早期發育滸苔對2株常見赤潮藻的化感效應*
2015-11-28 11:09:20周名江張清春林佳寧
海洋科學進展 2015年4期
關鍵詞:生長
劉 青,顏 天,周名江,張清春,林佳寧
(1.中國科學院 海洋研究所海洋生態與環境科學重點實驗室,山東 青島266071;2.中國科學院大學,北京100049)
自2007年我國黃海海域連續8a發生了大規模的滸苔(Ulvaprolifera)綠潮災害。滸苔綠潮形成后,暴發生物量大,覆蓋面積廣,持續時間久,生態環境效應不容忽視。滸苔與微藻同處于海洋生態系統的初級生產者地位,生態位有一定的重疊,因此不可避免地產生相互競爭。目前普遍認為大型海藻對海域中微型浮游植物的影響是通過多種途徑進行的。滸苔在風浪的影響下脫離固著基,漂浮在海面上,通過遮蔽陽光來影響微藻的生長;它們還能夠快速地吸收營養鹽,與微藻進行營養競爭[1-2];同時它們還能向環境中分泌抑藻物質,抑制微藻的生長,甚至可以防止赤潮的暴發性增殖[3-4]。已有研究表明,形成綠潮的大型海藻可通過化感機制影響海域的生物種類分布與生態格局,從而改變海洋生態系統的結構和演替順序[5-7]。
針對我國黃海海域連年暴發的滸苔綠潮災害的研究工作均表明,滸苔成熟藻體能通過化感作用來抑制微藻的生長,甚至致其死亡[8-11]。滸苔早期發育階段的微觀繁殖體是否具有同樣的影響效應呢?微觀繁殖體作為大型綠藻的早期形成產物,包括孢子、配子、合子這些無細胞壁保護的裸露單細胞及固著萌發后細胞結構較為簡單的萌發幼體。在適宜的附著基和環境條件下,微觀繁殖體會迅速萌發成藻絲體,這個萌發期是滸苔生長發育必經的重要階段[12-14]。Song等[15]和Li等[16]分別在綠潮發生的不同時期,對黃海綠潮形成前后不同時空的滸苔微觀繁殖體進行定量分析,發現微觀繁殖體不僅是綠潮形成前的最初形式,也是綠潮規模擴增的途徑之一。同時作為種子庫為來年滸苔綠潮暴發提供了一定的物質基礎,因此它們在整個綠潮發生發展過程中起到關鍵性的作用。然而關于這些微觀繁殖體對微藻的影響研究目前還未見報道。
自2000年我國赤潮的發生頻率和規模也呈現出急劇擴大的態勢,并且有毒、有害的赤潮原因種也在不斷增加。塔瑪亞歷山大藻可分泌藻毒素,屬于一類危害較嚴重的有毒甲藻。近年來調查發現,由其引發的赤潮在我國南北各大海域均有發生。東海原甲藻是我國海域近年來常見的藻華原因種,因其引發的赤潮暴發規模大,持續時間長而引起了廣泛關注[17-18]。因此,我們在實驗室控制條件下探索了滸苔早期發育階段(微觀繁殖體萌發階段)對這2株赤潮藻類的化感作用,以期豐富滸苔綠潮對海洋浮游植物潛在影響效應這一學科領域的理論研究,并對赤潮和綠潮有關方面的研究提供一定的理論資料。
1 材料和方法
1.1 微藻與滸苔的培養
實驗所用微藻均來自中國科學院海洋研究所海洋生態實驗室。塔瑪亞歷山大藻(Alexandriumtamarense)ATHK藻株于1995年分離自中國南海香港海域,由暨南大學提供;東海原甲藻(Prorocentrumdonghaiens)PDDH藻株于2004年分離自東海長江口海域,由國家海洋局第二海洋研究所提供。將上述微藻分別接種在500mL三角燒瓶中培養至指數生長期待用,培養液采用f/2營養鹽配方,培養溫度為(20±1)℃,光照4 000lx,光暗比為14∶10。實驗海水取自青島太平角的表層海水,經沙濾、沉淀,0.45μm混合纖維濾膜過濾后,再用高壓滅菌鍋滅菌(121℃,20min)。
