Trichothecium roseum 蛋白激發子誘導杏果抗黑斑病的作用機理
2015-12-13 07:02:37余曉燕李永才沈科萍胡林剛唐亞梅
食品科學 2015年4期
余曉燕,畢 陽,李永才*,燕 璐,沈科萍,胡林剛,唐亞梅
(甘肅農業大學食品科學與工程學院,甘 肅 蘭州 730070)
Trichothecium roseum 蛋白激發子誘導杏果抗黑斑病的作用機理
余曉燕,畢 陽,李永才*,燕 璐,沈科萍,胡林剛,唐亞梅
(甘肅農業大學食品科學與工程學院,甘 肅 蘭州 730070)
研究非親和菌株粉紅單端孢(Trichothecium roseum)菌體蛋白激發子對杏果實抗黑斑病的誘導效果及其作用機理。以‘蘭州大接杏’為試材,通過處理后不同時間損傷接種杏果黑斑病菌(Alternaria alternata),研究T. roseum蛋白激發子對杏果實黑斑病抗病性的 誘導及其時效性,同時進一步研究最佳處理質量濃度對杏果實組織活性氧及抗性相關酶的影響。結果表明,T. roseum蛋白激發子處理可顯著抑制損傷接種杏果病斑的擴展(P<0.05),其中14.4 μg/mL蛋白激發子的控制效果最好,且誘導處理72 h時杏果產生最大抗性。T. roseum蛋白激發子處理后杏果組織O2—·的生成速率和H2O2含量分別在48 h和72 h出現明顯的峰值,同時處理的果實組織超氧化物歧化酶、過氧化物酶、多酚氧化酶、β-1,3-葡聚糖酶和幾丁質酶活性均有所增強。T. roseum菌體蛋白激發子處理能顯著增強杏果組織的抗性,其可能是通過激發活性氧爆發和提高杏果實抗病性相關酶的活性來實現的。可見T. roseum蛋白激發子可以作為一種有潛力的誘抗劑應用于采后病害的控制。
粉紅單端孢菌;蛋白激發子;杏果實;活性氧;誘導抗病性;黑斑病
杏(Prunus armeniaca L.)原產我國,甘肅省為全國杏的主要產區,其中‘蘭州大接杏’以個大、色艷、氣香、味美、質優而久負盛名[1-2]。由于杏的成熟期集中,貨架期較短,常溫條件下的采后壽命只有3~4 d,低溫條件下也不足3周。此外,杏的采后 腐爛也頗為嚴重,其中由互隔交鏈孢(Alternaria alternata)引起的黑斑病是低溫條件下杏的主要采后病害,該病原菌具有潛伏侵染的特性,對杏在低溫冷鏈物流期間的腐爛率影響很大,常常導致嚴重的經濟損失[3-5]。采用化學殺菌劑可有效控制杏的黑斑病,但由于存在藥物殘留、環境污染、以 及病原物產生抗藥性等問題而逐漸受到限制[6]。因此,開發新型安全可持續的采后病害防腐技術便顯得十分必要。生物農藥在綠色食品生產發展和環境保護中發揮著積極作用,其中微生物源激發子的開發與應用顯現出巨大的應用潛力[7]。該類激發子按化學性質分為蛋白類、寡糖類、肽類、糖蛋白類以及脂類等,其中蛋白類激發子又分為過敏蛋白、隱地蛋白、激活蛋白及其他蛋白,這些激發子均具有廣譜提高植物抗性、有效控制病害 的作用[8]。例如,用從極細鏈格孢菌(A. tenuissima)中獲得的蛋白激發子(Hrip1)處理煙草,能夠激活水楊酸途徑的蛋白激酶并且提高相關防衛基因的表達[9];將Hrip1基因轉入擬南芥,植株對灰霉病的抗性顯著增強[10];將稻瘟病菌(Magnaporthe grisea)的蛋白激發子PemG1基因轉入水稻可提高植株對稻瘟病的抗性[11];將梨火疫病菌(Erwinia amylovora)產生的過敏蛋白Harpin基因導入馬鈴薯可提高對晚疫病的抗性[8],用Harpin處理蘋果和甜瓜可有效提高果實的抗病性[12-13]。雖然蛋白類激發子在控制植物病害中的作用已有一些研究,但 多集中于田間病害,尚未見T. roseum源蛋白類激發子在果實上的應用報道。本實驗擬以‘蘭州大接杏’為試材,研究T. roseum菌體蛋白激發子處理對Alternaria alternata挑戰接種杏果黑斑病的控制效果;探索處理后果實O2—·產生速率和H2O2含量的積累,以及主要抗氧化酶活性的變化;分析處理對果實病程相關蛋白的影響,以期為真菌源激發子在果蔬采后病害控制中的進一步應用提供理論依據。
