優(yōu)勢菌與高鹽環(huán)境中的細菌和一些脫氮細菌有較高的相似性.然而由于進行DGGE試驗的PCR產(chǎn)物序列較短(僅236bp),不能夠用于對反應器中細菌的鑒定.采用厭氧氨氧化細菌特異性引物對馴化后的樣品進行16SrDNA PCR擴增、測序(1073bp),對反應器中厭氧氨氧化細菌進行進一步分析.
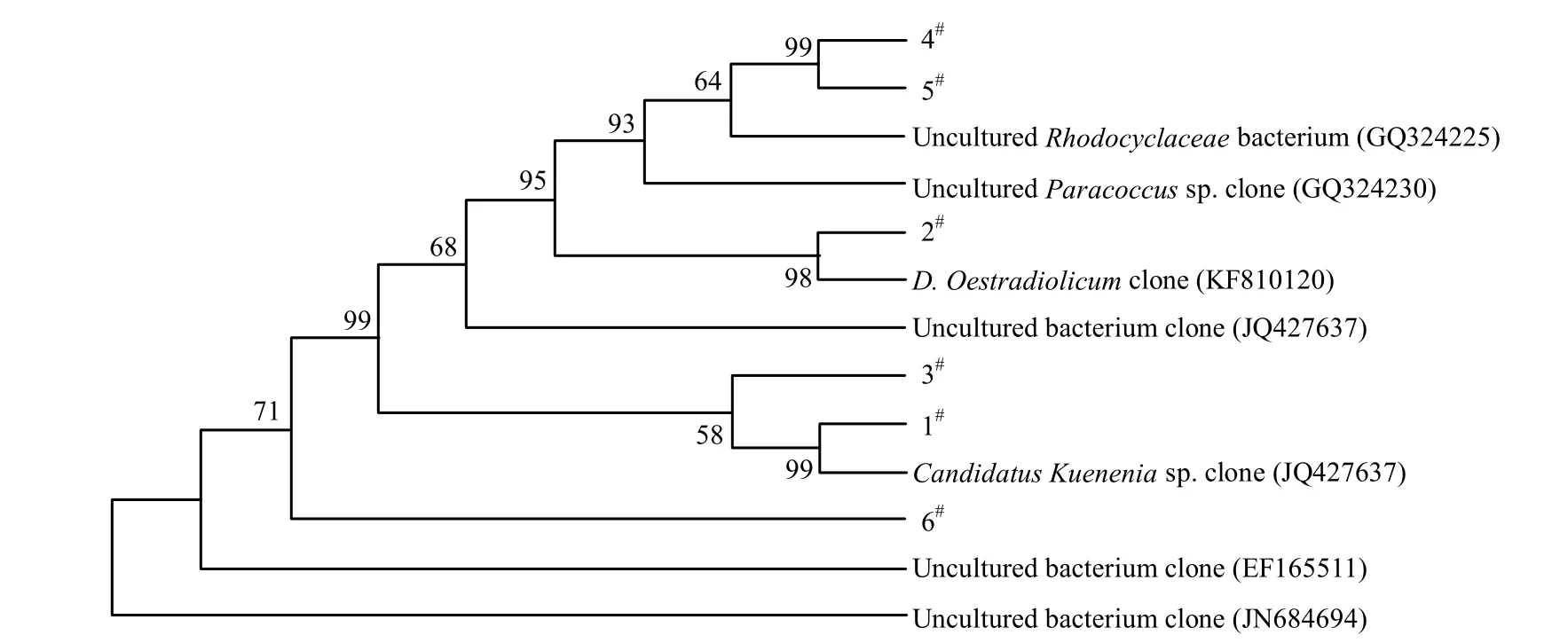
圖6 馴化前后優(yōu)勢菌的系統(tǒng)發(fā)育樹Fig.6 Phylogenetic tree of the dominant bacteria in the non-acclimated and acclimated sludge
通過BLAST比對,結果顯示該序列與厭氧氨氧化菌Candidatus Kuenenia Stuttgartiensis基因組片段(CT573071)的相似度為99%,可認為該菌為Ca. K. Stuttgartiensis. Ca. K. Stuttgartiensis為淡水環(huán)境中的厭氧氨氧化菌,目前未見報道其具有菌毛狀結構,這與形態(tài)觀察結果不一致,這一現(xiàn)象需要進一步研究.
