石渠縣2001-2013年鼠疫疫源地流行病學分析
2015-11-20 11:42:06汪立茂段勇軍李光軍譚文明
中國人獸共患病學報 2015年5期
祁 騰,楊 孔,汪立茂,段勇軍,謝 飛,楊 軍,李光軍,譚文明
2.西南民族大學生命科學與技術學院;
3.四川省疾病預防控制中心;
4.甘孜州疾病預防控制中心;
5.石渠縣疾病預防控制中心
鼠疫是由鼠疫耶爾森菌引起的人獸共患病,它引起過3次世界鼠疫大流行,給人類文明造成極大危害[1-3]。石渠縣青海田鼠鼠疫自然疫源地發現于1997年,位于四川與青海交界的俄多瑪鄉,平均海拔為4200m,該疫源地鼠疫菌為田鼠型鼠疫菌,與布氏田鼠型鼠疫菌在生物學特征上相似[4-6]。疫源地內主要宿主是青海田鼠,次要宿主是根田鼠、長尾倉鼠等,主要媒介是細鉤蓋蚤、直緣雙蚤指名亞種,次要媒介是五側纖蚤鄰近亞種等。1997-2013年該地動物鼠疫一直流行,研究該地鼠疫監測數據,有助于了解疫源地鼠疫流行的規律。
1 材料與方法
1.1 材料
1.1.1 數據來源 石渠縣2001-2013年鼠疫疫源地監測數據。
1.1.2 檢測試劑 鼠疫血凝試劑由青海省地方病預防控制所提供;鼠疫菌噬菌體由中國疾病預防控制中心鼠疫布氏菌病預防控制基地提供;鼠疫赫氏干燥培養基由北京陸橋公司生產;生理鹽水由四川科倫公司提供,有效期內按說明書使用。
1.2 方法
1.2.1 宿主調查 青海田鼠的調查采用樣方(25m×25m),夜行鼠的調查采用5m夾線發調查,捕獲的樣本單獨裝袋計數并鑒定。
1.2.2 體蚤調查 捕獲的鼠類分別單只裝袋,乙醚麻醉后,在檢蚤盆內用小梳梳檢鼠類體表,裝瓶進行分類鑒定。
1.2.3 青海田鼠穴蚤調查 用工具挖開青海田鼠巢穴后,取巢墊物及表面部分巢土單獨裝袋,扎緊袋口送回,用檢蚤盆單巢檢蚤,裝瓶進行分類鑒定。
