續斷菊與玉米間作系統不同植物部位Cd、Pb分配特征
2015-12-06 09:43:39譚建波陳興郭先華李元祖艷群
生態環境學報 2015年4期
關鍵詞:質量
譚建波,陳興,郭先華,李元,祖艷群
云南農業大學資源與環境學院,云南 昆明 650201
續斷菊與玉米間作系統不同植物部位Cd、Pb分配特征
譚建波,陳興,郭先華,李元,祖艷群*
云南農業大學資源與環境學院,云南 昆明 650201
采用植物修復的方法對會澤鉛鋅礦周圍受Cd、Pb污染的農用地進行修復,將超富集植物續斷菊和農作物玉米在污染農用地間作種植,以減少農用地重金屬含量,并獲得合格的農產品。通過田間小區直接種植的方式,測定植物生物量及各器官重金屬含量,研究續斷菊(Sonchus asper L.Hill)與玉米(Zea mays L.)間作系統植物不同器官Cd、Pb分配特征。結果表明,(1)間作續斷菊生物量相比單作增加了31.55%,間作玉米生物量相比單作增加了29.02%。(2)從拔節期到成熟期,間、單作玉米各器官Cd質量分數呈下降趨勢。成熟期與拔節期相比,間作玉米根、莖、葉Cd質量分數分別降低了24.51%、29.06%、55.32%,單作玉米根、莖、葉 Cd質量分數分別降低了 22.05%、7.20%、45.02%。在不同時期,間作玉米根、葉Cd質量分數都低于單作玉米根、葉Cd質量分數。根部Cd質量分數在抽穗期和成熟期差異顯著;葉部Cd質量分數在成熟期差異性顯著;莖部Cd質量分數在抽穗期和成熟期呈現出單作大于間作,在成熟期差異顯著。玉米各部位Pb質量分數呈現出先增加后下降趨勢,且成熟期根、莖Pb質量分數小于拔節期,而該期葉Pb質量分數則大于拔節期。不同時期,間作玉米根、葉Pb質量分數小于單作玉米根、葉,根在抽穗期和成熟期Pb質量分數差異性顯著;間作玉米籽粒Cd、Pb質量分數顯著小于單作。(3)續斷菊根部及地上部Cd質量分數隨時間推移而呈現逐漸增加趨勢,間作續斷菊根部及地上部Cd質量分數分別增加16.88、15.45 mg·kg-1,單作續斷菊根部及地上部Cd質量分數分別增加5.5、10.09 mg·kg-1,間作續斷菊地上部Cd質量分數顯著大于單作,間作根部大于單作根部,在抽穗期和成熟期出現顯著差異。(4)間作、單作種植模式下土壤Cd、Pb質量分數均有下降,土壤Cd質量分數在間作模式下降25.99%,單作續斷菊模式下降23.19%,單作玉米模式下降14.78%;土壤Pb質量分數在間作模式下降6.78%,單作續斷菊模式下降7.53%,單作玉米模式下降7.23%。結果表明,續斷菊和玉米間作促進了續斷菊各部位對鎘的積累量,同時抑制了玉米各部位鎘的積累量。
玉米;續斷菊;間作;Cd;Pb;土壤修復
土壤重金屬污染產生的主要原因是采礦、冶煉、農業等人為因素以及地震、火山噴發等自然因素,其中人為因素的化學和冶金行業是環境中重金屬的最主要來源(Navarro等,2008;陳炳睿等,2012;Vaalgamaa和Conley,2008)。滇東北會澤縣是中國有名的鉛鋅礦冶煉區,長期的土法冶煉對環境造成了嚴重的污染,導致了Cd、Pb、Zn等重金屬在土壤、水體和大氣中的高度積累,造成土壤重金屬污染,植被破壞,影響了整個流域的生態環境(房輝和曹敏,2009;陸泗進等,2014)。重金屬在土壤中積累,在農作物中殘留,隨食物鏈轉移,最終在動物、人體內積累,對人類身體健康造成危害,因此對重金屬污染土壤的修復迫在眉睫(雷冬梅等,2007)。
