外源蔗糖對蘿卜幼苗品質及代謝酶活性的影響
2015-12-13 05:41:18趙曉幗羅云波
食品科學 2015年9期
關鍵詞:生長
趙曉幗,朱 毅,羅云波
(中國農業大學食品科學與營養工程學院,北京 100083)
外源蔗糖對蘿卜幼苗品質及代謝酶活性的影響
趙曉幗,朱 毅*,羅云波
(中國農業大學食品科學與營養工程學院,北京 100083)
以“滿堂紅”品種的蘿卜為材料,探究不同質量濃度的外源蔗糖噴灑處理對不同生長天數的蘿卜幼苗營養品質(花青素、葉綠素、類胡蘿卜素、總酚和芥子油苷等含量)及代謝酶(過氧化物酶(peroxidase,POD)和苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL))活性的影響。結果表明:對于生長3、5 d和7 d的蘿卜幼苗,0.4 g/100 mL和0.6 g/100 mL蔗糖處理均能夠顯著提高其花青素、葉綠素、類胡蘿卜素、總酚、芥子油苷含量及POD、PAL活性(P<0.05),而0.2 g/100 mL蔗糖處理對其營養物質含量及抗氧化性的影響不顯著(P>0.05);0.4 g/100 mL和0.6 g/100 mL蔗糖處理在改善其營養價值方面的差別不大。因此,在農業生產中,可采用質量濃度0.4 g/100 mL蔗糖噴灑處理蘿卜幼苗來提高其營養價值。
蘿卜;幼苗;蔗糖;品質;代謝酶
蘿卜是一種重要的十字花科蔬菜,廣泛被人類食用[1]。十字花科蔬菜中含有豐富的營養物質,如VC、VE、酚類化合物、類胡蘿卜素、花青素、葉綠素和芥子油苷等。有研究報道,十字花科芽苗菜中營養物質(如芥子油苷、類胡蘿卜素和酚類化合物)的含量是成熟蔬菜(如蘿卜、西蘭花、甘藍等)的15~50 倍[2-4],國內外研究者采用不同的處理方式來提高其營養價值[1,5-9]。蔗糖是調控植物生長發育的信號分子,關于其促進植物營養物質累積的文獻已有報道,但是蔗糖噴灑處理對蘿卜幼苗品質的影響還未見報道。鑒于此,本實驗以“滿堂紅”品種蘿卜為試材,探究了不同質量濃度的外源蔗糖噴灑處理對不同生長天數的蘿卜幼苗營養品質及代謝酶活性的影響,以期為農業生產提供一定的理論參考。
1 材料與方法
1.1 材料與試劑
“滿堂紅”蘿卜種子 北京京研盛豐種苗研究所。
2-丙烯基芥子油苷(sinigrin) 美國Sigma公司;Folin-酚試劑、沒食子酸、乙醇、愈創木酚、過氧化氫國藥集團化學試劑有限公司。
1.2 儀器與設備
DK-98-I恒溫水浴鍋 天津市泰斯特儀器公司;HZC-250恒溫振蕩培養箱 江蘇太倉實驗設備廠;5424R Eppendorf離心機 德國Eppendorf公司;M200 Pro多功能酶標儀 奧地利帝肯公司。
1.3 蘿卜苗的栽培
挑選顆粒飽滿、無霉變的滿堂紅蘿卜種子用蒸餾水清洗后,置于5 mL/L的NaClO溶液中浸泡30 min。將NaClO溶液瀝干并用蒸餾水清洗3~4 次后,置于蒸餾水中浸泡8 h。將浸泡后的種子均勻播撒在30 cm×20 cm的育苗盤內,育苗盤上鋪有4 層紗布以防止育苗過程中水分過度流失。將播撒種子后的育苗盤置于23 ℃的培養室中避光催芽2 d后,轉移至16 h光照/8 h黑暗的條件下培育生長,晝/夜溫度為23 ℃/20 ℃,相對濕度為70%~75%。在培育過程中,每隔8 h分別用蒸餾水(對照)、0.2、0.4、0.6 g/100 mL蔗糖噴灑種子或幼苗。采集生長3、5 d和7 d的蘿卜苗,在液氮中快速冷凍后,經封口袋包裝置于—80 ℃冰箱中保存,以供后續實驗用。