滸苔(石莼屬綠藻)于2011-07采自青島匯泉灣。將采集的新鮮滸苔去除泥沙,再用滅菌海水漂洗3~4次,鏡檢確認無雜藻后,暫養于f/2培養液中,培養條件和所用海水均與微藻相同。
1.2 滸苔微觀繁殖體的獲得
選取滸苔配子主要是通過形態學觀察(鞭毛數和個體大小)及趨光性檢測來判斷:配子大小約為6μm×3μm,呈長橢圓形,前端透明,后端為綠色的色素體,細胞內一般含有1個蛋白核,頂生2根鞭毛使其可以快速運動;在趨光性檢測中呈現明顯的正趨光。
利用切斷的方法誘導滸苔釋放配子。挑選單株滸苔配子體,將藻體切為1~2cm長的片段置于盛有200mL f/2培養液的三角燒瓶中,誘導2~3d,觀察部分滸苔藻段變為黃色后,單獨挑取黃色藻段放入培養皿中,直至滸苔釋放配子。待培養皿中藻體釋放完畢變為白色后,取出藻段,將2根鞭毛的配子收集。整個過程為防止配子固著,在盡量短的時間內完成操作。將收集的配子液搖勻,顯微計數后,迅速進行實驗。
1.3 實驗方法
1.3.1 滸苔配子在萌發階段對2株赤潮藻生長的影響
實驗中對照組為各赤潮藻單培養組,實驗組分別為滸苔配子與各赤潮藻共培養組,在盛有40mL培養液的小燒杯中進行。實驗開始前1天,將新鮮釋放的滸苔配子(2×104個/mL)接種到含f/2培養液的小燒杯中,避光培養以保證滸苔配子的隨機固著。實驗開始時恢復光照培養便于配子萌發,并加入微藻藻株,微藻的初始密度設置為0.1×104個/mL,每組包含3個重復。每日定時搖動小燒杯,防止微藻貼壁生長。每天取0.2mL培養液,用Lugol's試劑固定后,計數藻細胞密度,同時向小燒杯中加入0.2mL 10倍f/2培養液,以維持培養液體積恒定并避免營養限制。實驗維持10d,在實驗結束時計算微藻的生長抑制率I=(1-N/N0)×100%(N為共培養組中藻細胞密度;N0為單培養組中藻細胞密度)。
1.3.2 不同處理的滸苔在萌發階段對2株赤潮藻生長的影響
1)不同密度的滸苔配子添加組對塔瑪亞歷山大藻生長的影響
實驗同樣采用滸苔配子和微藻共培養系統。滸苔的起始密度分別設為2×104,1×104,0.1×104和0.01×104個/mL,塔瑪亞歷山大藻的初始密度設置為0.1×104個/mL。實驗開始前1天,分別將不同接種密度的滸苔配子避光培養以保證其隨機固著。實驗開始時恢復正常光照培養條件,并加入塔瑪亞歷山大藻藻株,實驗組為滸苔配子與塔瑪亞歷山大藻共培養組,對照組為塔瑪亞歷山大藻單培養組,每組包含3個重復。每隔1d取0.2mL培養液用于計數,同時加入0.2mL 10倍f/2培養液,以維持培養液體積恒定并避免營養限制。實驗維持20d,并分別在第10天和第20天計算微藻生長抑制率。
2)滸苔配子一次性培養濾液對塔瑪亞歷山大藻和東海原甲藻生長的影響
將初始接種密度為2×104個/mL的滸苔配子在小燒杯中培養10d,將培養液經滅菌的0.45μm混合纖維濾膜過濾,并用10倍f/2營養液加富,得到滸苔配子培養濾液。立即分別接種處于對數生長期的塔瑪亞歷山大藻和東海原甲藻,對照組為相同條件下培養于f/2加富海水中的塔瑪亞歷山大藻和東海原甲藻單培養組。微藻的初始密度都設置為0.1×104個/mL。實驗維持20d,并分別在第10天和20天計算各微藻生長抑制率。