植物修復是指將具有一定重金屬富集能力的植物種植于污染土地上,生長一定時間后將植物地上部或整株收獲并集中填埋或焚燒,使土壤重金屬含量降低到可接受的水平(黑亮等,2007;李凱榮等,2013)。與土壤污染的傳統物理、化學修復治理技術相比,植物修復方式具有可在污染現場進行、修復成本低、對土壤擾動性小、不造成二次污染等優點,是土壤重金屬污染修復的最佳方法(鮑桐等,2008;胡亞虎等,2010;胡鵬杰等,2014)。但是,用于植物修復的超富集植物,生物量小,生長緩慢,吸收重金屬緩慢,使得修復時間漫長,效率低,效果不理想,限制了植物修復的推廣。
間作措施在中國農業生產中的應用歷史悠久。將農作物與超富集植物進行間作,可以實現“邊生產邊修復”的理念。利用間作種植,形成一套高效修復土壤重金屬污染的同時,實現在污染土壤上作物安全生產的種植模式(蔣成愛等,2009;衛澤斌等,2010)。本研究采用本土超累積植物續斷菊(sonchus asper L.Hill)與玉米(Zea mays L.)間作,期望能夠去除農用地中含量過高的Cd、Pb等重金屬,同時使農產品玉米的重金屬含量下降,為超累積植物與農作物間作條件下污染農用地的植物修復系統提供科學依據。
1 材料與方法
1.1 供試材料及實驗方法
選取玉米(會單4號)、續斷菊(本土植物)為試驗材料,于2014年4月至2014年9月進行試驗。
試驗地位于云南省會澤縣者海鎮瑪色卡村,海拔2100 m,年平均氣溫12.6 ℃,年降水量840 mm。土壤背景值:有機質質量分數為16.1 g·kg-1,全磷15.8 g·kg-1,速效磷 28.2 mg·kg-1,速效鉀 122 mg·kg-1;CEC 28.2 cmol·kg-1;鎘質量分數 25 mg·kg-1,鉛質量分數600 mg·kg-1;pH值5.5。
種植模式分為玉米與續斷菊間作、玉米單作和續斷菊單作 3個模式。單作續斷菊株行距均是 10 cm;單作玉米株距是20 cm,行距是30 cm。間作模式為1行玉米,2行續斷菊,玉米株距是20 cm,續斷菊株行距均是 10 cm,玉米和續斷菊行距 10 cm。每個種植模式重復3次,小區間筑梗,隨機排列,共9個小區,每個小區面積為15 m2(3 m×5 m)。續斷菊種子采自會澤 Pb/Zn礦區,播種前種子用10%的H2O2消毒30 min,使用烤煙型基質和漂盤育苗,待苗長到5~6 cm,選擇長勢良好,大小均一的幼苗在玉米播種時(4月15日)進行移栽。玉米播種期是2014年4月15日,拔節期是6月15日,抽穗期是7月15日,成熟期是8月15日。
1.2 采樣方法及預處理
在玉米拔節期(6月15日),抽穗期(7月15日),成熟期(8月15日)均取樣,每次單作小區取3株玉米或續斷菊及根際土壤,間作小區取3株玉米和對應3株續斷菊及根際土壤,帶回實驗室處理。
將續斷菊分成地上和地下2部分,玉米分成根、莖、葉、果實(成熟期)4部分,分別用自來水沖洗后,再用去離子水沖洗干凈,晾干后于105 ℃殺青30 min,然后70 ℃烘干至恒質量,分別測定干物質量。烘干樣品用粉碎機全部粉碎、混勻,過0.25 mm篩后裝袋備用。
土壤樣品避光自然風干,充分攪拌均勻后,過0.25 mm篩備用。
1.3 指標測定及計算方法
土壤樣品和植物樣品分別用濃硝酸-濃鹽酸-高氯酸、硝酸-高氯酸消煮后測定Cd、Pb含量,Cd、Pb含量的測定分別采用石墨爐-原子吸收光譜法、火焰-原子吸收光譜法。
Cd、Pb累積特征用富集系數和轉移因子表示,其計算公式分別為:
續斷菊富集系數=植物地上部重金屬含量/土壤重金屬含量(李元等,2008);
轉運系數=植物地上部含量/根部含量(秦麗等,2013);
有效轉運系數=植物地上部含量×地上部生物量/(根部含量×根部生物量)(秦麗等,2013);
收獲指數=籽粒質量/總質量(夏海勇等,2013)。
1.4 數據處理方法
數據采用Excel進行常規分析,土壤與植物之間、植物地上部與根之間重金屬含量的差異性、相關性采用SPSS軟件分析。