3 結論
3.1 淡水厭氧氨氧化污泥經(jīng)過馴化后,可以在30‰海水鹽度下取得較好的脫氮性能,總氮去除速率可達2.80kgN/(m3·d).
3.2 厭氧氨氧化基質(zhì)代謝動力學表明當海水鹽度由0升高至10‰、20‰及30‰的過程中,厭氧氨氧化反應速率經(jīng)歷了升高、降低、再升高的過程.
3.3 在海水鹽度30‰階段獲得的高效耐海水型厭氧氨氧化菌細胞結構發(fā)生改變,細胞形狀更加不規(guī)則,且細胞壁上出現(xiàn)類菌毛狀結構.經(jīng)16S rDNA PCR擴增測序鑒定該優(yōu)勢厭氧氨氧化菌為Candidatus Kuenenia Stuttgartiensis.
[1]孫曉杰,徐迪民,于德爽.海水沖廁污水的短程硝化試驗研究 [J].中國給水排水, 2007,23(3):40-43.
[2]于德爽,彭永臻,張相忠,等.海水鹽度對短程硝化反硝化的影響[J]. 工業(yè)水處理, 2003,23(1):50-54.
[3]Chowdhury P, Viraraghavan T, Srinivasan A. Biological treatment processes for fish processing wastewater- A review [J]. Bioresour. Technol., 2010,101(2):439-449.
[4]Hu Z Y, Lotti T, van Loosdrecht M, et al. Nitrogen removal with the anaerobic ammonium oxidation process [J]. Biotechnol. Lett.,2013,35(8):1145-1154.
[5]van Niftrik L, Jetten M S M. Anaerobic ammonium-oxidizing bacteria: unique microorganisms with exceptional properties [J]. Microbiol. Mol. Biol. Rev., 2012,76(3):585-596.
[6]Khramenkov S V, Kozlov M N, Kevbrina M V, et al. A novel bacterium carrying out anaerobic ammonium oxidation in a reactor for biological treatment of the filtrate of wastewater fermented sludge [J]. Microbiol., 2013,82(5):628-636.
[7]金仁村,馬 春,鄭 平,等.鹽度對Anammox的短期影響研究[J]. 高校化學工程學報, 2013,27:322-329.
[8]Ma C, Jin R C, Yang G F, et al. Impacts of transient salinity shock loads on Anammox process performance [J]. Bioresour. Technol.,2012,112:124-130.
[9]Chen H, Ma C, Ji Y X, et al. Evaluation of the efficacy and regulation measures of the anammox process under salty conditions [J]. Sep. Purif. Technol., 2014,132:584-592.
[10]Liu C L, Yamamoto T, Nishiyama T, et al. Effect of salt concentration in anammox treatment using non woven biomass carrier [J]. J. Biosci. Bioeng., 2009,107(5):519-523.
[11]Kartal B, Koleva M, Arsov R, et al. Adaptation of a freshwater anammox population to high salinity wastewater [J]. J. Biotechnol., 2006,126(4):546-553.
[12]Windey K, De Bo I, Verstraete W. Oxygen-limited autotrophic nitrification-denitrification (OLAND) in a rotating biological contactor treating high-salinity wastewater [J]. Water Res., 2005, 39(18):4512-4520.
[13]陳 敏.化學海洋學 [M]. 北京:海洋出版社, 2009:13-21.
[14]孫佳晶,張 蕾,張 超,等.有機物作用的厭氧氨氧化菌代謝特性研究進展 [J]. 化工進展, 2012,31(8):1834-1837.
[15]操沈彬,王淑瑩,吳程程,等.有機物對厭氧氨氧化系統(tǒng)的沖擊影響 [J]. 中國環(huán)境科學, 2013,33(12):2164-2169.
[16]李澤兵,劉常敬,趙白航,等.多基質(zhì)時厭氧氨氧化菌、異養(yǎng)反硝化污泥活性及抑制特征 [J]. 中國環(huán)境科學, 2013,33(4):648-654.