1 材料與方法
1.1 材料與儀器
供試‘蘭州大接杏’(Armeniaca vulgaris cv. Lam.)2013年7月采自甘肅省臨洮縣商業化杏園,采收后剔除傷、病果,選擇大小、果色均勻、八成熟的杏果實,5 h運抵甘肅農業大學食品科學與工程學院采后生物學與技術實驗室,0 ℃條件下貯藏待用。粉紅單端孢(T. roseum)由本實驗室提供;互隔交鏈孢(A. alternata)分離于自然發病的杏果實,經純化鑒定后馬鈴薯葡萄糖瓊脂培養基上保存待用。
SW-CJ-2FD潔凈工作臺 蘇凈集團蘇州安泰空氣技術有限公司;LDZX-30KBS立式壓力蒸汽滅菌鍋 上海申安醫療器械廠;DHP-9272B型恒溫培養箱 上海一恒科技有限公司。
1.2 方法
1.2.1 T. roseum菌體粗蛋白激發子的制備
參照Zhao Mingzhi等[14]方法稍有改動,從低溫保存的種管挑取少量菌絲體,轉接到固體馬鈴薯葡萄糖瓊脂培養基上,培養3~5 d后制備孢子懸浮液,按照1.5%(V/V)接種量接種到液體培養基,150 r/min、28℃振蕩培養,7 d后真空抽濾用蒸餾水洗滌多次的菌體并獲得抽干菌體,—80 ℃保存備用。取菌絲體10 g置于液氮研磨成粉末,迅速加入40 mL蛋白提取液(20 mmo1/L Tris-HCl緩沖液pH 7.0,2 mmo1/L乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA))振蕩混勻并提取1 h,沸水浴30 min后迅速冷卻,于4 ℃、12 000 r/min離心30 min,取上清液用0.45 μm濾膜過濾,即為粗蛋白激發子。
1.2.2 孢子懸浮液配制
參照周德慶[15]方法。取28 ℃條件下培養7 d的A. alternata,加入含0.05% Tween 20的無菌水20 mL,用涂布棒刮下平板上的病原菌孢子,然后轉入50 mL三角瓶中,在WYX-A微型旋渦混合器上振蕩15 s,再用雙層無菌紗布過濾,濾液用血球計數板計數算出孢子懸浮液的濃度后,最后稀釋至所需濃度(1×106孢子/mL)。
1.2.3 果實的處理
將果實分兩組,第1組分別用蒸餾水(CK)、144、14.4、1.44 μg/mL的粗蛋白激發子(以蛋白質量濃度計)噴灑處理,并晾干表面水分別裝箱于常溫,處理后12、24、36、48、60、72 h的杏果用滅菌打孔器(直徑3 mm)在果實陰陽面的赤道部位均勻刺孔2個(深約3 mm),待孔中果實汁液晾干后,分別接入20 μL A. alternata孢子懸浮液(1.0×106孢子/mL),晾干后將相同處理每3個裝入一個帶孔保鮮袋(厚0.01 mm),常溫(25±2) ℃貯藏,第3天觀察病斑并采用十字交叉法測量病斑直徑。每個處理15 個杏果,重復3 次。
第2組分別用蒸餾水和最佳處理質量濃度的粗蛋白激發子噴灑,晾干后裝箱置于2 ℃冷庫中。貯藏期間每隔12、24、48、72、96、120 h取3~5 mm果皮組織3 g(取樣前回溫1 h),用錫箔紙包好后立即用液氮進行冷凍,然后保存于—80 ℃待用。
1.2.4 O2—·產生速率和H2O2含量的測定
O2—·產生速率的測定參考王云飛等[16]方法略修改。稱取3 g果皮組織,加5 mL、pH 7.8、100 mmol/L磷酸緩沖液,在冰浴條件下研磨成漿,4 ℃、12 000 r/min離心15 min。取1 mL上清液,加入1 mL磷酸緩沖液和0.5 mL 10 mmol/L的鹽酸羥胺溶液,在25 ℃反應1 h。然后加入1 mL 17 mmol/L對氨基苯磺酸和1 mL 7 mmol/L α-萘胺,于25 ℃保溫30 min。然后530 nm波長處測定OD值。以nmol/(min·g)表示O2—·產生速率,平行3次。