1.2.4 血清學檢驗和細菌學培養 按照《鼠疫診斷標準(WS279-2008)》的附錄A和附錄C執行。
1.2.5 數據分析 運用Excel和SPSS11.5進行數據統計分析、圖表制作。
2 結 果
2.1 青海田鼠密度監測 2001-2013年捕獲青海田鼠14 147只,樣方57.807ha,密度244.73只/ha,其中2002年密度最高為775.43只/ha,2012年密度最低為107.64只/ha,添加對數曲線,青海田鼠的鼠密度總體呈下降趨勢,見圖1。
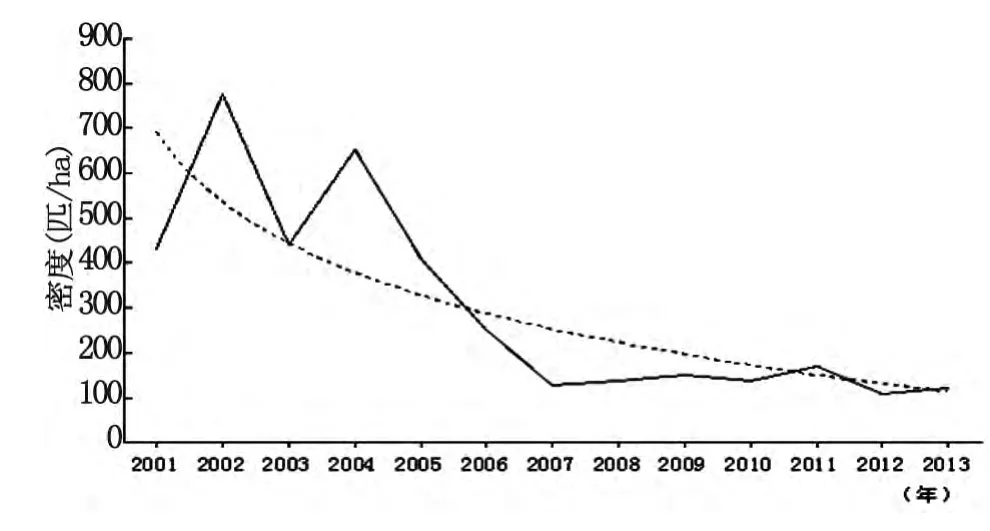
圖1 青海田鼠密度動態變化Fig.1 Dynamic of Microtus fuscus density
2.2 青海田鼠體蚤監測 2001-2013年在14 202只青海田鼠體表獲蚤12 733匹,染蚤青海田鼠6 101只,染蚤率為42.96%,染蚤率最高在2003年,(65.54%),最低在2001年(22.72%);平均蚤指數為0.90,2007年蚤指數最高(1.58),2001年最低(0.39)見表1。
2.3 青海田鼠巢蚤監測 2001-2013年共取青海田鼠巢405個,獲蚤22 512匹,所有鼠巢染蚤,染蚤率為100%;巢蚤蚤指數為55.59,巢蚤蚤指數最高是2001年為79.80,最低為2012年為30.21。在青海田鼠鼠巢中,所獲蚤類經鑒定共3科4屬8種,其中其中角葉蚤科蓋蚤屬(細鉤蓋蚤Csparsilis13 519匹、端園蓋蚤C.kozlovi68匹)、倍蚤屬(嘩倍蚤指明亞種A.c.clarus39匹);細蚤科雙蚤亞科雙蚤屬(直緣雙蚤指明亞種A.t.tuta7 926匹,原雙蚤指明亞種A.p.primaris40匹,青海雙蚤A.qinghaiensis53匹);纖蚤亞科纖蚤屬(五側纖蚤鄰近亞種R.d.vicina852匹、腹竇纖蚤R.li15匹)。