[17]張 蕾.厭氧氨氧化性能的研究 [D]. 杭州:浙江大學, 2009.
[18]鄭 平,徐向陽,胡寶蘭.新型生物脫氮理論與技術 [M]. 北京:科學出版社, 2004:90-91.
[19]國家環(huán)境保護總局.水和廢水監(jiān)測分析方法 [M]. 4版.北京:中國環(huán)境科學出版社, 2002:258-281.
[20]Strous M, Kuenen J G, Jetten M S M. Key physiology of anaerobic ammonium oxidation [J]. Appl. Environ. Microbiol.,1999,65(7):3248-3250.
[21]李偉剛,于德爽,李 津.ASBR反應器厭氧氨氧化脫氮II:反應動力學 [J]. 中國環(huán)境科學, 2013,33(12):2191-2200.
[22]Schmid M, Twachtmann U, Klein M, et al. Molecular evidence for genus level diversity of bacteria capable of catalyzing anaerobic ammonium oxidation [J]. Syst. Appl. Microbiol., 2000,23(1):93-106.
[23]Strous M, Fuerst J A, Kramer E H M, et al. Missing lithotroph identified as new planctomycete [J]. Nature, 1999,400(6743):446-449.
[24]Isaka K, Date Y, Sumino T, et al. Growth characteristic of anaerobic ammonium-oxidizing bacteria in an anaerobic biological filtrated reactor [J]. Appl. Microbiol. Biotechnol., 2006, 70(1):47-52.
[25]Tang C J, Zheng P, Chen T T, et al. Enhanced nitrogen removal from pharmaceutical wastewater using SBA-ANAMMOX process [J]. Water Res., 2011,45(1):201-210.
[26]吳月紅.鹽環(huán)境細菌群落結構分析和多相分類學研究 [D]. 杭州:浙江大學, 2009.
[27]van Niftrik L, Geerts W J C, van Donselaar E G, et al. Linking ultrastructure and function in four genera of anaerobic ammonium-oxidizing bacteria: cell plan, glycogen storage, and localization of cytochrome c proteins [J]. J. Bacteriol., 2008,190(2):708-717.
Acclimation of a highly-efficient and seawater tolerant anammox sludge.
LI Zhi-xing1, ZHANG Lei1*, CHEN Xiao-bo2, LI Hang1, LI Dan1(1.Department of Marine Science and Environmental Engineering, Dalian Ocean University,Dalian 116023, China;2.Dalian Municipal Design and Research Institute CO., LTD, Dalian 116021, China). China Environmental Science, 2015,35(3):748~756
The performance of biological nitrogen removal in treating seawater-containing wastewater is not satisfying. Here we report the acclimation of a freshwater-origin anaerobic ammonium oxidation (anammox) sludge by using seawater with increasing salt concentrations. Nitrogen removal performance, morphological characteristics and dominant strains of the bacterial community in the reactor were investigated. After 145days of acclimation, this freshwater-origin anammox sludge was fully adapted to seawater, with a total nitrogen removal rate (TNRR) reaching 2.80kgN/(m3·d). The anaerobic ammonium oxidizing rate first increased, then decreased, and then increased again when the seawater content was gradually increased from 0‰ to 30‰. A seawater content of 20‰ was the critical point during the acclimation. In terms of morphological characteristics, cell shape of the acclimated anammox bacteria was irregular and pilus-like appendages were developed on the cell wall. Change in the bacterial community composition after acclimation was also observed through DGGE.The dominant anammox strain was identified as “Candidatus Kuenenia Stuttgartiensis” based on the sequence of the 16S rDNA gene.
anammox;seawater;acclimation;nitrogen removal
X703.1
A
1000-6923(2015)03-0748-09
李智行(1989-),男,河南新鄉(xiāng)人,大連海洋大學碩士研究生,主要從事廢水生物脫氮研究.
2014-07-31
國家自然科學基金資助項目(41106097);遼寧省高校杰出青年學者成長計劃資助項目(LJQ2012065)
* 責任作者, 副教授, zhanglei@dlou.edu.cn