2 結果與分析
2.1 不同生育時期續斷菊與玉米生物量
拔節期和抽穗期,續斷菊生物量均是地上部大于根部,且間作續斷菊地上部大于單作續斷菊地上部(表1)。成熟期,間作續斷菊地上部干質量(生物量)顯著大于單作續斷菊地上部干質量。間作后,續斷菊生物量相比單作增加,3個時期分別增加了1.35%、14.29%、31.55%。
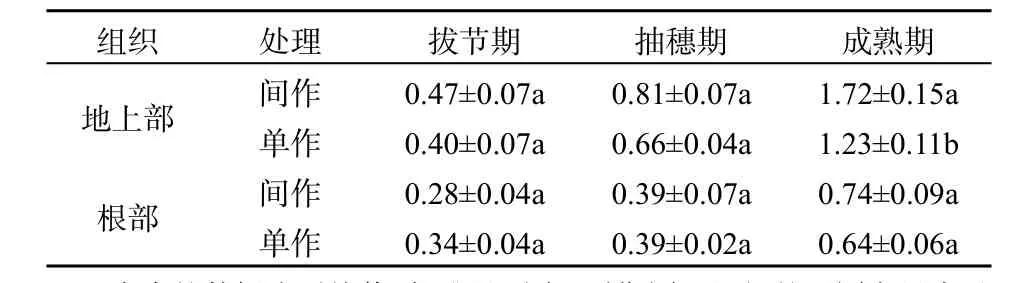
表1 續斷菊不同生育期生物量Table 1 Biomass of S. asper in different growth periods g·plant-1
在拔節期、抽穗期和成熟期,無論是間作還是單作,玉米生物量均為:莖>葉>根(表2)。間作玉米的總生物量(根+莖+葉)大于單作,玉米的根、莖、葉各部分生物量均為:間作>單作。間作玉米和單作玉米均是籽粒干質量最大,且間作玉米籽粒干重顯著大于單作玉米籽粒干重(表2)。總體來看,間作玉米生物量相比單作提高了29.02%。
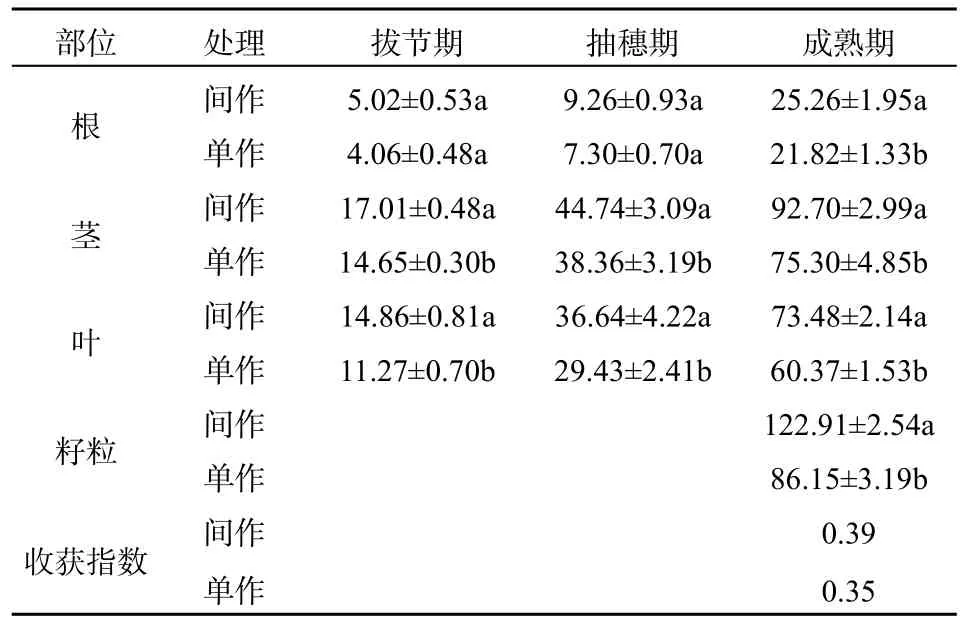
表2 玉米不同生育期生物量Table 2 Biomass of Z. Mays in different growth period g·plant-1
2.2 不同生育期玉米不同部位Cd、Pb質量分數
玉米各部位Cd質量分數由拔節期向成熟期呈遞減規律,間作玉米成熟期根、莖、葉Cd質量分數相對于拔節期分別降低了 24.51%、29.06%、55.32%,單作玉米成熟期根、莖葉Cd質量分數相對于拔節期分別降低了22.05%、7.20%、45.02%,同一部位,間作Cd質量分數下降大于單作(圖1)。

圖1 玉米不同部位不同時期Cd質量分數Fig. 1 Cd contents of different parts of Z. mays in different growth periods
拔節期,單作與間作玉米Cd質量分數分布均為:根>葉>莖,間作玉米根、莖、葉之間沒有差異性,而單作玉米的根部Cd質量分數顯著大于莖、葉部分,且間作玉米根、莖葉平均 Cd質量分數(13.97 mg·kg-1)小于單作平均Cd質量分數(14.54 mg·kg-1);抽穗期,間作玉米Cd質量分數分布為:根>葉>莖,單作分布為:根>莖>葉,均是根部 Cd質量分數顯著大于莖葉部分,且間作玉米根、莖、葉平均Cd質量分數(10.41 mg·kg-1)小于單作平均Cd質量分數(11.90 mg·kg-1);成熟期,單作與間作玉米根、莖、葉、籽粒Cd質量分數差異顯著,大小順序為:根>莖>籽粒>葉,且間作玉米根、莖、葉、籽粒平均Cd質量分數(8.74 mg·kg-1)小于單作平均Cd質量分數(10.94 mg·kg-1)。
拔節期到抽穗期,玉米各部位 Pb質量分數均呈現增加趨勢,抽穗期到成熟期,Pb質量分數呈現減少趨勢,且成熟期根莖 Pb質量分數均小于拔節期,葉則大于拔節期(圖2)。
拔節期與抽穗期,單作與間作玉米 Pb質量分數分布為:根>莖>葉。拔節期,單作與間作玉米各部位 Pb質量分數差異顯著,且間作玉米根莖葉平均Pb質量分數(52.10 mg·kg-1)小于單作Pb質量分數(56.42 mg·kg-1);抽穗期,間作玉米各部位Pb質量分數差異顯著,單作玉米根部 Pb質量分數顯著大于莖、葉,且間作玉米根、莖、葉平均 Pb質量分數(65.30 mg·kg-1)小于單作Pb質量分數(71.34 mg·kg-1);成熟期,間作玉米Pb質量分數分布為:根>葉>莖>籽粒,單作玉米Pb質量分數分布為:根>籽粒>葉>莖,均是根部顯著大于其他器官,且間作玉米根、莖、葉平均Pb質量分數(44.90 mg·kg-1)小于單作Pb質量分數(51.97 mg·kg-1)。