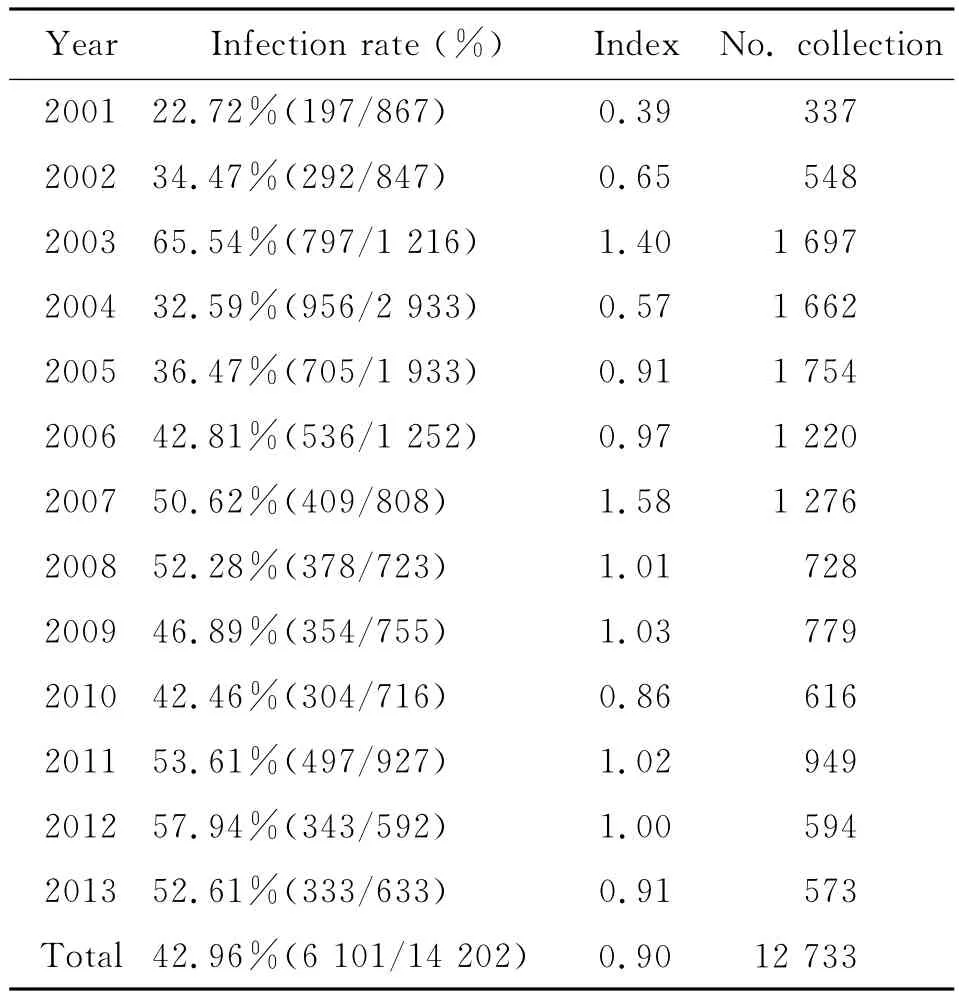
表1 2001-2013年青海田鼠體蚤監測Tab.1 Fleas survey of Microtus fuscus carring from 2001to 2013
2.4 青海田鼠夜行鼠監測 2001-2013年捕獲夜行鼠420只,捕獲率1.56%,其中2001年捕獲率最高為7.30%,2008年最低為0.70%。所捕獲的鼠類經鑒定共3目4科5屬9種和一種未定種的水棲鼠,嚙齒目倉鼠科田鼠亞科田鼠屬(青海田鼠Microtusfuscus231只、根田鼠Microtusoeconomus51只),嚙齒目倉鼠科倉鼠亞科倉鼠屬(長尾倉鼠CricetuluslongicaudatusMilne-Edwards21只、藏倉鼠Cricetuluskamensis2只),嚙齒目林跳鼠科林跳鼠亞科林跳鼠屬(四川林跳鼠Eozapussetchuanus3只);兔形目鼠兔科鼠兔屬(高原鼠兔Ochotonacurzoniae105只、格氏鼠兔Ochotonagloveri1只);食蟲目鼩鼱科鼩鼱屬(暗色鼩鼱Sorexsinalis2只),未定水棲鼠4只,見表2。
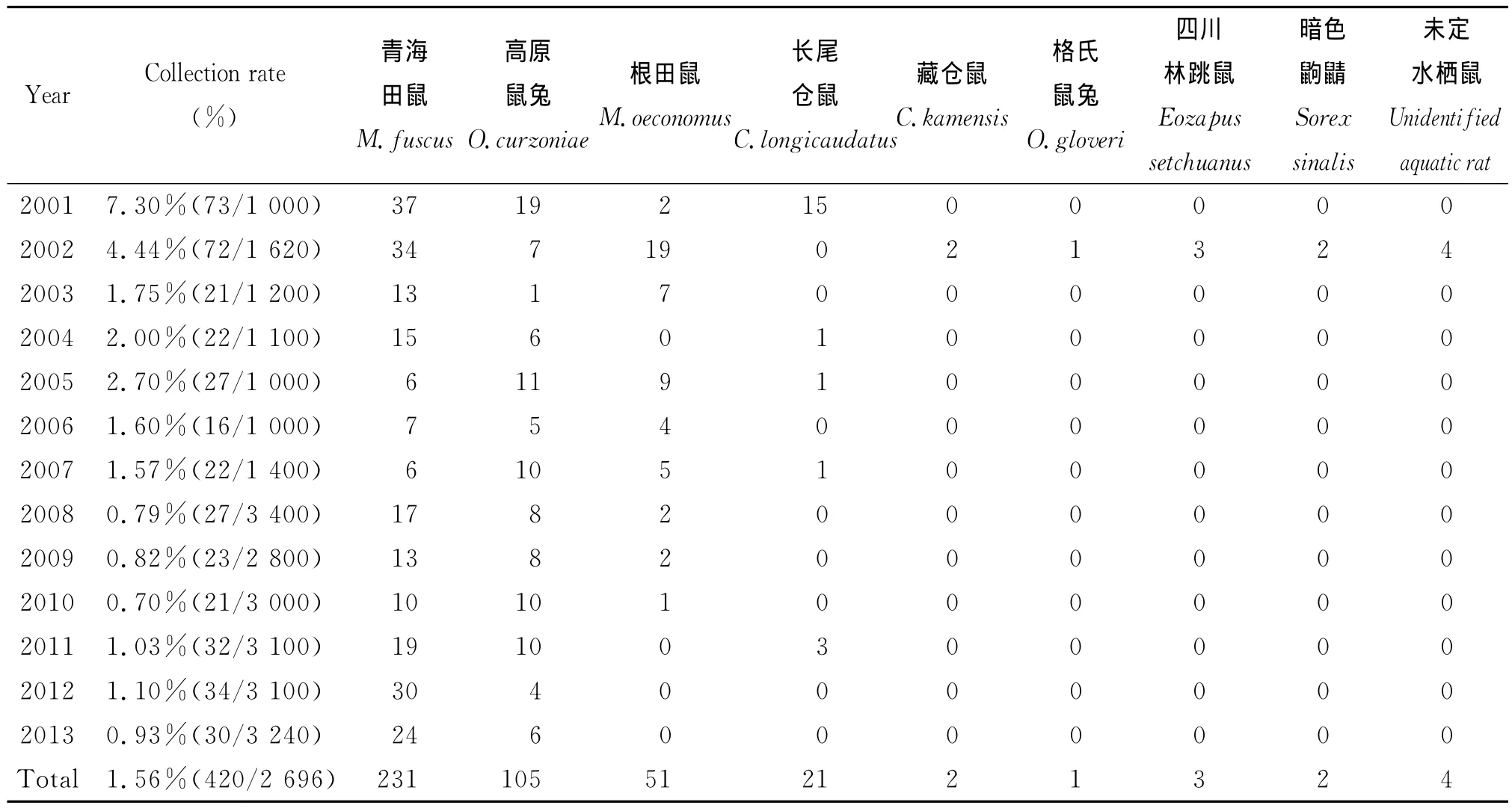
表2 2001-2013年青海田鼠疫源地夜行鼠監測Tab.2 Nocturnal mice survey of Microtus fuscus plague foci from 2001to 2013
2.5 青海田鼠和牧犬血清鼠疫F1抗體檢測2001-2013年血清學檢測青海田鼠血清1 787份,陽性100份,陽性率為5.60%,2014年陽性率最高為23.08%;檢測牧犬血清784份(2008年沒有監測牧犬血清),陽性75份,陽性率9.54%,2001年陽性率最高為20.34%,青海田鼠和牧犬血清F1抗體陽性率動態基本一致,但總體陽性率都在下降,見圖2。