H2O2含量的測定參照高雄杰[17]方法略修改。稱取3 g果皮組織,加入3 mL經4 ℃預冷的丙酮,冰浴條件下研磨成漿,4 ℃、12 000 r/min離心15 min。取1 mL上清液,加入100 μL 20%的四氯化鈦溶液和200 μL濃氨水,混勻反應5 min后離心10 min。沉淀部分用經4 ℃預冷的丙酮洗滌8 次,最后將沉淀溶于3 mL 1 mol/L H2SO4溶液中,于410 nm波長處測定OD值。以標準H2O2溶液制作標準曲線,H2O2含量以μmol/g表示,平行3 次。
1.2.5 超氧化物歧化酶(su peroxide dismutase,SOD)、過氧化物酶(peroxidase,POD)、多酚氧化酶(polyphenol oxidase,PPO)活性的測定
SOD活性的測定參考曹建康等[18]方法略有修改。稱取3 g果皮組織,加入5 mL經4℃預冷的100 mmol/L磷酸緩沖液,在冰浴中充分研磨,4 ℃、12 000 r/min離心20 min,上清液立即用于酶活性測定。于指形管中加入1.5 mL 50 mmol/L磷酸緩沖液,300 ?L 130 mmol/L甲硫氨酸,300 ?L 750 ?mol/L氮藍四唑,300 ?L 100 ?mol/L EDTA-Na2,100 ?L粗酶液,最后加入300 ?L 20 ?mol/L核黃素,2支對照管以緩沖液代替酶液,混勻后將1支對照管置于暗處,其他各管于4 000 lx日光燈下反應15 min。以不照光管做空白參比,于560 nm波長處分別測定各管的OD值。活性以含每分鐘每克果實組織的反應體系抑制氮藍四唑光化還原的50%為一個活力單位(U)計算見公式(1):
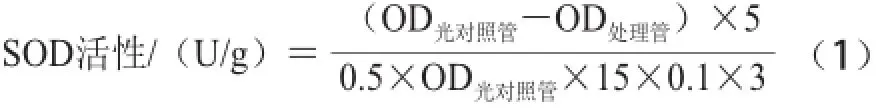
POD活性測定參照Bi Yang等[13]方法略修改。取3 g果皮組織,加入5 mL 50 mmol/L pH 5.9的磷酸緩沖液,在冰浴中充分研磨,4 ℃、12 000 r/min離心20 min,上清液用于POD活性測定。酶促反應體系為:1.8 mL 0.05 mol/L愈創木酚、100 μL粗酶液和100 μL 2% H2O2,以蒸餾水作對照,2 min內在470 nm波長處測定OD值的變化,以每克果蔬樣品每分鐘光密度值增加1時為1個POD活性單位,其表示為ΔOD470nm/(min·g)。
PPO活性的測定參照任亞琳等[19]方法略修改。稱3 g果皮組織,加入5 mL 50 mmol/L pH 5.5的磷酸緩沖液,在冰浴中充分研磨,4 ℃、12 000 r/min離心20 min,上清液用于PPO活性測定。酶促反應體系為:4 mL磷酸緩沖液、1 mL 50 mmoL鄰苯二酚、100 μL的酶提取液,以蒸餾水作參比,2 min內在420 nm波長處測定OD值的變化,以每克果蔬樣品每分鐘光密度值增加1為1個PPO酶活性單位,其表示為ΔOD420nm/(min·g)。
1.2.6 β-1,3-葡聚糖酶(β-1,3-glucanase,GLU)和幾丁質酶(chitinas,CHT)活性的測定
CHT活性的測定參考曹建康等[18]方法并修改,稱3 g果皮組織,加5 mL的29 mg EDTA、35 μL β-巰基乙醇、pH 5.2、50 mmol/L乙酸-乙酸鈉緩沖液,在冰浴中充分研磨,4℃、12 000 r/min離心30 min,取上清液,與5 倍體積的預冷丙酮混合,置于—20 ℃沉淀蛋白質,離心收集沉淀,最后用氮氣吹干,將其溶解于提取緩沖液中,同樣條件下離心收集上清,反應酶體系為:0.5 mL 50 mmol/L乙酸-乙酸鈉緩沖液、0.5 mL 10 g/L膠狀幾丁質懸浮液、0.5 mL的酶提取液,以滅活的酶液作對照。反應管置于37 ℃反應1 h,后加入0.2 mL 0.6 mol/L四硼酸鉀溶液,沸水浴中煮沸3 min,迅速冷卻,在585 nm波長處進行操作,重復3 次。以每秒每克樣品中酶分解膠狀幾丁質產生1× 10—6N-乙酰葡萄糖胺為一個CHT活性單位,單位是10—6mol/(s·g),計算見公式(2):
