BikDD腫瘤靶向基因治療抑制淋巴瘤細胞生長研究*
2015-12-06 02:32:14李建璜
成都醫學院學報 2015年1期
曹 軍,羅 寧,李建璜△
1.中南大學湘雅醫學院附屬腫瘤醫院 胸部內二科(長沙 410013);2.中南大學湘雅醫院 腫瘤科(長沙 410008)
惡性淋巴瘤是危害人類健康的重要疾病之一,其主要治療方法為化學藥物治療和放射治療,但上述方法對于進一步提高其生存率作用有限。隨著基因工程技術的飛速發展,基因治療已成為淋巴瘤治療研究的熱點,并取得了較多的成果,但目的基因表達靶向性差及表達效率低是限制其臨床應用的關鍵問題。本課題組成員前期研究已成功構建出能在淋巴瘤細胞中高效特異性表達目的基因的S-VISA載體[1],很好地解決了以上問題。因而,如何選擇有效的治療基因成為淋巴瘤基因治療的關鍵。
細胞凋亡通路障礙是淋巴瘤發生發展過程中最重要的分子事件之一,因此,誘導腫瘤細胞凋亡成為淋巴瘤基因治療的思路之一。Bcl-2家族包括凋亡抑制蛋白Bcl-2、Bcl-XL、Bcl-w、Mcl-1等,促凋亡蛋白質如 Bik、Bad、Bax等[2]。Bik(Bcl-2interacting killer)作為Bcl-2家族中一種僅含BH3結構域的促凋亡蛋白質,能結合的Bcl-2家族凋亡抑制蛋白最多,且與 Bcl-2及 Mcl-1的結合力最強[3]。既往研究[4-6]表明,將Bik的第33位蘇氨酸殘基和第35位絲氨酸殘基替換為門冬氨酸,使其成為突變型Bik(BikDD),BikDD在肺癌、乳腺癌及前列腺癌等惡性腫瘤中促進細胞凋亡和抑制細胞生長的能力比野生型Bik更強。
本實驗擬構建含BikDD、Survivin啟動子及VISA系統的表達質粒S-VISA-BikDD,將其轉染淋巴瘤細胞株,研究其在淋巴瘤細胞中的作用。
1 材料與方法
1.1 材料
1.1.1 質粒 pGL3-Survivin質粒由本科室唐友紅博士惠贈[7],pCMVgDWPRE質粒由意大利帕爾馬大 學 Dr Gaetano Donofrio 惠 贈[8],TSTA-NSN質粒由美國加州大學洛杉磯分校醫學院Dr Mike Carey惠 贈[9],pGL3-CMV、pGL3-Basic質 粒 購 自Promega公司。
1.1.2 細胞 人淋巴瘤 Ramos、U937、Raji細胞株購自上海麥莎生物科技有限公司,人肝細胞株Chang Liver購自中科院上海生命科學院細胞所。
1.1.3 試劑 實驗所用內切酶均為NEB產品,連接酶(寶靈曼B.M 公司),DNA連接酶、luciferase assay system試劑盒(美國Promega公司)。新生小牛血清(杭州四季青生物公司),DMEM、RPMI 1640培養基(美國Gibco公司),質粒小量提取試劑盒(美國Omega公司)、膠回收試劑盒(TaKaRa公司),基因定點突變試劑盒(碧云天生物技術研究所),鼠抗人Bik單克隆抗體(臺灣Abnova公司)。
1.2 方法
1.2.1 pGL3-Bik載體的構建 根據 GenBank的Bik基因序列設計引物,上游引物序列為:5'CGAAGATCTATGTCTGAAGTAAGACC 3'(包含起始密碼子,并在5'端引入BgiⅡ酶切位點);下游引物序列為:5'CCAAGCTTTCACTTGAGCAGC AGGT 3'(在5'端引入 HindⅢ酶切位點),引物由上海英駿生物技術有限公司合成。Trizol法提取人類腸基因組RNA,以此為模板,PCR擴增Bik基因,反應條件:94℃預變性4min,94℃30s,53℃30 s,72℃30s,30個循環,72℃延伸8min。PCR產物用1.0% 瓊脂糖電泳檢測,觀察結果。用BgiⅡ/HindⅢ對PCR產物進行雙酶切后純化回收,連入同樣用BgiⅡ/HindⅢ雙酶切的pGL3-Basic載體,構建成pGL3-Bik載體。利用PCR及 HindⅢ/BgiⅡ雙酶切鑒定后送上海英駿生物技術有限公司純化測序。