2.3 不同生育期續斷菊不同部位Cd、Pb質量分數及Cd、Pb累積特征
間作與單作續斷菊根部、地上部Cd質量分數從拔節期到成熟期呈現增加趨勢,間作續斷菊根部與地上部 Cd質量分數分別增加 16.88、15.45 mg·kg-1,單作續斷菊根部與地上部Cd質量分數分別增加5.5、10.09 mg·kg-1,間作根部、地上部Cd質量分數增加量大于單作(圖3)。在拔節期、成熟期,單作與間作續斷菊Cd質量分數分布均是地上部顯著大于根部。
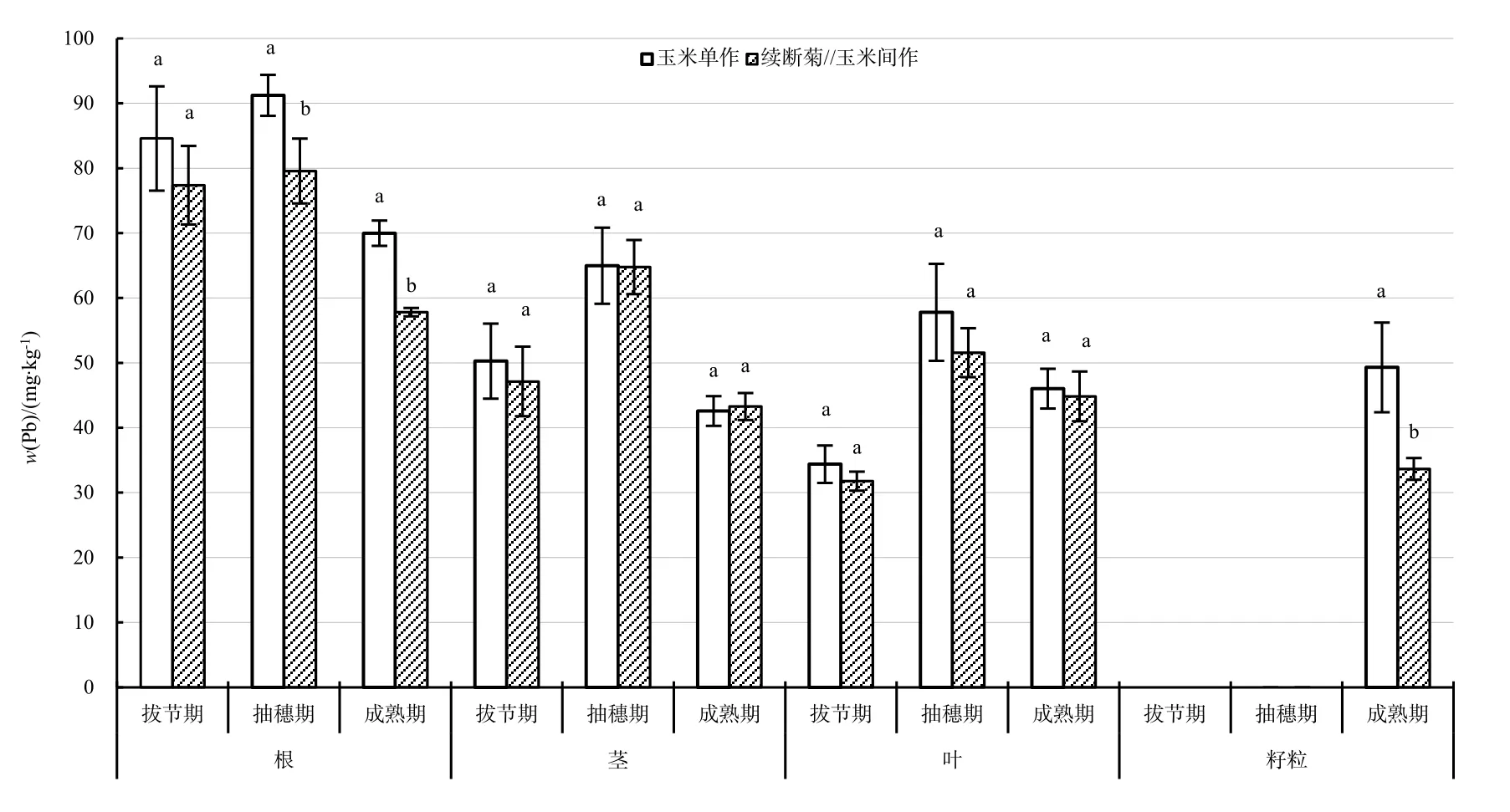
圖2 玉米不同部位不同時期Pb質量分數Fig. 2 Different parts Pb contents of Zea mays L. in different periods
間作、單作續斷菊地上部 Pb質量分數在不同生育期沒有顯著差異,根部 Pb質量分數從抽穗期到成熟期呈增加趨勢,單作續斷菊根部 Pb質量分數增加量(112.77 mg·kg-1)大于間作增加量(54.99 mg·kg-1)。整個生育期,單作與間作續斷菊均是根部Pb質量分數顯著大于地上部(圖4)。
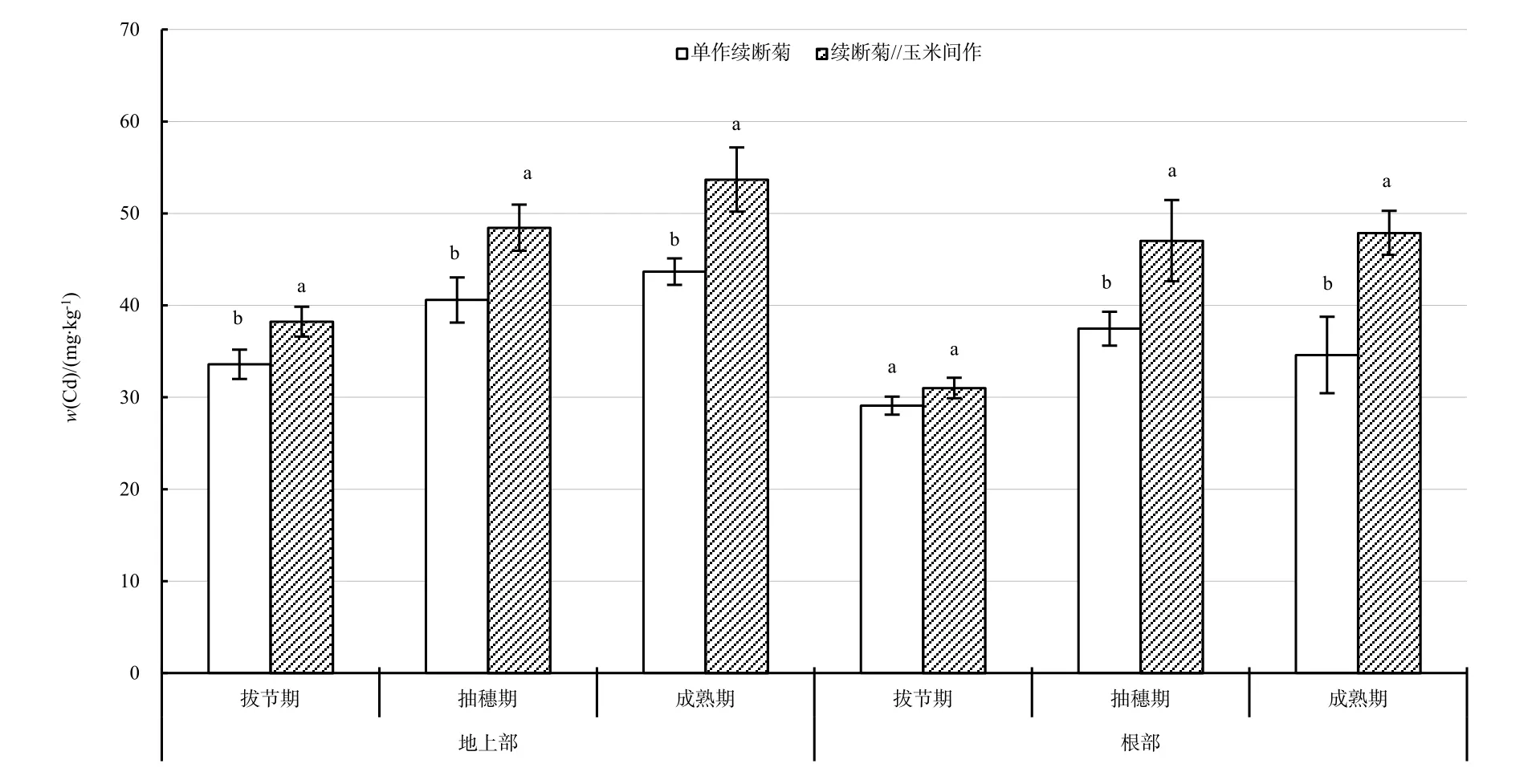
圖3 不同時期續斷菊Cd質量分數Fig. 3 Cd contents of S. asper. in different growth periods
3個時期,單作與間作續斷菊Cd轉運系數均大于1,間作續斷菊Cd平均轉運系數1.13,單作續斷菊Cd平均轉運系數1.16,轉運系數在拔節期為間作大于單作,抽穗期與成熟期均為間作小于單作;間作與單作續斷菊有效轉運系數與富集系數均大于1,且隨著時間的推移逐漸增加,相比于拔節期,成熟期的間作續斷菊有效轉運系數與富集系數分別增加了 26.09%、89.74%,單作分別增加了78.65%、69.23%,3個時期有效轉運系數與富集系數均為間作大于單作。
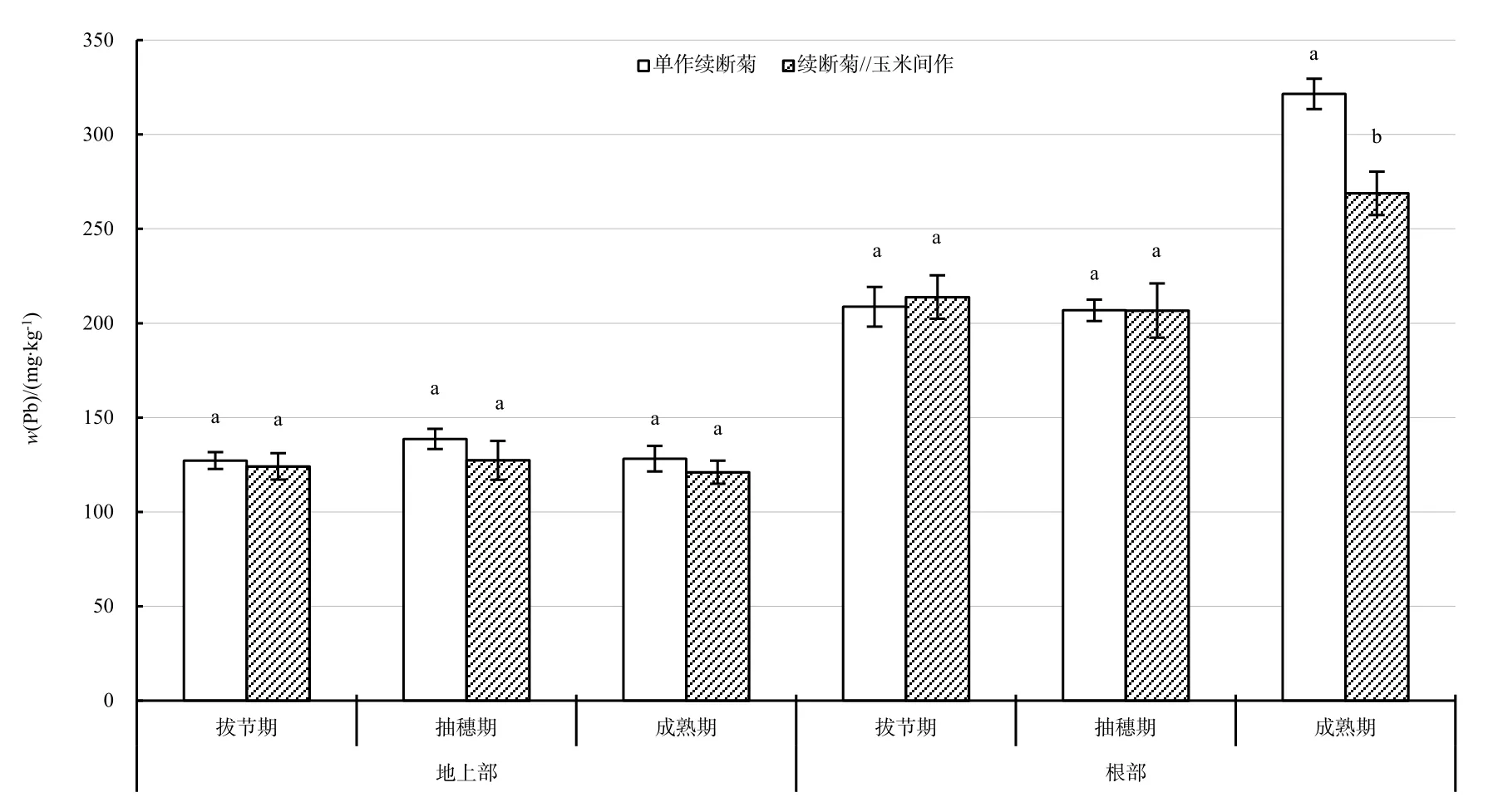
圖4 不同時期續斷菊Pb質量分數Fig. 4 Pb contents of S. asper in different growth periods
3個時期,間作與單作續斷菊Pb轉運系數、富集系數均小于1(表3),間作Pb平均轉運系數與平均富集系數分別為0.55、0.24,單作分別為0.56、0.25。續斷菊由拔節期到成熟期,間作續斷菊轉運系數降低了22.41%,單作降低了34.43%,但有效轉運系數間作增加8.25%,單作增加6.94%。
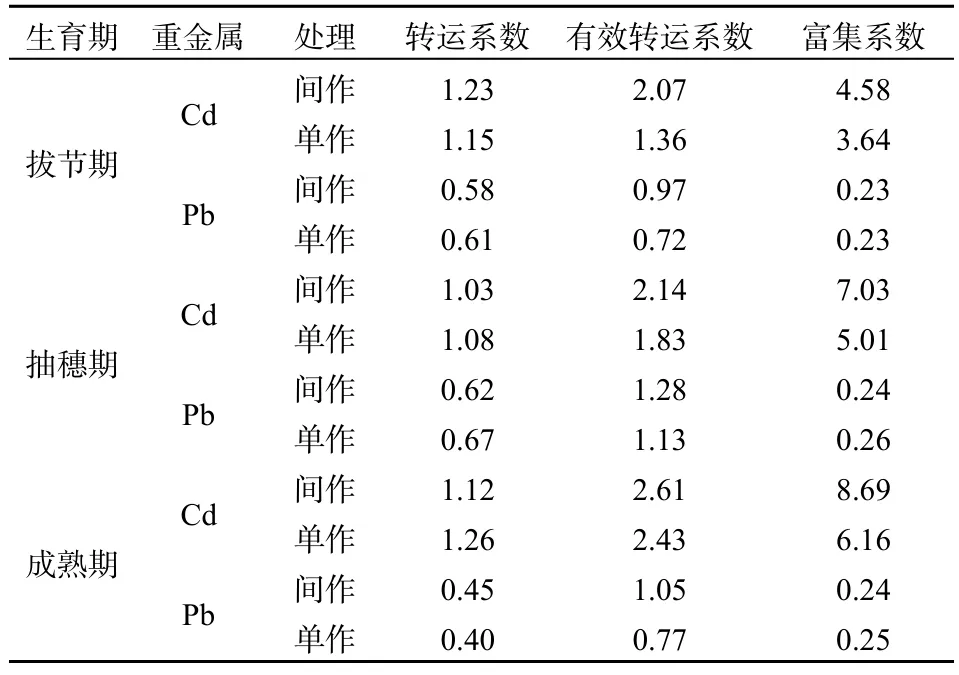
表3 續斷菊轉運系數與富集系數Table 3 The enrichment coefficient and transport coefficients of S. asper
2.4 不同生育期土壤Cd、Pb質量分數
在整個生育期,Cd、Pb質量分數在土壤中均是單作玉米>單作續斷菊>間作。間作、單作種植模式下土壤Cd、Pb質量分數均有下降,土壤Cd質量分數下降幅度大小為:間作>單作續斷菊>單作玉米。土壤 Pb質量分數下降幅度大小為:單作續斷菊>單作玉米>間作。間作模式能顯著降低土壤Cd、Pb質量分數,從拔節期到成熟期土壤Cd質量分數下降25.99%,土壤Pb質量分數下降6.78%;單作續斷菊模式同樣有利于土壤Cd、Pb質量分數的降低,從拔節期到成熟期土壤 Cd質量分數下降23.19%,土壤 Pb質量分數下降7.53%;單作玉米模式對土壤Cd、Pb質量分數有同樣的降低效果,從拔節期到成熟期土壤Cd下降14.78%,下降效果沒有間作、單作續斷菊兩種種植模式明顯,土壤Pb質量分數下降7.23%,下降幅度與間作、單作玉米模式無明顯差別(圖5、圖6)。