3)滸苔半連續培養濾液對塔瑪亞歷山大藻和東海原甲藻生長的影響
實驗組分別為滸苔配子培養過濾液與赤潮微藻共培養組,將塔瑪亞歷山大藻和東海原甲藻接種在滸苔的培養過濾液中,滸苔配子培養過濾液獲取同一次性培養濾液。每隔1d將小燒杯中的培養液移出4mL,然后添加4mL營養加富的新鮮滸苔配子培養濾液以保持培養液體積的恒定。對照組添加4mL f/2加富的滅菌海水,培養條件、營養水平、密度設置與共培養組一致。實驗設置同一次性培養濾液。
4)高溫處理的滸苔配子培養過濾液對塔瑪亞歷山大藻和東海原甲藻生長的影響
將培養滸苔配子的過濾液在高壓滅菌鍋中滅菌,冷卻后分別接種塔瑪亞歷山大藻和東海原甲藻。對照組為相同條件下培養于f/2加富消毒海水中的塔瑪亞歷山大藻和東海原甲藻單培養組。實驗設置同滸苔配子一次性培養濾液。
1.4 數據分析
實驗所得數據均以平均值±標準誤表示(n=3)。采用Excel做圖,統計軟件SPSS 16.0進行單因素方差分析(one-way ANOVA),并用Duncan進行多重比較檢驗和差異顯著性分析,P<0.05被認為是差異顯著。
2 結果與分析
2.1 滸苔配子萌發階段對2株赤潮微藻生長的影響
在與滸苔配子共培養情況下,2株赤潮微藻的生長均受到了一定的影響(圖1)。實驗組中微藻的生物量水平全部低于對照組的,即滸苔配子添加對2株微藻的生長均表現出抑制作用,在第10天對塔瑪亞歷山大藻和東海原甲藻的生長抑制率分別達到了28%和39%,與對照組達到了顯著性差異(P<0.05)。
2.2 不同初始密度的滸苔配子對塔瑪亞歷山大藻生長的影響
共培養條件下不同初始密度下的滸苔配子對塔瑪亞歷山大藻的生長均表現出一定的影響,并隨著滸苔接種密度的增大,其對塔瑪亞歷山大藻生長的抑制作用越加明顯(圖2)。與對照組相比,其中2×104個/mL滸苔配子添加組,塔瑪亞歷山大藻藻細胞指數期增長較為緩慢,然后迅速進入衰亡期,在實驗進行至第22天即完全死亡,所能達到的最高細胞密度僅為對照組的47%;1×104個/mL滸苔配子添加組,塔瑪亞歷山大藻的生命周期也短于對照組,所能達到的最高細胞密度僅為對照組的65%;0.1×104個/mL滸苔配子添加組的抑藻效果稍弱,但赤潮微藻的細胞生長也較為緩慢,所達到的最大細胞密度為對照組的80%。較低密度(0.01×104個/mL)的滸苔配子添加組對塔瑪亞歷山大藻的生長沒有明顯的抑制作用,實驗組與對照組的生長曲線較為相似。
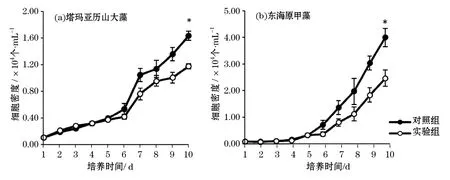
圖1 滸苔配子在萌發階段對2株赤潮微藻生長的影響Fig.1 Effects of U.proliferagametes at germination stage on the growth of two microalgal red tide strains

圖2 不同接種密度的滸苔配子對塔瑪亞歷山大藻生長的影響Fig.2 Effects of different initial densities s of U.proliferagametes on the growth of A.tamarense