1.2.2 pGL3-BikDD 載體的構建 根據 GenBank的Bik基因序列,為實現將Bik基因編碼蛋白第33位蘇氨酸殘基和第35位絲氨酸殘基替換為門冬氨酸,對其編碼基因進行定點突變,分別將第33位蘇氨酸殘基編碼基因ACT突變為GAC,第35位絲氨酸殘基編碼基因TCT突變為GAT。引物設計如下:上游引物5'GTTCTTGGCATGGACGAC GATGAAGAGGACC3',下游 引物 5'GGTCCTCT TCATCGTCGTCCATGCCAAGAAC 3'。以 pGL3-Bik質粒為模板,對Bik基因編碼序列進行PCR擴增,反應條件:95℃預變性1min,95℃40s,60℃60s,68℃5min,18個循環,72℃延伸8min。擴增產物轉化感受態大腸桿菌DH5α,氨卞抗性篩選后提取質粒,KpnI/NheI酶切鑒定,進一步測序證實pGL3-BikDD構建成功。
1.2.3 pGL3-CMV-BikDD載體的構建 BgiⅡ和HindⅢ雙酶切pGL3-BikDD和pGL3-CMV載體,回收約500bp的BikDD片段和約5.4kb的pGL3-CMV載體片段。用T4DNA連接酶連接,構建成pGL3-CMV-BikDD載體。利用Bik引物進行PCR篩選及雙酶切(BgiⅡ/HindⅢ)鑒定。
1.2.4 S-VISA-BikDD 載體的構建 1)G5E4TBikDD-WPRE載體的構建:BgiⅡ和HindⅢ雙酶切pGL3-BikDD和 G5E4T-WPRE載體,回收約500 bp的BikDD片段和約5.6kb的G5E4T-WPRE載體片段。用T4DNA連接酶連接,構建成G5E4TBikDD-WPRE過渡載體。利用Bik引物進行PCR篩選和雙酶切(BgiⅡ/HindⅢ)鑒定。2)S-VISABikDD載體的構建:KpnI和SalI雙酶切S-TSTA和G5E4T-BikDD-WPRE載體,回收約4.8kb的STSTA片段(保留Survivin啟動子、除G5E4T外的TSTA系統和部分載體序列)和約3.3kb的G5E4T-BikDD-WPRE片段(保留 G5E4T、BikDD、熒光素酶編碼基因、WPRE和poly A序列)。用T4 DNA連接酶連接,構建成S-VISA-BikDD載體,分別利用G5E4T和Bik引物進行PCR篩選及雙酶切(KpnI/SalI)鑒定。
1.2.5 質粒轉染 將培養至對數生長期的Ramos、Raji、U937和chang liver細胞與S-VISA/CMV-BikDD/S-VISA-BikDD/pGL3-basic 質 粒 分別溶于無血清RPMI 1640的電穿孔介質中,將細胞懸液移入石英電轉杯中,電轉條件:電壓270V,穿孔時間14.4ms。電轉完成后立即將細胞懸液轉移至25cm2培養瓶中,加入含10%胎牛血清的RPMI 1640培養液,置于37℃5%CO2培養箱中,48h后終止培養,收獲細胞。
1.2.6 Western blot檢測Bik蛋白表達 分別收集質粒轉染前及轉染48h后各組細胞,加入RIPA蛋白裂解液裂解細胞抽提蛋白,BCA法測定蛋白濃度,蛋白變性后取60μg總蛋白進行10%SDS-聚丙烯酰胺凝膠電泳分離,蛋白通過濕轉法轉移至硝酸纖維膜上。一抗(鼠抗人Bik單克隆抗體)4℃孵育過夜,TBST洗膜3次,二抗為辣根過氧化物酶(horseradish peroxidase,HRP)標記的羊抗鼠抗體,室溫孵育1h,TBST洗膜3次,化學發光試劑ECL(enhanced chemiluminescence)發光、顯影和定影。
1.2.7 流式細胞儀檢測Raji細胞凋亡 離心、收集轉染S-VISA、CMV-BikDD、S-VISA-BikDD質粒48h后的Raji細胞1×106個/mL,加入PI室溫避光孵育40min,流式細胞儀分析檢測細胞凋亡率。
1.2.8 MTT 實驗取轉染S-VISA、CMVBikDD、S-VISA-BikDD質粒48h后的細胞,各孔加入 MTT溶液(5mg/mL)20μL,放入培養箱繼續培養4h后棄上清液,再加入DMSO 150μL/孔,充分振蕩15min溶解結晶,用酶標儀在570nm波長處測定其吸光度(OD)值。以轉染pGL3-basic的細胞為對照組,抑制率=(對照組OD值-質粒轉染組OD值)/對照組OD值×100%。