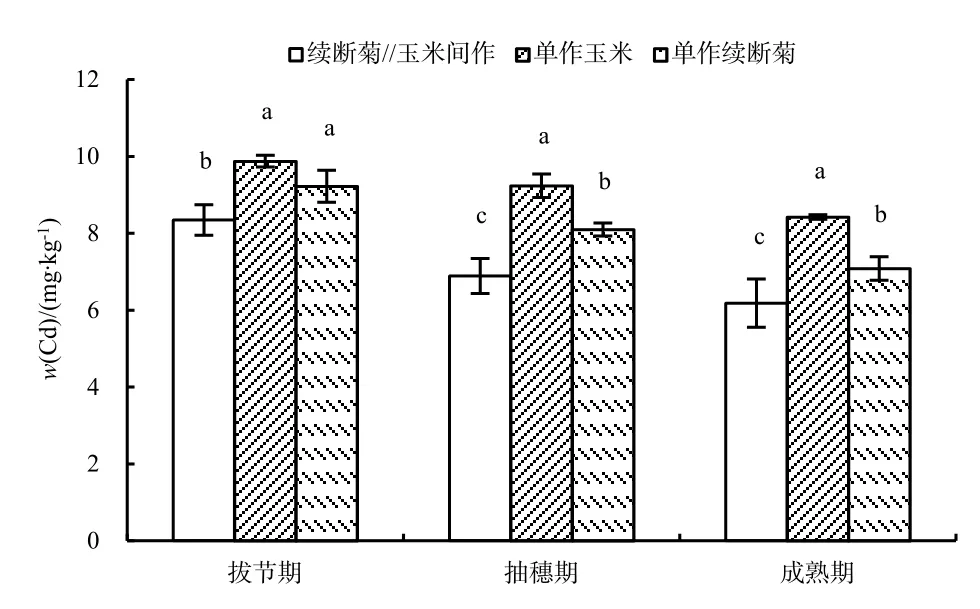
圖5 不同時期土壤Cd質量分數Fig. 5 Cd contents of soil in different growth periods
3 討論
本試驗表明,在拔節期與抽穗期,間作對續斷菊和玉米根部生物量沒有影響,但能顯著提高間作玉米莖部和葉部生物量;成熟期,間作在顯著提高續斷菊地上部生物量的同時,也顯著提高了間作玉米生物量。蔣成愛等(2009)研究表明玉米和東南景天間作,顯著提高了富集植物東南景天的生物量。間作條件下的玉米生物量與續斷菊生物量比單作時顯著增加的原因可能是間作條件下續斷菊與玉米互相促進光合作用(焦念元等,2013a,2013b)、增加了土壤養分、維持土壤肥力、提高養分利用效率(呂越等,2014;蘇本營等 2013)。因此,在對輕中度重金屬污染的土壤進行植物修復時,可以通過超積累植物與農作物的間作促進超積累植物生物量,加強對污染土壤的修復,同時實現在污染土壤上農作物的安全生產。
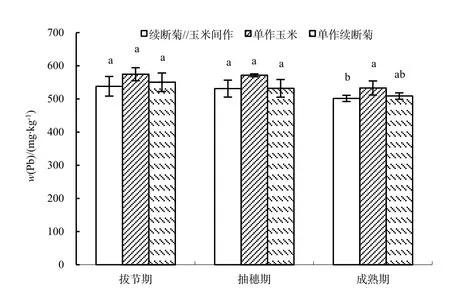
圖6 不同時期土壤Pb質量分數Fig. 6 Pb contents of soil in different growth periods
間作條件下能夠顯著增加了玉米的生物量(秦歡等,2012),本實驗研究表明,續斷菊與玉米間作后,間作玉米籽粒產量高于單作玉米籽粒產量,間作玉米籽粒產量的收獲指數(0.39)與單作玉米籽粒產量的收獲指數(0.35)出現差異性,間作在提高籽粒產量的同時,卻降低了玉米籽粒中Cd的濃度。可能的原因是間作時提高了玉米籽粒產量,導致玉米籽粒Cd元素濃度的“稀釋效應”,有研究表明氮肥施用導致較高的玉米籽粒產量,使玉米籽粒中重金屬Zn元素濃度產生“稀釋效應”(Feil等,2005;夏海勇等,2013),間作后玉米籽粒鉻的濃度會降低(An等,2011)。另一可能的原因是間作玉米Cd從營養器官向籽粒的轉移或再活化速率受到抑制,從而其質量分數低于單作玉米,有研究表明在適宜條件和施用Zn肥情況下,高N肥施用能夠促進營養器官對 Fe和 Zn的吸收和再活化(Kutman等,2011)。夏海勇等(2013)研究表明玉米與蠶豆、鷹嘴豆和大豆間作,使間作玉米籽粒Fe、Mn、Cu和 Zn濃度降低,是由 Fe、Mn、Cu和Zn從營養器官向籽粒轉移或再活化的比例降低所致。在間作體系中,通過根系分泌物對根際環境的pH、Eh等物理、化學性質,以及土壤酶活性、根際周圍土壤中微生物種群分布等對土壤重金屬的有效性以及植物對重金屬的吸收產生重要影響(陳英旭等,2000;梁開明等,2014;常學秀等,2000)。所以導致間作后玉米Cd質量分數低于單作的可能原因是續斷菊與玉米間作后,通過根系分泌有機酸與Cd進行螯合作用,降低土壤pH,提高土壤 Cd的活化效率(土壤有效態鎘/土壤全鎘),而續斷菊是Cd的超積累植物,對Cd具有強吸收作用(秦麗等,2013)。從本實驗結果來看,雖然成熟期間作續斷菊轉運系數小于單作,但是間作續斷菊根部、地上部Cd質量分數大于單作續斷菊,且間作有效轉運系數大于單作,說明間作后續斷菊的轉運能力強于單作。衛澤斌等(2005)研究表明,間作方式可以提高植物對重金屬的提取效率。將重金屬富集植物和農作物間作對污染土壤進行植物修復,富集植物能抑制農作物對重金屬的吸收量,同時能提高富集植物對重金屬的提取效率。玉米與續斷菊在間作體系下,既能提高玉米產量,又能增強超積累植物對Cd的吸收,是一種“雙贏”模式。