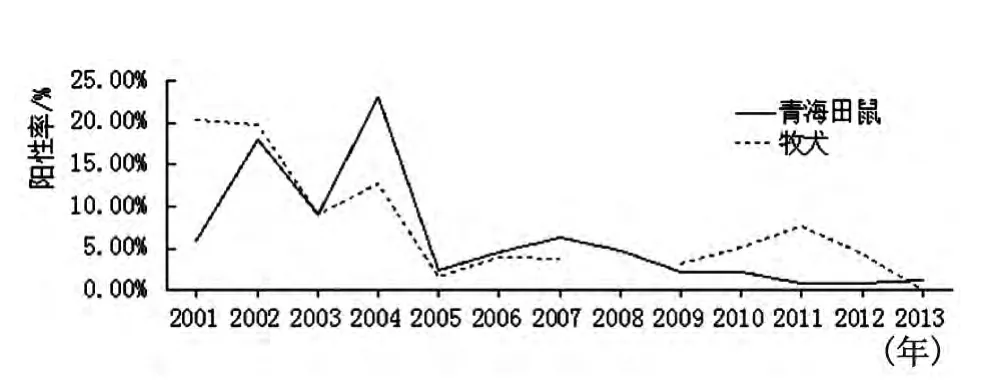
圖2 青海田鼠和牧犬血清F1抗體陽性率變化趨勢Fig.2 Serum F1positive rate trend of Microtus fuscus and shepherd
2.6 青海田鼠細菌學檢測 2001-2013年細菌學檢驗自斃青海田鼠357只,其中陽性52只,鼠疫菌檢出率為14.57%,檢出率最高在2001年63.16%,2003年自斃青海田鼠無鼠疫菌分離。檢驗活體青海田鼠12 955只,陽性29只,鼠疫菌檢出率為0.22%,2001年鼠疫菌檢出率最高為0.66%,2011-2013年活體青海田鼠鼠疫菌檢出數為0,活體青海田鼠的鼠疫菌檢出率在下降,見圖3。
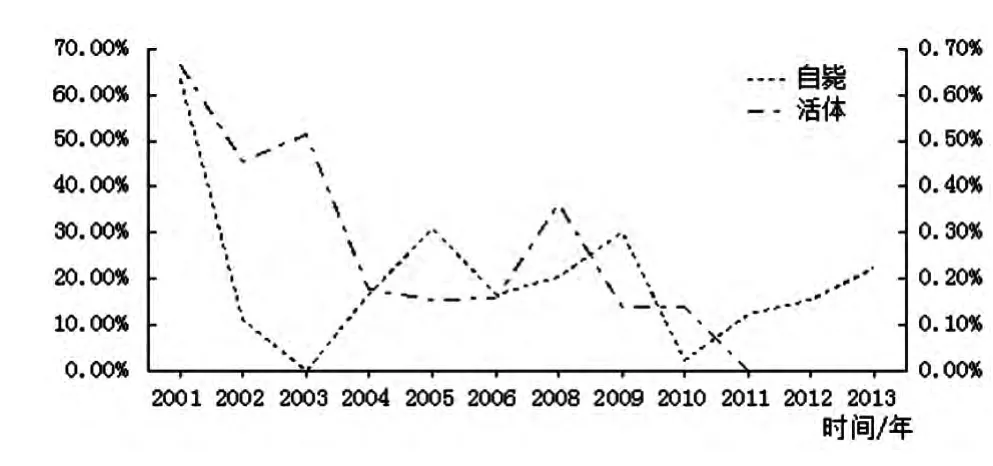
圖3 青海田鼠鼠疫菌檢出率Fig.3 Rate of Y.pestis caught in live and dead Microtus fuscus
3 討 論
鼠疫是一種人獸共患病,同時也是一種自然疫源地性疾病,鼠疫菌是由假結核菌通過獲得、失活、丟失部分基因簇進化而來,而田鼠型鼠疫菌是由古老鼠疫菌的napA硝酸鹽還原酶突變和araC基因失活演變而來。[7-8]
調查發現13年里石渠縣青海田鼠鼠疫自然疫源地的主要宿主青海田鼠的密度呈現下降趨勢,但是密度還是較高,這樣會增加鼠與鼠交叉感染的幾率,有利于鼠疫的傳播,這是造成該疫源地鼠疫流行的主要原因之一。作為鼠疫傳播的重要媒介(蚤)[9],主要寄生在青海田鼠體表和其巢里蚤的數量、種類對鼠疫的流行起著重要的作用,本研究發現青海田鼠蚤指數和染蚤率變化不大,青海田鼠蚤指數圍繞在0.90、染蚤率在42.96%上下波動,染蚤率較高。