1.3 統計學方法
采用SPSS 19.0統計軟件對各組數據進行分析比較。本實驗中所有數據均采用均數±標準差(±s)表示。多組間比較采用One-Way ANOVA檢驗,多個樣本均數間兩兩比較采用S-N-Kq檢驗法,相關分析采用直相關計算Pearson相關系數,P<0.05為差異有統計學意義。
2 結果
2.1 pGL3-BikDD的鑒定
以人類腸基因組RNA為模版,PCR擴增Bik基因后連入pGL3-basic載體,構建成pGL3-Bik載體。利用定點突變技術將第33位蘇氨酸殘基編碼基因ACT突變為GAC,第35位絲氨酸殘基編碼基因TCT突變為GAT,即分別將第97~99及103~105位的堿基突變,PCR酶切鑒定結果(圖1)。測序結果與GeneBank上公布的Bik基因序列一致。

圖1 pGL3-BikDD載體酶切鑒定圖
2.2 CMV-BikDD載體的鑒定
重組載體CMV-BikDD的酶切鑒定。BgiⅡ和HindⅢ雙酶切pGL3-BikDD和pGL3-CMV載體,回收BikDD片段和pGL3-CMV載體片段。用T4 DNA連接酶連接,構建成 pGL3-CMV-BikDD 載體。雙酶切鑒定結果(圖2)。
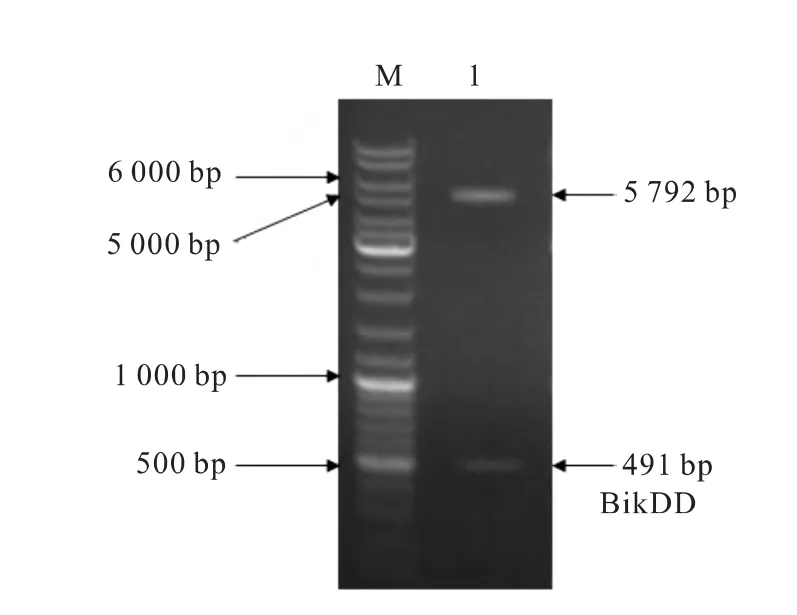
圖2 CMV-BikDD載體酶切鑒定
2.3 S-VISA-BikDD載體的構建
重組載體S-VISA-BikDD的構建及酶切鑒定。BgiⅡ和 HindⅢ雙酶切pGL3-BikDD和 G5E4TWPRE載體,回收的BikDD片段和G5E4T-WPRE片段用T4DNA連接酶連接,構建成G5E4TBikDD-WPRE過渡載體。再經KpnI和SalI雙酶切S-TSTA和G5E4T-BikDD-WPRE載體后,回收到約4.8kb的 S-TSTA片段和約3.3kb 的G5E4T-BikDD-WPRE片段,進一步用T4DNA連接酶連接,PCR雙酶切鑒定結果(圖3)。

圖3 S-VISA-BikDD載體酶切鑒定
2.4 轉染前后淋巴瘤細胞和肝細胞的Bik表達水平
收集S-VISA-BikDD轉染前、后的淋巴瘤細胞Raji、Ramos和 U937和肝細胞chang liver作Western blot檢測。轉染前的淋巴瘤細胞中Bik蛋白表達均低于chang liver細胞,轉染后的淋巴瘤細胞中Bik蛋白表達均較前有上升,但仍低于chang liver細胞的Bik蛋白表達水平,而chang liver細胞在BikDD處理前后無明顯變化(圖4)。

圖4 S-VISA-BikDD載體轉染前后淋巴瘤細胞和肝細胞的Bik表達水平
2.5 S-VISA-BikDD對淋巴瘤細胞的增殖抑制作用
MTT實驗結果顯示:轉染S-VISA-BikDD對于淋巴瘤細胞Raji、Ramos和U937有明顯生長抑制作用,并且均強于CMV-BikDD,差異有統計學意義(P<0.05);對于chang liver細胞無顯著生長抑制作用,差異無統計學意義(P>0.05)。轉染SVISA對于淋巴瘤細胞Raji、Ramos、U937、hut78和chang liver細胞均無生長抑制作用,差異無統計學意義(P>0.05)。轉染CMV-BikDD對于淋巴瘤細胞Raji、Ramos、U937和chang liver細胞均有顯著生長抑制作用,差異有統計學意義(P<0.05)(圖5)。

圖5 S-VISA-BikDD與CMV-BikDD轉染對淋巴瘤和肝細胞的生長抑制作用比較
2.6 流式細胞儀檢測細胞凋亡