在本研究中,不同時期下玉米、續斷菊對Cd、Pb的積累能力不同,與任立民等(2006)研究發現并總結出同一植物吸收積累不同重金屬元素的能力是不同的結論一致,而且同一植物不同器官對同一種重金屬元素的吸收富集能力也有很大的差異(陳建軍等,2014)。Cd質量分數在玉米植株不同器官分布依次為根部>莖部>籽粒>葉部,且間作、單作玉米根部Cd質量分數與莖、葉籽粒出現顯著差異,說明根部更容易累積Cd。根中含量較高可能與Cd進入根的皮層細胞后和根內蛋白質、多糖類、核酸等化合成為穩定的大分子絡合物或不穩定性有機大分子而沉積有關(張旭紅等,2008)。重金屬在植物體內的分布規律是在新陳代謝旺盛的器官(如根部)蓄積量較大,而營養儲存器官(如莖部、葉片、果實、籽粒)蓄積量則較少(關共湊等,2006)。而隨時間的推移,間作與單作玉米根、莖、葉部Pb積累量先上升后下降,在成熟期根莖積累量均下降到最小,原因可能是玉米根部在拔節期生長旺盛,對Pb大量吸收,因為根部積累較多Pb,根部的代謝加強,導致 Pb從根部不斷向莖葉部分遷移,同時根部化學性狀的改變以及根系分泌物等作用抑制了根部的吸收機能,使根部從土壤中吸收的 Pb逐漸減少(關共湊等,2006;曠遠文等,2003;徐衛紅等,2006;吳林坤等,2014)。因此,隨著時間的推移,在根部積累的Pb就越來越少。
4 結論
Cd在玉米植株不同部位的積累隨時間的變化是:間作與單作玉米根、莖、葉部積累量均在拔節期最大,隨著時間的推移,其積累的重金屬越來越少。Pb在玉米植株不同部位的積累隨時間的變化是:間作與單作玉米根、莖、葉部積累量先上升后下降,根莖積累量均下降到最小,葉積累量抽穗期和成熟期都大于拔節期。
Cd、Pb在土壤中被玉米與續斷菊吸收以后,對于玉米,主要集中在根部;對于續斷菊地上部主要集中Cd,根部主要積累Pb。Cd質量分數在玉米植株不同器官分布依次為根部>莖部>籽粒>葉部;在續斷菊中分布為地上部>根部,且在間作體系下,能顯著減少玉米根莖葉和籽粒Cd、Pb質量分數,能增加續斷菊對Cd的積累量。玉米根、莖、葉、籽粒中重金屬元素積累量分布是:w(Pb)> w(Cd)。
AN L Y, PAN Y H, WANG Z B, et al. 2011. Heavy metal absorption status of five plant species in monoculture and intercropping [J]. Plant Soil, 345(1-2): 237-245.
FEIL B, MOSER S B, JAMPATONG S, et al. 2005. Mineral composition of the grains of tropical maize varieties as affected by pre-anthesis drought and rate of nitrogen fertilization [J]. Crop Science, 45(2): 516-523.
KUTMAN U, YILDIZ B, CAKMAK I. 2011. Effect of nitrogen on uptake, remobilization and partitioning of zinc and iron throughout the development of durum wheat [J]. Plant Soil, 342(1-2): 149-164.
NAVARRO M C, PEREZ-SIRVENT C, MARTINEZ-SANCHEZ M J, et al. 2008. Abandoned mine sites as a source of contamination by heavy metals: A case study in a semi-arid zone [J]. Journal of Geochemical Exploration, 96(2-3): 183-193.