所有的青海田鼠鼠巢全部染蚤,并且巢蚤蚤指數非常高,最低值都是30.21(2012年)。體蚤和巢蚤有著相同的蚤種分布(3科4屬8種),且主要的寄生蚤是細鉤蓋蚤和直緣雙蚤指明亞種(體蚤中分別 占 65.54%、31.52%,在 巢 蚤 中 分 別 占60.05%、35.21%)二者合計占總蚤數量的90%以上,并且細鉤蓋蚤的比例最大在60%以上,這兩種蚤都分離出鼠疫菌[10-11],巢蚤的數量大,可以很快補充死去的蚤類和體蚤的數量,保障的鼠疫傳播中蚤數量的穩定。其他寄生蚤雖然所占比重少,但是蚤種類的多樣性是鼠疫菌在疫源地內持續存在的原因之一[12],蚤種多樣性越高,以青海田鼠鼠巢為基礎的小的生態系統就越“健康”,對鼠疫菌的保持、傳播起著重要的作用,比如腹竇纖蚤和五側纖蚤鄰近亞種也分離出鼠疫菌[13-14],青海田鼠體蚤和巢蚤的高染蚤率、高蚤指數和蚤的多樣性高是造成疫源地內動物鼠疫流行的又一重要原因。
該疫源地內夜行鼠共有9種,數量排在前3位的是青海田鼠、高原鼠兔和根田鼠,占總數的55.00%、25.00%和12.14%。藏倉鼠、格氏鼠兔、四川林跳鼠、暗色鼩鼱和水棲鼠除2002年捕獲外其他年份并末捕捉到,是因為調查的生境不同造成的,所占比重較少的夜行鼠也對鼠疫菌的儲存和傳播起重要的作用,根田鼠和長尾倉鼠在鼠疫細菌學上已得到證實。該地區的青海田鼠和高原鼠兔呈混合分布,相互交串頻繁,形成了寄生蚤的相互交替,這對于維持疫源性和擴大疫情的蔓延具有重要的流行病學意義[15]。青海田鼠和牧犬血清鼠疫F1抗體的檢出,說明該疫源地非常活躍。說明該疫源動物間鼠疫非常活躍,而且動物鼠疫會持續流行。
[1]Kreppel KS,Caminade C,Telfer S,et al.A non-stationary relationship between global climate phenomena and human plague incidence in Madagascar[J].PLoS Negl Trop Dis,2014,8(10):e3155.DOI:10.1371/journal.pntd.0003155
[2]Green MH,Jones L,Little LK,et al.Yersiniapestisand the three plague pandemics[J].Lancet Infect Dis,2014,14(10):918.DOI:10.1016/S1473-3099(14)70877-1
[3]Harbeck M,Seifert L,Hansch S,et al.YersiniapestisDNA from skeletal remains from the 6th century AD reveals insights into Justinianic plague[J].PLoS Pathog,2013,9(5):e1003349.DOI:10.1371/journal.ppat.1003349
[4]Qi ZZ,Zhao HH,Li CX,et al.Biological features comparison ofYersiniapestisisolated from Microtus Fuscus with that from Microtus Brandti in China[J].Chin J Endemiol,2005,24(5):485-487.(in Chinese)祁芝珍,趙海紅,李存香,等.我國青海田鼠與布氏田鼠鼠疫菌生物學特征比較[J].中國地方病學雜志,2005,24(5):485-487.