2.3 不同處理的滸苔配子在萌發階段對塔瑪亞歷山大藻和東海原甲藻生長的影響
與對照組相比,在實驗進行的第10天和第20天,只有滸苔配子添加組和滸苔配子培養濾液連續添加組顯著抑制了塔瑪亞歷山大藻的生長(P<0.05),而滸苔配子培養濾液一次性添加組未對微藻的生長產生明顯的影響(圖3)。這說明抑藻物質不但存在于滸苔組織中,還可以分泌到培養液中起作用,但是需要連續的添加才能起到較好的抑藻效果。高溫處理過的滸苔配子培養濾液也對塔瑪亞歷山大藻的生長無顯著的抑制作用,表明該抑藻物質不穩定,易在高溫下降解。實驗第10天時,滸苔配子添加組和滸苔配子培養濾液連續添加組對塔瑪亞歷山大藻的生長抑制率為28%和25%,在第20天,對其的生長抑制率分別達到83%和63%。說明隨著共培養實驗的進行,這2個處理組對塔瑪亞歷山大藻的抑制作用不斷加強,而滸苔生長所分泌克生物質的不斷增加應該是導致此現象的直接原因。
與對照組相比,在實驗進行的第10天和20天時,滸苔配子添加組和滸苔配子培養濾液連續添加組均對東海原甲藻的生長產生了明顯的抑制作用(P<0.05),而滸苔配子培養濾液一次性添加組和高溫處理過的滸苔配子培養濾液添加組均未表現出抑藻效果(圖4)。這也說明該滸苔分泌的抑藻物質容易隨著時間或在高溫下降解。比較發現,第10天2個實驗組對微藻的生長抑制率分別為39%和24%,滸苔配子添加組比滸苔配子培養濾液添加組對東海原甲藻生長的抑制作用更明顯。第20天時,這2個不同的處理組對東海原甲藻生長的抑制率分別達到了40%和27%,稍高于第10天的。

圖3 不同處理組的滸苔配子在萌發階段對塔瑪亞歷山大藻生長的影響Fig.3 Effects of U.proliferagametes at germination stage on the growth of A.tamarense
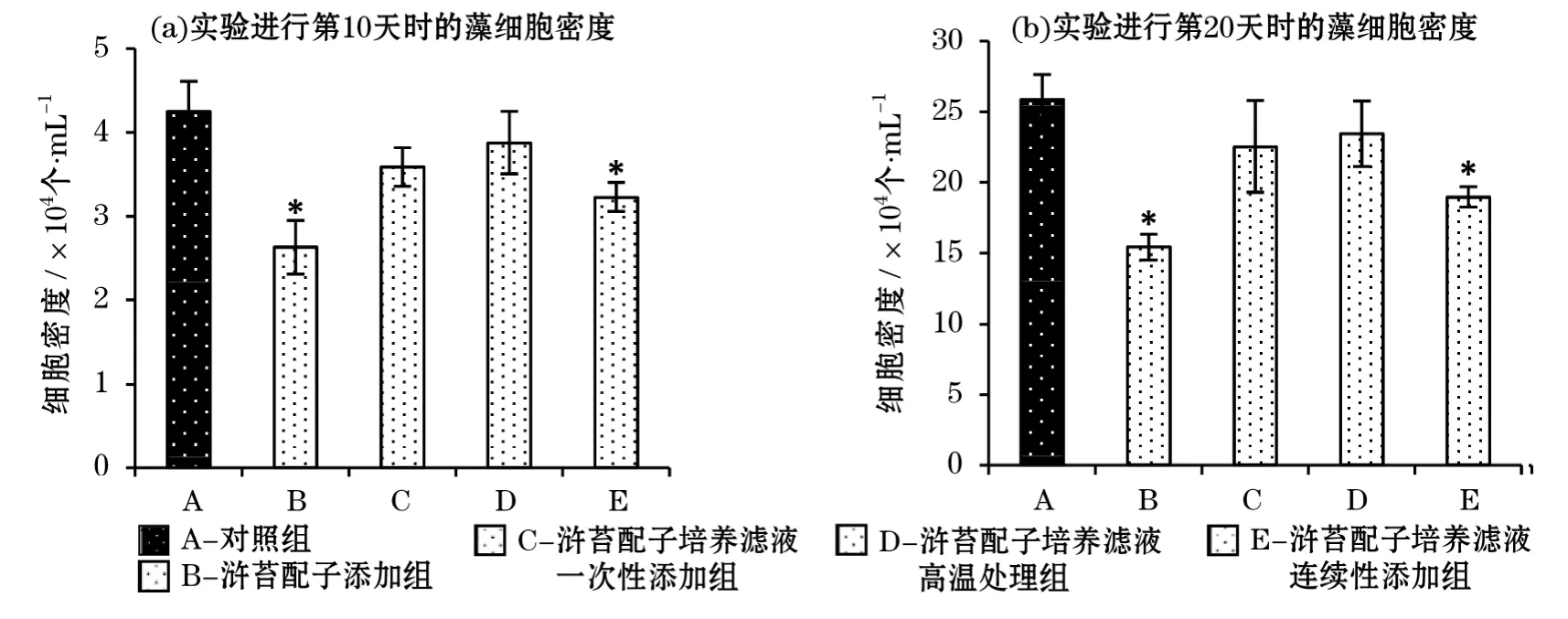
圖4 不同處理組的滸苔配子在萌發階段對東海原甲藻生長的影響Fig.4 Effects of U.proliferagametes at germination stage on the growth of P.donghaiense
3 討論
藻類的化感作用是普遍存在的現象,在浮游植物種間競爭、群落演替甚至赤潮發生發展消亡過程中均發揮著重要作用,已引起了研究人員的廣泛關注。石莼屬綠藻對海洋微藻的影響是由多種作用機制造成的,化感作用和營養競爭是其中2種最重要的機制。化感作用,是一種生物通過向環境釋放化學物質而對其附近的另一些生物產生直接或間接的、有利或有害作用的現象[19]。近年來我國黃海海域大面積的滸苔綠潮與赤潮藻的化感作用研究表明,滸苔成熟藻體會通過分泌化感物質來影響微藻的生長[8-11]。我們在實驗室控制條件下進行的研究,首先排除了溫度、光照等環境因素的變化可能對實驗結果產生的影響,其次在實驗過程中通過添加充足的營養鹽來避免營養競爭。因此,我們可以判定化感作用應是引起這種影響效應的根本原因。而且滸苔在早期微觀繁殖體萌發階段即可通過化感效應抑制某些赤潮微藻的生長,并不局限于成熟的藻體階段。