VAALGAMAA S, CONLEY D J. 2008. Detecting environmental change in estuaries: Nutrient and heavy metal distributions in sediment cores in estuaries from the Gulf of Finland, Baltic Sea [J]. Estuarine, Coastal and Shelf Science, 76(1): 45-56.
鮑桐, 廉梅花, 孔麗娜, 等. 2008. 重金屬污染土壤植物修復研究進展[J].生態環境學報, 17(2): 858-865.
常學秀, 段昌群, 王煥校. 2000. 根分泌作用與植物對金屬毒害的抗性[J]. 應用生態學報, 11(2): 315-320.
陳炳睿, 徐超, 呂高明, 等. 2012. 6種固化劑對土壤Pb、Cd、Cu、Zn的固化效果[J]. 農業環境科學學報, 31(7): 1330-1336.
陳建軍, 于蔚, 祖艷群, 等. 2014. 玉米(Zea mays)對鎘積累與轉運的品種差異研究[J]. 生態環境學報, 23(10): 1671-1676.
陳英旭, 林琦, 陸芳, 等. 2000. 有機酸對鉛、鎘植株危害的解毒作用研究[J]. 環境科學學報, 20(4): 467-472.
房輝, 曹敏. 2009. 云南會澤廢棄鉛鋅礦重金屬污染評價[J]. 生態學雜志, 28(7): 1277-1273.
關共湊, 徐頌, 黃金國. 2006. 重金屬在土壤-水稻體系中的分布、變化及遷移規律分析[J]. 生態環境學報, 15(2): 315-318.
黑亮, 吳啟堂, 龍新憲, 等. 2007. 東南景天和玉米套種對 Zn污染污泥的處理效應[J]. 環境科學, 28(4): 4852-4858.
胡鵬杰, 李柱, 鐘道旭, 等. 2014. 我國土壤重金屬污染植物吸取修復進展[J]. 植物生理學報, 50(5): 577-584.
胡亞虎, 魏樹和, 周啟星, 等. 2010. 螯合劑在重金屬污染土壤植物修復中的應用研究進展[J]. 農業環境科學學報, 29(11): 2055-2063.
蔣成愛, 吳啟堂, 吳順輝, 等. 2009. 東南景天與不同植物混作對土壤重金屬吸收的影響[J]. 中國環境科學, 29(9): 985-990.
焦念元, 寧堂原, 楊萌珂, 等. 2013a. 玉米花生間作對玉米光合特性及產量形成的影響[J]. 生態學報, 33(14): 4324-4330.
焦念元, 楊萌珂, 寧堂原, 等. 2013b. 玉米花生間作和磷肥對間作花生光合特性及產量的影響[J]. 植物生態學報, 37(11): 1010-1017.
曠遠文, 溫達志, 鐘傳文, 等. 2003. 根系分泌物及其在植物修復中的作用[J]. 植物生態學報, 27(5): 709-717.
雷冬梅, 段昌群, 王明. 2007. 云南不同礦區廢棄地土壤肥力與重金屬污染評價[J]. 農業環境科學學報, 26(2): 612-616.
李凱榮, 閆寶環, 時亞坤. 2013. 黃土高原南緣煤矸石廢棄地不同植物對重金屬污染的耐受性研究[J]. 草地學報, 21(6): 1093-1100.
李元, 方其仙, 祖艷群. 2008. 2種生態型續斷菊對Cd的累積特征研究[J].西北植物學報, 28(6): 1150-1154.