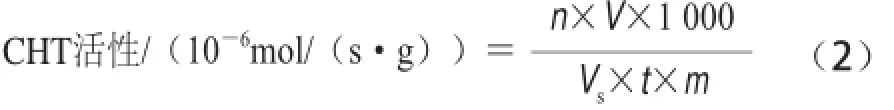
式中:n為從標準曲線查得的N-乙酰葡萄糖胺的量/μmol;V為樣品提取液總體積/mL;Vs為測定時所取樣品提取液體積/mL;t為酶促反應時間/s;m為樣品質量/g。GLU活性的測定參考曹建康等[18]方法并修改,稱3 g果皮組織,加入預冷5 mL的29 mg EDTA、0.1 g/L抗壞血酸、35 μL β-巰基乙醇、pH 5.2、0.1 mol/L的乙酸-乙酸鈉緩沖液,在冰浴中充分研磨,4 ℃、12 000×g離心30 min,上清液用于活性測定。200 μL 4 g/L昆布多糖為底物,以每秒每克樣品中酶分解昆 布多糖產生1×10—3葡萄糖為一個GLU活單位,單位是10—3mol /(s·g),計算見公式(3):
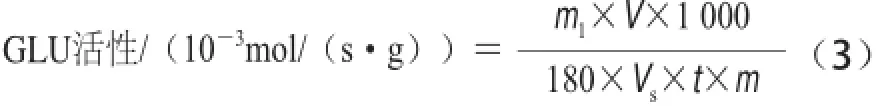
式中:m1為從標準曲線查得的葡萄糖的質量/mg;V為樣品提取液總體積/mL;Vs為測定時所取樣品提取液體積/mL;t為酶促反應時間/s;m為樣品質量/g;180為葡萄糖相對分子質量。
1.2.7 蛋白含量測定
參照Brad ford[20]方法,以牛血清蛋白為標準蛋白作標準曲線,計算蛋白含量。
1.3 數據分析
用SPSS 17.0軟件對實驗數據進行多重差異顯著分析和Excel 2007軟件對實驗數據進行分析并繪制圖表。
2 結果與分析
2.1 激發子處理對杏果抗黑斑病的影響
激發子處理均可明顯降低杏果損傷接種A. alternata的病斑直徑(P<0.05)(圖1),但不同質量濃度激發子處理之間無顯著性差異(P>0.05),其中以14.4 μg/mL的處理效果較好。處理與損傷接種的間隔時間對病斑直徑的影響較大(P<0.01),當間隔48、60、72 h后病斑直徑僅分別為同期對照的53.3%、49.1%和35.5%,說明不同處理時間而效果不同,誘導72 h時的抗病效果為處理組中最好。
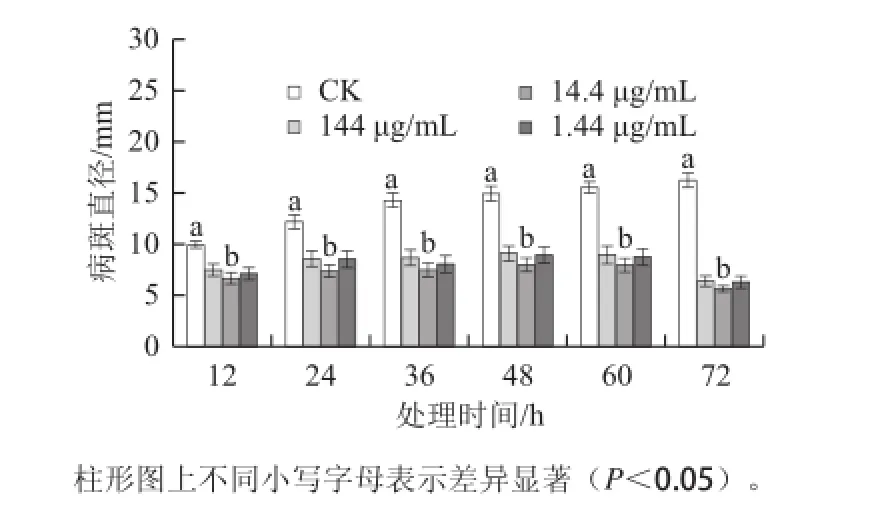
圖1 T. roseum seum蛋白激發子處理對杏果抗黑斑病的影響Fig.1 Effect of treatment with protein elicitor from T. roseum on resistance in apricot fruits against Alternaria rot圖2 T.rosseeuumm蛋白激發子處理對杏果生成速率(aa)和H2O2含量(bb)的影響Fig.2 Effect of treatment with protein elicitor from T. roseum on· generation rate and H2O2content in apricot fruit tissue
2.2 激發子處理對杏果組織O2—·生成速率和H2O2含量的影響
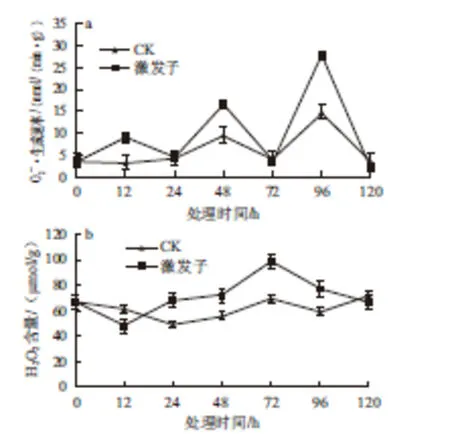
圖1 T. roseum seum蛋白激發子處理對杏果抗黑斑病的影響Fig.1 Effect of treatment with protein elicitor from T. roseum on resistance in apricot fruits against Alternaria rot圖2 T.rosseeuumm蛋白激發子處理對杏果生成速率(aa)和H2O2含量(bb)的影響Fig.2 Effect of treatment with protein elicitor from T. roseum on· generation rate and H2O2content in apricot fruit tissue