為探討上述載體轉染對Raji細胞凋亡的影響,筆者用流式細胞儀檢測了Raji、Ramos、U937和chang liver 4種細胞的凋亡情況,結果顯示:對照組(2.67±0.31)%;S-VISA 組(7.34±0.76)%,SVISA-BikDD組(28.93±4.25)%,CMV-BikDD組(17.62±2.17)%。結果表明,S-VISA-BikDD 及CMV-BikDD轉染對淋巴瘤Raji細胞有明顯凋亡誘導作用,且S-VISA-BikDD對于Raji細胞的凋亡誘導作用較CMV-BikDD明顯,差異有統計學意義(P<0.05)。
3 討論
Bcl-2家族是細胞凋亡的關鍵調節因子,Bik是其中一種重要的促凋亡蛋白。最新研究發現,伴隨著死亡信號的刺激,Bik磷酸化后,可以與抗凋亡蛋白質結合,從而使其他促凋亡蛋白直接激活Bax。Bax移位到線粒體并形成同源二聚體通道,促進細胞色素c的釋放,導致caspase的激活和細胞凋亡[10]。
Sturm等[11]研究顯示,Bik高表達于正常腎上皮中,而腎細胞癌中由于染色體22q13.2中的Bik位點雜合性丟失及表觀遺傳學啟動子沉寂而導致Bik基因表達失活。Van Keimpema等[12]闡明彌漫大B細胞淋巴瘤患者的生存率低與Bik蛋白編碼基因表達減低相關。Brosseau等[13]發現,治療套細胞淋巴瘤的一種新的有效藥物來那度胺與1α,25-二羥維生素D3聯合應用,明顯促進了腫瘤細胞死亡,其作用與Bik表達上調相關,這種表達上調與Bik mRNA水平增長有關。Bik基因沉寂阻止了這兩種藥物誘導的凋亡,證實Bik直接參與細胞死亡。Hong等[14]在 MCF-7和 MDA-MB231人類乳腺癌細胞系中發現植物血凝素與阿霉素聯合應用,促進癌細胞凋亡,同時檢測到包括Bik蛋白在內的促凋亡蛋白表達上升。Guo等[15]通過對濾泡性淋巴瘤患者基因表達譜的分析,鑒定出包括Bik在內的幾種基因在zeste同源物2(EZH2)啟動子突變型病例中其mRNA表達水平降低。本實驗結果顯示,淋巴瘤細胞Raji、Ramos及U937的Bik蛋白表達水平與作為正常對照的肝細胞chang liver相比顯著降低,與前述研究結果一致。
Xie等[16]構建了含 BikDD、hTERT 啟動子及VISA系統的腫瘤靶向治療基因T-VISA-BikDD,將其導入多種乳腺癌鼠模型中,發現該治療基因可引起有效的抗腫瘤效應,延長病鼠生存期。近來,Xie等[17]在前列腺癌鼠模型中進行腫瘤靶向性基因治療研究,將T-VISA-BikDD載體進一步聯合雄激素反應區域(ARR)構建成AT-VISA-BikDD載體,導入前列腺癌鼠模型,抑制了前列腺癌生長并延長病鼠存活期,且對機體無明顯毒副作用。以上實驗均證實,T-VISA作為啟動子的毒性作用明顯低于CMV 啟動子。本實驗將 S-VISA-BikDD、CMVBikDD和對照S-VISA瞬時轉染人淋巴瘤細胞Raji、Ramos、U937和肝細胞chang liver后,筆者發現S-VISA-BikDD及CMV-BikDD對于上述3種淋巴瘤細胞均有明顯生長抑制作用,S-VISA-BikDD的生長抑制作用強于 CMV-BikDD(P<0.05);SVISA-BikDD轉染對于chang liver細胞無明顯生長抑制作用(P>0.05),但 CMV-BikDD 對 chang liver細胞表現出抑制作用(P<0.05)。本研究結果表明,S-VISA載體選擇性地驅動BikDD在淋巴瘤細胞中的表達,實現了BikDD在淋巴瘤細胞中的高效、特異性表達,避免了對正常細胞的急性毒性作用。凋亡檢測的結果進一步證實了S-VISA-BikDD特異性促淋巴瘤細胞凋亡的作用,其作用較CMVBikDD明顯增強。因此,對于淋巴瘤的基因治療而言,S-VISA-BikDD可能比 CMV-BikDD更為有力,且對正常細胞的毒性低,與以上實驗結果一致。
綜上所述,S-VISA-BikDD載體的成功構建及其促凋亡作用研究,為淋巴瘤基因治療找到了新的思路及靶標,為進一步開展動物實驗和臨床實驗的可行性及安全性提供了可靠依據,具有重要的理論意義和應用前景。
[1]羅寧,肖志強,李建璜.淋巴瘤特異性高效基因表達載體survivin-VISA的構建與鑒定[J].腫瘤防治研究,2014,41(7):724-727.