滸苔配子在萌發階段,就可以影響赤潮微藻塔瑪亞歷山大藻和東海原甲藻的生長,并且隨著初始接種密度的升高,其對塔瑪亞歷山大藻的抑制作用逐漸加強。配子添加組和濾液連續添加組均對這2株赤潮藻的生長產生了明顯的抑制作用。這說明滸苔微觀繁殖體組織中確實存在著抑藻物質,并且可以分泌到培養液中起作用,但是抑制效應會隨著時間的進行而減弱,需要連續的添加累積才能起到較好的抑藻效果。滸苔成體的抑藻效應也有相同的報道,研究者認為化感物質具有一定的累積效應,當累積到一定的閾值時這種效應才會顯現[9]。因此實驗中化感物質的連續分泌及添加是抑制微藻生長的關鍵。高溫處理過的滸苔濾液對2種微藻的生長均無顯著的抑制作用,表明該抑藻物質不穩定,易在高溫下降解。霍元子等[8]也曾報道新鮮的滸苔藻體培養濾液在經高溫后對某些微藻抑制作用消除的現象。
隨著對石莼屬綠藻化感作用研究的日益增多,關于其分泌的化感物質也已經形成了一些初步的看法。Alamsjah等[20]從孔石莼中分離出抑制赤潮異彎藻的活性物質,主要成分為多不飽和脂肪酸;Jin和Dong[21]也認為不飽和脂肪酸可能是孔石莼產生的克生物質的重要組成成分之一。夏鈺妹[22]從腸滸苔的有機相提取物中分離純化出3個活性化合物,可能的抑藻成分被認為是α-亞麻酸。韓秀榮等[10]認為滸苔分泌的化感物質具有相對較高的極性;孫穎穎等[23]對滸苔抑藻組分進行進一步分離,發現其含有內酯和香豆素類化合物。這都表明石莼屬不同藻種分泌的化感物質是不同的,這種差異性可能與報道的化感物質含有多種成分有關。Jensen[24]指出大型海藻的微觀繁殖體在早期的固著萌發階段,會通過釋放一定的代謝產物來影響周圍的微環境。Amsler和Searles的研究[25]更表明大型海藻一些特性的形成,是在其微觀繁殖體階段就建立的。我們的實驗結果符合這種效應,說明滸苔能夠分泌化感物質的特性在早期發育階段就已經形成,至于滸苔微觀繁殖體是否同成熟藻絲體分泌相同的抑藻物質,還有待進一步實驗證實。
對比滸苔微觀繁殖體與成熟藻體的抑藻試驗結果,我們發現微觀繁殖體在萌發階段的第10天對塔瑪亞歷山大藻的生長抑制率為28%,而生物量為1.0g/L的滸苔成熟藻絲體在實驗進行第10天時對塔瑪亞歷山大藻的生長抑制率大約為50%[8]。滸苔微觀繁殖體在本實驗進行的第10天僅剛完成萌發過程,細胞結構還較為簡單,生物量顯著低于成熟藻體的1.0g/L。因此倘若換算成同等生物量,其對微藻產生的抑制作用并不低于滸苔藻絲體,所以滸苔微觀繁殖體的化感作用是不容忽視的。滸苔綠潮在其整個孕育、發生發展及消亡過程中,都有大量微觀繁殖體的形成。因此,要全面地評價滸苔綠潮的生態效應,就不僅要從滸苔暴發后形成大量生物量遮蔽陽光,快速吸收營養鹽,消亡時腐爛分解的影響方面考慮,還要考慮大量滸苔微觀繁殖體孕育階段可能造成的影響。我們的研究結果豐富并證實了滸苔綠潮潛在的生態效應。另外,滸苔對常見赤潮藻種生長的影響效應,可能會降低赤潮發生的幾率,因此在開展赤潮的生物防治以及海域的生態修復等方面都有著積極的意義。
(References):
[1]SFRISO A,PAVONI B.Macroalgae and phytoplankton competition in the central Venice lagoon[J].Environmental Technology,1994,15(1):1-14.