2.3 激發子處理后對杏果組織SOD、POD、PPO活性的影響
激發子處理能顯著提高杏果組織SOD、POD和PPO的活性(圖3)。在處理后12 h內,處理組和對照組果實SOD活性無明顯變化,之后處理組活性逐漸上升,48 h達到最大值,后期逐漸下降,而對照組在24 h之后呈先下降后上升趨勢,且在48 h處理組SOD活性比對照組高出2.25 倍(圖3a);對照組或處理組果實的POD活性均呈現先增加后降低趨勢,但在12 h后處理組POD活性顯著高于對照,在72 h達到最大值,其活性為對照的1.5 倍(圖3b);處理組果實的PPO活性也呈現先上升后下降的趨勢,尤其在處理48 h后,其活性顯著高于對照,且為對照的3.86 倍,而對照組隨時間的變化不大(圖3c)。
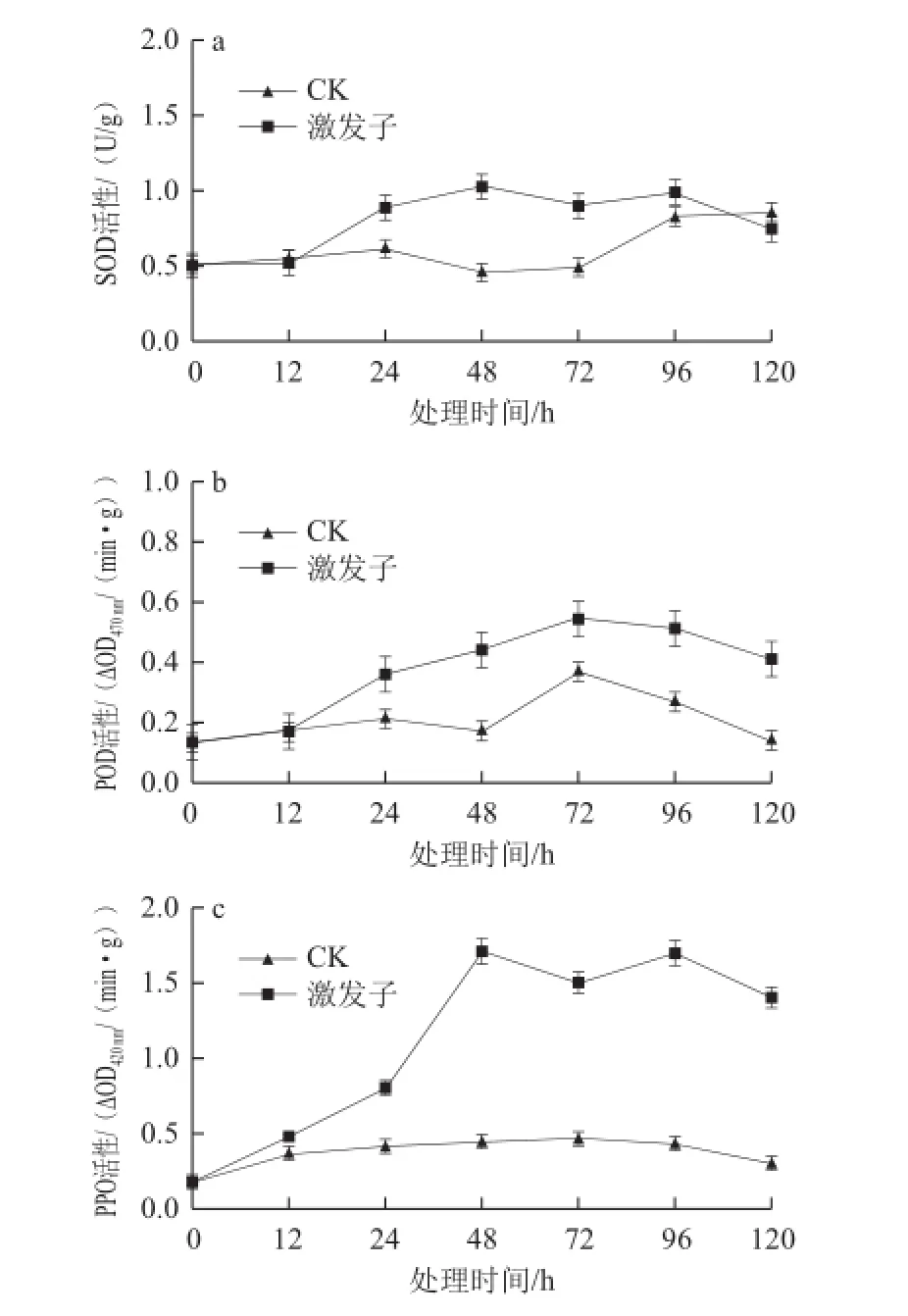
圖 3 T. rseum蛋白激發子處理對杏果組織SOD(a)、PODD(bb)、PPO(c)活性的影Fig.3 The activities of SOD (a), POD (b) and PPO (c) in apricot fruit tissue treated with protein elicitor from T. roseum
2.4 激發子處理后對杏果組織GLU和CHT活性的影響
激發子處理均能有效提高杏果組織GLU、CHT的活性(圖4)。杏的GLU活性呈現先上升后下降趨勢,在12 h后處理組GLU活性高于對照組,72 h達到最大值,其活性是對照的1.3 倍(圖4a);在120 h內處理組果實CHT活性明顯高于對照,且活性呈上升趨勢,在120 h處理組果實的CHT活性最大,為對照的2 倍(圖4b)。
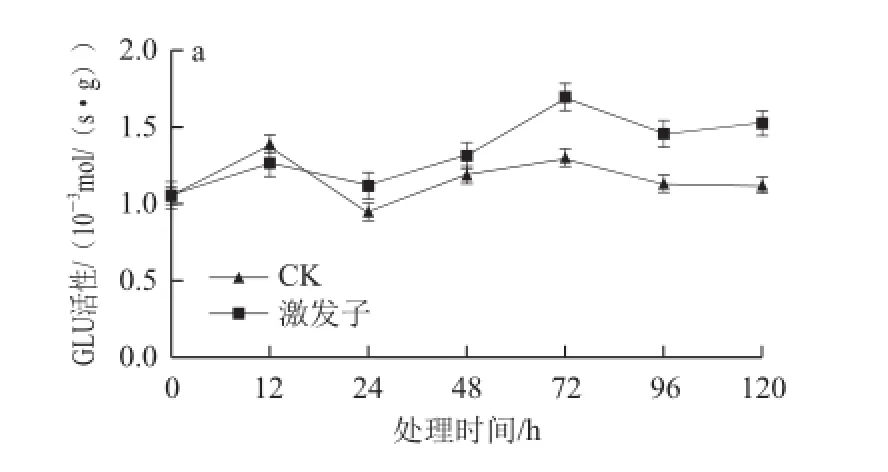

圖4 4 T. roseum seum蛋白激發子處理對杏果組織GLU(a)和CHT(b)HTb活性的影響Fig.4 The activities of GLU (a) and CHT (b) in apricot fruit tissue treated with protein elicitor from T. roseum
3 討論與結論
本研究結果表明,T. roseum菌體粗蛋白激發子可顯著抑制‘蘭州大接杏’損傷接種A. alternate病斑直徑的擴展,促進活性氧(reactive oxygen species,ROS)爆發,增強SOD、POD和PPO的活性以及病程相關(pathogenesis related,PR)蛋白的積累,從而誘導了果實的抗病性。不同質量濃度激發子對果實抗病性的誘導效果存在差異,其中14.4 μg/mL的誘導效果較好。處理與損傷接種間隔時間對誘導效果影響較大,以間隔72 h時效果最好。該結果與前人在使用Harpin處理甜瓜[13]和甜橙[21]的結果類似,此外,真菌源蛋白激發子采前處理還可誘導葡萄對采后病害的抗性[22]。由此表明,微生物源蛋白激發子具有誘導果蔬抗病性的潛力,一定程度上可抑制采后病害的發生和發展。