[2]Yip KW,Reed JC.Bcl-2family proteins and cancer[J].Oncogene,2008,27(50):6398-6406.
[3]Garg H,Salcedo R,Trinchieri G,et al.Improved nonviral cancer suicide gene therapy using survivin promoter-driven mutant Bax[J].Cancer Gene Therapy,2010,17(3):155-163.
[4]Sher YP,Liu SJ,Chang CM,et al.Cancer-Targeted BikDD Gene Therapy Elicits Protective Antitumor Immunity against Lung Cancer[J].Mol Cancer Ther,2011,10(4):637-647.
[5]Lang JY,Hsu JL,Meric-Bernstam F,et al.BikDD Eliminates Breast Cancer Initiating Cells and Synergizes with Lapatinib for Breast Cancer Treatment[J].Cancer Cell,2011,20(3):341-356.
[6]Xie X,Xia W,Li Z,et al.Targeted Expression of BikDD Eradicates Pancreatic Tumors in Noninvasive Imaging Models[J].Cancer Cell,2007,12(1):52-65.
[7]Tang YH,Zhong MZ,Liu W,et al.To clone human Survivin gene promoter and research the transcriptional activity of it in the lymphoma cells[J].Bai Xue Bing Lin Ba Liu,2010,19(2):84-87.
[8]Donofrio G,Cavirani S,Vanderplasschen A,et al.Recombinant bovine herpesvirus 4(BoHV-4)expressing glycoprotein D of BoHV-1is immunogenic and elicits serumneutralizing antibodies against BoHV-1in a rabbit model[J].Clinical and Vaccine Immunology,2006,13(11):1246-1254.
[9]Zhang L,Adams JY,Billick E,et al.Molecular engineering of a Two-step transcription amplification(TSTA)system for transgene delivery in Prostate cancer[J].Molecular Therapy,2002,5(3):223-232.
[10]Garg H,Salcedo R,Trinchieri G,et al.Improved nonviral cancer suicide gene therapy using survivin promoter-driven mutant Bax[J].Cancer Gene Therapy,2010,17(3):155-163.
[11]Sturm I,Stephan C,Gillissen B,et al.Loss of the tissuespecific proapoptotic BH3-only protein Nbk/Bik is a unifying feature of renal cell carcinoma[J].Cell Death Differ,2006,13(4):619-627.
[12]Van Keimpema M,Grüneberg LJ,Mokry M,et al.FOXP1 directly represses transcription of proapoptotic genes and cooperates with NF-κB to promote survival of human B cells[J].Blood,2014,124(23):3431-3440.
[13]Brosseau C,Dousset C,Touzeau C,et al.Combination of lenalidomide with vitamin D3induces apoptosis in mantle cell lymphoma via demethylation of BIK[J].Cell Death Dis,2014,5:1389.
[14]Hong CE,Park AK,Lyu SY.Synergistic anticancer effects of lectin and doxorubicin in breast cancer cells[J].Mol Cell Biochem,2014,394(1-2):225-235.
[15]Guo S,Chan JK,Iqbal J,et al.EZH2mutations in follicular lymphoma from different ethnic groups and associated gene expression alterations[J].Clin Cancer Res,2014,20(12):3078-3086.
[16]Xie X,Li L,Xiao X,et al.Targeted expression of BikDD eliminates breast cancer with virtually no toxicity in noninvasive imaging models[J].Mol Cancer Ther,2012,11(9):1915-1924.
[17]Xie X,Kong Y,Tang H,et al.Targeted BikDD expression kills androgen-dependent and castration-resistant prostate cancer cells[J].Mol Cancer Ther,2014,13(7):1813-1825.