[2]BARCO J W,JAMES W F.Effects of submerged aquatic macrophytes on nutrient dynamics,sedimentation,and resuspension[C]∥JEPPESEN E,S?NDERGAARD M,CHRISTOFFERSEN K.The structuring role of submerged macrophytes in lakes.New York:Springer,1998:197-217.
[3]ANDERSON D M.Turning back the harmful red tide[J].Nature,1997,388:513-514.
[4]JEONG J H,JIN H J,SOHN C H,et al.Algicidal activity of the seaweedCorallinapiluliferaagainst red tide microalgae[J].Journal of Applied Phycology,2000,12(1):37-43.
[5]JASSER I.The influence of macrophytes on a phytoplankton community in experimental conditions[J].Hydrobiologia,1995,306(1):21-32.
[6]FLETCHER R T.The occurrence of'green tide'[C]∥SCHRAMM W,NIENHUIS P H.Marine benthic vegetation-recent changes and the effects of eutrophication.Berlin:Springer,1996:7-43.
[7]KOMER S,NICKIISCH A.Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes[J].Journal of Phycology,2002,38(5):862-871.
[8]HUO Y Z,TIAN Q T,XU S N,et al.Allelopathic effects ofUlvaproliferaon growth ofKareniamikimotoi[J].Marine Environmental Science,2010,29(4):496-508.霍元子,田千桃,徐珊楠,等.滸苔對米氏凱倫藻生長的克生作用[J].海洋環境科學,2010,29(4):496-499.
[9]JIA R,WU M,CAI C E,et al.Allelopathic effects ofUlvaproliferaonHeterosigmaakashiwo[J].Journal of Fisheries of China,2012,36(4):562-567.賈睿,吳敏,蔡春爾,等.滸苔對赤潮異灣藻的克生作用[J].水產學報,2012,36(4):562-567.