ROS可使寄主產生多種抗性反應,如細胞壁加厚、過敏性細胞死亡、抗性基因的激活以及抗性物質的累積[23]。氧爆發是植物細胞對病原菌侵染和激發子處理后產生的一種重要早期事件[24]。本研究結果顯示,T. roseum菌體粗蛋白激發子導致了杏果實O2—·產生速率和H2O2含量短暫而快速的升高,O2—·在處理后48 h達到最大 值,SOD活性也在48 h迅速增加,同時72 h時H2O2含量達到最大值,這可能是激發子處理后杏產生的O2—·在SOD作用下可迅速轉化成H2O2,而在96 h處O2—·大量生成可能與其衰老有關。類似的研究[9,25]報道表明,真菌源蛋白激發子處理可短暫而快速的生成大量H2O2、NO,且激發煙草、水稻等抗病性增強。可見氧爆在介導微生物源激發子誘導植物抗病性中具有重要作用,近年來研究者發現ROS尤其是H2O2在植物SAR中可能是作為信號傳導分子而發揮作用[26],其中位于原生質膜上的NADPH氧化酶利用NADPH作為電子供體,通過還原O2能夠催化O2—·的產生,是植物氧爆過程中產生ROS的主要來源[27]。這些結果表明,蛋白激發子誘導可產生大量的ROS,其可能在激發子誘導的杏采后抗病反應中起到了重要作用,但是NOX與前期ROS產生的關系有待進一步研究。
盡管ROS在增強植物的抗病性中有著重要的作用,然而植物體內積累的ROS含量過高時就會對植物組織造成毒害作用,導致脂質過氧化、損傷 膜脂和膜蛋白、加速植物衰老等[28],為了避免ROS的傷害,植物形成了自己的抗氧化系統使ROS不能過量積累。本結果顯示T. roseum激發子能誘導杏果SOD、POD及PPO的活性增加,在蛋白激發子處理番茄葉片[29]和棉花葉片[30]時也觀察到這一現象。可見SOD作為調控ROS的關鍵酶,可通過催化組織內O2—·快速轉變為H2O2。而POD可通過氧化酚類物質分解H2O2,與對照相比激發子處理的杏果其POD在72 h顯著增加,由此表明在H2O2大量生成的時候,其PO D的活性也隨之增強達到保護杏的抗氧化能力。同時POD和PPO活性增加可促進果實木質素合成,木質化不僅形成抵御病原菌侵入的物理屏障物,而且PPO可利用O2將酚類物質氧化形成的醌類物質對病原菌也有很強的毒害作用[31]。PPO的活性隨著處理時間而增強,在處理期間48 h時其活性最強。類似報道表明,Harpin處理可誘導甜瓜POD活性增強[13];真菌源激發子處理誘導水稻、番茄的PPO和POD活性增強從而提高抗性[25,32]。這可能是通過調節蛋白激發子處理植物和果實組織后其體內ROS水平和一系列抗性相關酶SOD、POD、PPO活性之間的平衡關系來維持自身的抗病性,同時也可能因為PPO利用ROS使細胞壁加厚有關,抵御病原物侵入。
激發子被識別后,信號傳導網絡被激活、胞內相關基因重編碼,與此同時具有PR功能蛋白也被激活表達[33]。本研究顯示,激發子處理后杏GLU和CHT活性較對照組均有顯著增加,隨著處理時間的推移其活性逐漸增強。與此類似的報道表明,Harpin處理可誘導甜瓜CHT活性增強[13],目前未見真菌源激發子誘導植物組織產生PR蛋白的報道。這可能由于激發子被識別后產生ROS,其作為信號分子激活了果實體內的SA積累,誘導一系列抗氧化酶活性的變化,同時激活了GLU和CHT的大量表達,從而誘導杏的抗病性。因此該蛋白激發子處理后誘導杏果抗病性過程中產生的ROS與GLU、CHT可通過SA聯系起來進行進一步研究。
T. roseum菌體粗蛋白激發子處理能顯著降低接 種A. alternate‘蘭州大接杏’杏果病斑直徑的擴展。其中14.4 μg/mL的蛋白激發子抗病效果最顯著,72 h時抗病效果最好。用激發子處理后有短暫而大量的ROS產生,并誘導了杏果抗性相關酶SOD、PPO、POD、CHT、GLU等活性的提高。由此表明,T. roseum菌體粗蛋白激發子處理可通過影響果實ROS及抗性相關酶來增強杏對采后病害的控制。激發子一般是通過早期信號被激活、影響能量代謝最后通過苯丙烷代謝過程形成抗性相關物質來體現其誘導抗性,因此本實驗杏抗病性的研究中需進一步系統研究能量及苯丙烷代謝過程進一步闡述其誘導抗性作用機理;同時需進一步純化菌體粗蛋白,研究何種蛋白在抗性中起關鍵性作用。
[1] 張波, 韓舜愈, 蔣玉梅, 等. 硅酸鈉處理對杏品質和風味物質的影響[J].農業工程學報, 2011, 27(6): 375-381.