梁開明, 傅玲, 章家恩, 等. 2014. 水稻/再力花單、間作系統中作物對鎘脅迫的葉綠素熒光響應及鎘積累特征[J]. 華南農業大學學報, 35(4): 35-41.
陸泗進, 王業耀, 何立環. 2014. 會澤某鉛鋅礦周邊農田土壤重金屬生態風險評價[J]. 生態環境學報, 23(11): 1832-1838.
呂越, 吳普特, 陳小莉, 等. 2014. 玉米-大豆間作系統的作物資源競爭[J]. 應用生態學報, 25(1): 139-146.
秦歡, 何忠俊, 熊俊芬, 等. 2012. 間作對不同品種玉米和大葉井口邊草吸收積累重金屬的影響[J]. 農業環境科學學報, 31(7): 1281-1288.
秦麗, 祖艷群, 湛方棟, 等. 2013. 續斷菊與玉米間作對作物吸收積累鎘的影響[J]. 農業環境科學學報, 32(3): 471-477.
任立民, 劉鵬, 鄭啟恩, 等. 2006. 廣西大新縣錳礦區植物重金屬污染的調查研究[J]. 亞熱帶植物科學, 35(3): 5-8.
蘇本營, 陳圣賓, 李永庚, 等. 2013. 間套作種植提升農田生態系統服務功能[J]. 生態學報, 33(14): 4505-4514.
衛澤斌, 郭曉方, 丘錦榮, 等. 2010. 間套作體系在污染土壤修復中的應用研究進展[J]. 農業環境科學學報, 29(增刊): 267-272.
衛澤斌, 吳啟堂, 龍新憲, 等. 2005. 利用套種和混合添加劑修復重金屬污染土壤[J]. 農業環境科學學報, 24(6): 1262-1263.
吳林坤, 林向民, 林文雄. 2014. 根系分泌物介導下植物-土壤-微生物互作關系研究進展與展望[J]. 植物生態學報, 38(3): 298-310.
夏海勇, 趙建華, 孫建好, 等. 2013. 油菜、蠶豆、鷹嘴豆和大豆對間作玉米籽粒Fe, Mn, Cu和Zn濃度及地上部累積量的影響[J]. 中國科學, 43(7): 557-568.
徐衛紅, 黃河, 王愛華, 等. 2006. 根系分泌物對土壤重金屬活化及其機理研究進展[J]. 生態環境學報, 15(1): 184-189.
張旭紅, 高艷玲, 林愛軍, 等. 2008. 植物根系細胞壁在提高植物抵抗金屬離子毒性中的作用[J]. 生態毒理學報, 3(01): 9-14.
Distribution Characteristics of Pb and Cd in Different Parts of Sonchus asper and Zea mays in An Intercropping System
TAN Jianbo, CHEN Xing, GUO Xianhua, LI Yuan, ZU Yuanqun*
College of Resources and Environment, Yunnan Agricultural University, Kunming 650201, China
This experiment was to use the method of phytoremediation to repair the polluted farmland by Cd and Pb around lead-zinc mine area in Huize. Hyperaccumulator compositae (Sonchus asper L.Hill) and maize (Zea mays L.) were intercropped in polluted farmland, expecting to not only reduce the heavy metal pollution in soil but also obtain qualified agricultural products. By field plot experiments , plant biomass and heavy metal contents in different organs were determined and we studied the plants on Pb, Cd distribution characteristics in different organs under intercropping system of maize and compositae. The results indicated that: (1) The biomass of intercropping compositae was increased by 31.55% than the monoculture and the biomass of intercropping maize was increased by 29.02% than the monoculture. (2) From jointing stage to maturity stage, the mass fraction of Cd in all organs of maize was decreased by monoculture or intercropping. The Cd mass fraction in roots, stems and leaves by intercropping maize was decreased by 24.51%,29.06%,55.32%. The Cd mass fraction in roots, stems and leaves by monoculture maize was decreased by 22.05%,7.20%,45.02%. In different periods, the Cd contents of roots and leaves by intercropping maize were lower than the monoculture. The Cd content of roots showed significant difference at heading stage and maturity stage. The Cd content of leaves was significant difference at maturity stage. The Cd content of stems at heading stage and maturity stage in monoculture was higher than the intercropping and had significant difference at maturity stage. The Pb mass fraction in all organs of maize increased firstly, and then decreased and the Pb mass fraction in roots and stems at maturity stage was lower than the jointing stage and the Pb mass fraction of leaves at the maturity stage was higher than the jointing stage. In different periods, the Pb contents of roots and leaves by intercropping maize were lower than the monoculture. The Pb content of roots at heading stage and maturity stage had significant difference. The Cd and Pb contents of maize grain were significantly lower than that of monoculture. (3) The Cd mass fraction of the compositae increased with the time passing and the Cd content of overground plant part by intercropping was significantly higher than the monoculture. At heading stage and maturity stage, the Cd content of roots by intercropping was significantly higher than the monoculture. The Cd mass fraction of roots and overground plant part by intercropping increased by 16.88, 15.45 mg·kg-1, respectively. The Cd mass fraction of roots and overground part under monoculture increased by 5.5, 10.09 mg·kg-1, respectively. (4) The Cd and Pb mass fraction in soil was decreased, both monoculture and intercropping. It decreased by 25.99% under intercropping. It decreased by 23.19% under monoculture compositae and 14.78% under monoculture maize. The Pb mass fraction in soil decreased by 6.78% under intercropping, and 7.53% under monoculture compositae and 7.23% under monoculture maize. The results showed that compositae and maize intercropping systems not only promoted Cd accumulation of all organs of compositae but also inhibited Cd accumulation of all organs of maize.
Maize; Sonchus asper L. Hill; intercropping; Cd; Pb; soil remediation
10.16258/j.cnki.1674-5906.2015.04.023
X173
A
1674-5906(2015)04-0700-08
譚建波,陳興,郭先華,李元,祖艷群. 續斷菊與玉米間作系統不同植物部位Cd、Pb分配特征[J]. 生態環境學報, 2015, 24(4): 700-707.
TAN Jianbo, CHEN Xing, GUO Xianhua, LI Yuan, ZU Yuanqun. Distribution Characteristics of Pb and Cd in Different Parts of Sonchus asper and Zea mays in An Intercropping System [J]. Ecology and Environmental Sciences, 2015, 24(4): 700-707.
國家自然科學-云南省聯合基金項目(U1202236)
譚建波(1990年生),男,碩士研究生,主要從事土壤重金屬污染與修復生態學研究。E-mail: 237958437@qq.com *通信作者:祖艷群(1966年生),女,教授,博士生導師。E-mail: 649332092@qq.com
2014-12-29
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