[10]HAN X R,GAO S,HOU J N,et al.Allelopathic effects of extracts fromUlvaproliferapowders on the growth ofProrocentrum donghaienseandSkeletonemacostatum[J].Acta Ecologica Sinica,2013,33(23):7417-7429.韓秀榮,高嵩,侯俊妮,等.滸苔干粉末提取物對東海原甲藻和中肋骨條藻的克生作用[J].生態學報,33(23):7417-7429.
[11]WANG R J,WANG Y,ZHOU J,et al.Algicidal activity ofUlvapertusaandUlvaproliferaonProrocentrumdonghaienseunder laboratory conditions[J].African Journal of Microbiology Research,2013,7(34):4389-4396.
[12]CHAPMAN A R O.Population and community ecology of seaweeds[C]∥BLAXTER J H S,SOUTHWOOD A J.Advances in Marine Biology.London:Academic Press,1986:1-161.
[13]CLAYTON M N.Propagules of marine macroalgae:structure and development[J].British Phycological Journal,1992,27(3):219-232.
[14]SANTELICES B,AEDO D,HOFFMANN A.Banks of microscopic forms and survival to darkness of propagules and microscopic stages of macroalgae[J].Revista Chilena de Historia Natural,2002,75:547-555.
[15]SONG W,LI Y,FANG S,et al.Temporal and spatial distributions of green algae micro-propagules in the coastal waters of the Subei Shoal,China[J].Estuarine,Coastal and Shelf Science,2014,doi:1016/j.cess.2014.08.006.
[16]LI Y,SONG W,XIAO J,et al.Tempo-spatial distribution and species diversity of green algae micro-propagules in the Yellow Sea during the large-scale green tide development[J].Harmful Algae,2014,39:40-47.
[17]QI Y Z,WANG Y.What the Prorocentrum species should be[J]?Chinese Journal of Applied Ecology,2003,14(7):1188-1190.齊雨藻,王艷.我國東海原甲藻赤潮應屬哪種[J]?應用生態學報,2003,14(7):1188-1190.
[18]ZHOU M J,YAN T,ZOU J Z.Preliminary analysis of the characteristics of red tide areas in Changjiang River estuary and its adjacent sea[J].Chinese Journal of Applied Ecology,2003,14(7):1031-1038.周名江,顏天,鄒景忠.長江口鄰近海域赤潮發生區基本特征初探[J].應用生態學報,2003,14(7):1031-1038.
[19]RICE E L.Allelopathy[M].Orlando,Florida:Academic Press,1984.
[20]ALAMSJAH M A,HIRAO S,ISHIBASHII F,et al.Isolation and structure determination of algicidal compounds fromUlvafasciata[J].Bioscience,Biotechnology,and Biochemistry,2005,69(11):2186-2192.
[21]JIN Q,DONG S L.Comparative studies on the allelopathic effects of two different strains ofUlvapertusaonHeterosigmaakashiwoand Alexandrium tamarense[J].Journal of Experimental Marine Biology and Ecology,2003,293(1):41-55.
[22]XIA Y M.Inhibitory effects and the mechanism of the marine algaeEnteromorphaintestinalison the growth of the red tide microalgae[D].Ningbo:Ningbo University,2012.夏鈺妹.大型綠藻滸苔對赤潮藻的抑制作用及其機理研究[D].寧波:寧波大學,2012.
[23]SUN Y Y,DONG X K,LI G,et al.Isolation of antialgal substances fromEnteromorphaproliferaand its growth inhibition on the red tide microalgae[J].Journal of Fisheries of China,2014,38(6):813-819.孫穎穎,董曉柯,李光,等.滸苔抑藻物質的分離及其對赤潮微藻的抑制作用[J].水產學報,2014,38(6):813-819.
[24]JENSEN A.Chemical conditioning of seawater by algal growth and development[C]∥FAULKNER D J,FENICAL W H.Marine Natural Products Chemistry,New York:Plenum Press,1977:329-344.
[25]AMSLER C D,SEARLES R B.Vertical distribution of seaweed spores in a water column offshore of North Carolina[J].Journal of Phycology,1980,16(4):617-619.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