[2] 于希志. 試談我國杏生產現狀與對策[J]. 北方果樹, 2003, 5(9): 25-25. [3] 陳松江, 畢陽, 李永才, 等. 一氧化氮處理對杏果實貯藏品質的影響[J].食品科技, 2011, 36(6): 46-50.
[4] ANTUNES M D C, MIGUEL M G, NEVES M A, et al. Quality changes during storage of apricot (Prunus armeniaca L.) cv. ‘Beliana’and ‘Lindo’ trated with calcium[M]//NUNES C. Environmentall y friendly and safe technologies for quality of fruit and vegetables. Universidade do A lgarve, 2010: 117-120.
[5] 王文娟, 李永才, 畢陽, 等. 硼酸鹽處理對采后杏果軟腐病的控制及貯藏品質的影響[J]. 食品科學, 2012, 34(18): 280-284.
[6] 范存斐, 畢陽, 王云飛, 等. 水楊酸對厚皮甜瓜采后病害及苯丙烷代謝的影響[J]. 中國農業科學, 2012, 45(3): 584-589.
[7] 邱德文. 生物農藥與生物防治發展戰略淺談[J]. 中國農業科技導報, 2011, 13(5): 88-92.
[8] 邱德文. 微生物蛋白農藥研究進展[J]. 中國生物防治, 2004, 20(2): 91-94.
[9] KULYE M, LIU Hua, ZHANG Yu liang, et al. Hrip1, a novel protein elicitor from necrotrophic fungus, Alternaria tenuissima, elicits cell death, expression of defence-related genes and systemic acquired resistance in tobacco[J]. Plant, Cell and Environment, 2012, 35(12): 2104-2120.
[10] 彭學聰, 楊秀芬, 邱德文, 等. 蛋白激發子Hrip1基因在擬南芥中表達可提高植株的耐鹽耐旱能力[J]. 作物學報, 2013, 39(8): 1345-1351.
[11] QIU Dewen, MAO Jianjun, YANG Xiufen, et al. Expression of an elicitor-encoding gene from Magnaporthe grisea enhances resistance against blast disease in transgenic rice[J]. Plant Cell Reports, 2009, 28(6): 925-933.
[12] CAPDEVILLE G D, BEER S V, WILSON C L, et al. Some cellular correlates of harpin-induced resistance to blue mold of apples[J]. Tropical Plant Patholo gy, 2008, 33: 103-113.
[13] BI Yang, TIAN Shiping, ZHAO Jie, et al. Harpin induces local and systemic resistance against Trichothecium roseum in harvested Hami melons[J]. Postharvest Biology and Technology, 2005, 38(2): 183-187.
[14] ZHAO Mingzhi, YANG Xiufen, ZHANG Ming, et al. Purification and bioactivities of a protein growth-activator from Aternaria tenuissima[J]. Chinese Journal of Biological Control, 2007, 23(2): 170-173.
[15] 周德慶. 微生物學實驗教程[M]. 2版. 北京: 高等教育出版社, 2006.
[16] 王云飛, 畢陽, 任亞琳, 等. 硅酸鈉處理對厚皮甜瓜果實采后病害的控制及活性氧代謝的作用[J]. 中國農業科學, 2012, 45(11): 2242-2248.
[17] 高雄杰. 采后硅酸鈉處理對‘玉金香’甜瓜活性氧及其代謝體系的誘導[D]. 蘭州: 甘肅農業大學, 2010.
[18] 曹建康, 姜微波, 趙玉梅. 果 蔬采后生理生化實驗指導書[M]. 北京:中國輕工業出版社, 2007.
[19] 任亞琳, 畢陽, 葛永紅, 等. BTH浸泡處理對厚皮甜瓜采后病害的控制及貯藏品質的影響[J]. 食品科學, 2013, 34(2): 267-272.
[20] BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding[J]. Analytical Biochemistry, 1976, 72: 248-254.