[5]Hai R,Wei JC,Cai H,et al.Molecular biology characters ofYersiniapestisstrains isolated from Shiqu county,Sichuan province,China[J].Chin J Vector Bio Ctrl,2002,13(1):19-23.(in Chinese)海榮,魏建春,蔡虹,等.四川石渠縣鼠疫耶爾森菌的分子生物學特征[J].中國媒介生物學及控制雜志,2002,13(1):19-23.
[6]Liu ZC,Hai R,Li FZ,et al.The discovery and study ofMicrotusFuscusnature plague foci in Qinghai-Tibet plateau[J].Chin J Ctrl Endem Dis,2001,16(6):321-327.(in Chinese)劉振才,海榮,李富忠,等.青藏高原青海田鼠鼠疫自然疫源地的發現與研究[J].中國地方病防治雜志,2001,16(6):321-327.
[7]Holt AC,Salkeld DJ,Fritz CL,et al.Spatial analysis of plague in California:niche modeling predictions of the current distribution and potential response to climate change[J].Int J Health Geogr,2009,8(38):1-14.DOI:10.1186/1476-072X-8-38
[8]Fang XY,Yang RF,Liu QY,et al.A novel method for typing natural plague foci in ChinaⅡ.Research on the typing methods for natural plague foci[J].Chin J Endemiol,2012,33(2):234-238.(in Chinese)方喜業,楊瑞馥,劉起勇,等.中國鼠疫自然疫源地分型研究Ⅱ鼠疫自然疫源地分型方法研究[J].中國地方病防治雜志,2012,33(2):234-238.
[9]Christopher FB,Austin KV,Clayton OJ,et al.Evaluation of the murine immune response toXenopsyllacheopisflea saliva and its effect on transmission ofYersiniapestis[J].PLoS One,2014,8(9):e3196.DOI:10.1371/journal.pntd0003196
[10]Yue SL,Yu XT,Li M,et al.The first time ofYersiniapestisisolated fromA.t.tuta[J].Endem Dis Bull,2002,17(3):89.(in Chinese)岳珊瓏,于曉濤,李民,等.首次自直緣雙蚤指名亞種體內檢出鼠疫菌[J].地方病通報,2002,17(3):89.
[11]Yue SL,Li M,Wang YZ,et al.The first time ofYersiniapestisisolated fromCsparsilis[J].Chin J Endemiol,2008,27(6):647-650.(in Chinese)岳珊瓏,李民,汪元忠,等.首次自細鉤黃蚤體檢出鼠疫菌[J].中國地方病學雜志,2008,27(6):647-650.
[12]Eisen RJ,Borchert JN,Mpanga JT,et al.Flea diversity as an element for persistence of plague bacteria in an East African plague focus[J].PLoS One,2012,7(4):e35598.DOI:10.1371/journal.pone.0035598
[13]Wei SZ,Li C,Zheng Y,et al.The epidemiological analysis of Qinghai plague natural foci for mainly fleas media ecology[J].Mod Prevent Med,2010,37(16):3171-3172.(in Chinese)魏紹振,李超,鄭誼,等.青海省鼠疫自然疫源地主要媒介蚤生態及流行病學意義分析[J].現代預防醫學,2010,37(16):3171-3172.
[14]Wang LM,Li GQ,Xu GR,et al.Analysis of epidemic characteristics ofMicrotusfuscusplague in recent years[J].Chin J Ctrl Endem Dis,2004,19(3):178-181.(in Chinese)汪立茂,李光清,許光榮,等.青海田鼠鼠疫流行特點分析[J].中國地方病防治雜志,2004,19(3):178-181.
[15]Wang LM,Li C,Li CX,et al.Study on faunal structure of fleas in shiqu county of Sichuan province[J].JPMI,2002,18(2):99-101.(in Chinese)汪立茂,李超,李存香,等.四川省石渠縣蚤類種群結構研究[J].預防醫學情報雜志,2002,18(2):99-101.