[21] LUCON C M M, GUZZO C O, de JESUS C O, et al. Postharvest harpin or Bacillus thuringiensis treatments suppress citrus black spot in ‘Valencia’ oranges[J]. Crop Protection, 2010, 29(7): 766-772.
[22] 關文強, 李寧, 段雙科. 真菌源激發子對采后葡萄果皮抗性產生的誘導[J]. 西北植物學報, 2005, 25(12): 2408-2412.
[23] VELLOSILLO T, VICENTE J, KULASEKARAN S, et al. Emerging complexity in reactive oxygen species production and signaling during the response of plants to pathogens[J]. Plant Physiology, 2010, 154(2): 444-448.
[24] ALBERT P, MIYA A, HIRATSUKA K, et al. A high-throug hput evaluation system for Arabidopsis mutants for defense signaling[J]. Plant Biotechnology, 2006, 23(5): 459-466.
[25] 袁肖寒, 顧成波, 邱德文, 等. 新型真菌源激活蛋白誘導水稻抗病性及其生理機制[J]. 植物研究, 2013, 33(2): 220-224.
[26] QUAN Lijuan, ZHANG Bo, SHI Weiwei, et al. Hydrogen peroxide in plants: a versatile molecule of the reactive oxygen species network[J]. Journal of Integrative Plant Biology, 2008, 50(1): 2-18.
[27] HYON G S, IKEDA K, HOSOGI N, et al. Inhibitory effects of antioxidant reagent in reactive oxygen species generation and penetration of appressoria of Alternaria alternata Japanese pear pathotype[J]. Phytopathology, 2010, 100: 840-847.
[28] GONZALEZ A G A, VILLA J A, AYALA J F, et al. Improvement of the antioxidant status of tropical fruits as a secondary response to some postharvest treatments[J]. Trends in Food Science and Technology, 2010, 21: 475-482.
[29] 李麗, 劉崢, 楊秀芬, 等. 植物激活蛋白對番茄防御酶活性的影響[J].湖南農業大學學報: 自然科學版, 2008, 34(5): 534-537.
[30] 張志剛, 楊曉萍, 盧翰, 等. 細極鏈格孢菌蛋白激發子對棉花生長相關酶活性的影響[J]. 棉花學報, 2010, 22(2): 120-124.
[31] 李永才, 畢陽, 胡潔, 等. 真菌菌絲細胞壁提取物對馬鈴薯塊莖組織抗干腐病的誘導效應[J]. 西北植物學報, 2009, 29(4): 749-753.
[32] MANDAL S, MITRA A. Reinforcement of cell wall in roots of Lycopersicon esculentum through induction of phenolic compounds and lignin by elicitors[J]. Physiological and Molecular Plant Pathology, 2007, 71(4): 201-209.
[33] GENDA Y, KANDA A, HAMADA H, et al. Two amino acid substitutions in the coat protein of pepper mild mottle virus are responsible for overcoming the L4gene-mediated resistance in capsicum spp[J]. Phytopathology, 2007, 97(7): 787-793.
Mechanism of Alternaria Rot Resistance Induced in Apricot Fruits by Protein Elicitor from Trichothecium roseum
YU Xiaoyan, BI Yang, LI Yongcai*, YAN Lu, SHEN Keping, HU Lingang, TANG Yamei
(College of Food Science and Engineering, Gansu Agricultural University, Lanzhou 730070, China)
The effect of protein elicitor from the nonhost Trichothecium roseum on black rot of apricot fruits and its possible induction mechanisms were studied. The inductive effect and its time-dependent manner of the protein elicitor in apricot fruits inoculated with Alternaria alternate at different times after treatment were studied. The effect of selected optimum concentration of protein elicitor on reactive oxygen species (ROS) and resistance-related enzymes in apricot fruit tissue were also evaluated. The lesion development of wound-inoculated apricot fruits were signifi cantly reduced by protein elicitor treatment(P < 0.05), and at 72 h after treatment with 14.4 μg/mL protein elicitorthe best induction effect on apricot fruits was observed. The rate of superoxide anion (O2-?) production and the content of hydrogen peroxide (H2O2) in fruit tissue reached the peak at 48 and 72 h, respectively. The activities of superoxide dismutase (SOD), peroxidase (POD), polyphenol oxidase (PPO), β-1,3-glucanase (GLU) and chitinase (CHT) in apricot fruit tissue treated with the protein elicitor were also increased. The protein elicitor treatment could signifi cantly increase the disease resistance of apricot fruit tissue, and the possible mechanism might be due to the outburst of RO S and the improved activities of resistance-related enzymes. So protein elicitor from Trichothecium roseum may be a potential resistance inducer and can be further applied to postharvest diseases control.
Trichothecium roseum; protein elicitor; apricot fruit; reactive oxygen species; induced disease resistance; Alternaria rot
TS201.3
A
1002-6630(2015)04-0254-06
10.7506/spkx1002-6630-201504050
2014-04-21
甘肅省農牧廳生物技術專項(GNSW-2011-11)
余曉燕(1988—),女,碩士研究生,研究方向為采后生物學與技術。E-mail:y198808150028@163.com
*通信作者:李永才(1972—),男,副教授,博士,研究方向為果蔬采后生物學與技術。E-mail:liyongcai@gsau.edu.cn