1.4 測定方法
花青素含量參考文獻[10]的方法測定,單位是U/g,表示每克鮮質量樣品所含的花青素單位數;葉綠素和類胡蘿卜素含量參考文獻[11]的方法測定;總酚含量參考文獻[12]的方法測定;芥子油苷含量的測定采用氯化鈀分光光度計法[13-14];苯丙氨酸解氨酶(phenylalanineammonialyase,PAL)活力參考文獻[15]的方法測定,以每小時每克樣品酶促反應體系吸光度增加0.01為1 個PAL活力單位,記為U;過氧化物酶(peroxidase,POD)活力采用愈創木酚法測定[16],以每克鮮質量樣品每分鐘吸光度變化量增加1.0為1個POD活力單位,記為U。
1.5 數據統計分析
所有實驗重復3 次,營養物質含量及酶活力均平行測定3 次,數據均采用SPSS 16.0(SPSS Inc., Chicago, IL,USA)軟件進行鄧肯氏多重差異分析,單因素方差分析(analysis of variance,ANOVA)(P<0.05)。
2 結果與分析
2.1 蔗糖對蘿卜幼苗花青素含量的影響
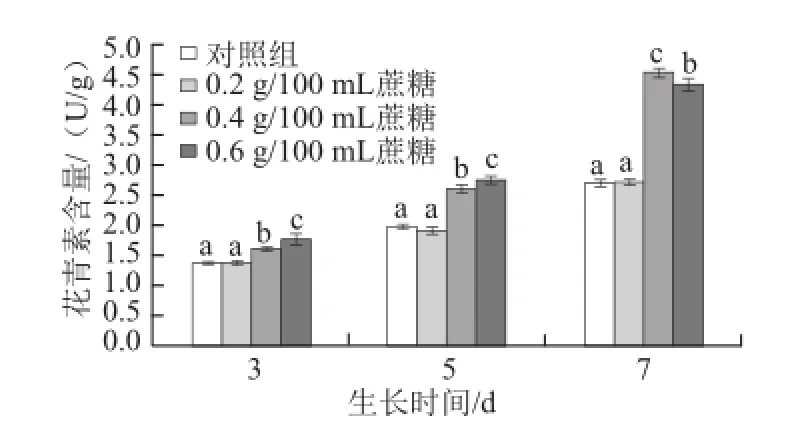
圖1 不同質量濃度蔗糖對不同生長天數的蘿卜苗花青素含量的影響Fig.1 Effects of different concentrations of sucrose on anthocyanin content in radish sprouts
花青素是植物中普遍存在的一類天然色素。在蘿卜苗中,花青素主要集中于子葉和莖中,不僅可以作為食用色素,而且對人體還具有一定的抗氧化和抗癌活性功效[10]。在生長過程中,分別采用蒸餾水(對照)、0.2、0.4、0.6 g/100 mL蔗糖溶液噴灑處理蘿卜苗,結果見圖1,隨著生長天數的增加,其花青素含量越來越高。與對照組相比,生長3、5 d和7 d的蘿卜苗,0.4 g/100 mL和0.6 g/100 mL蔗糖噴灑處理能顯著提高其花青素含量(P<0.05),且隨著蘿卜苗生長天數的延長,0.4 g/100 mL蔗糖處理對其合成和累積的促進效果越顯著(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理比對照組分別提高17.5%和29.2%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高31.0%和38.2%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高68.2%和60.9%,而0.2 g/100 mL蔗糖處理對花青素含量無顯著影響(P>0.05)。Guo Rongfang等[17]研究表明,蔗糖可以提高西蘭花幼苗中的花青素含量,與本實驗結果相一致。有研究報道稱,在擬南芥中,蔗糖作為一種信號分子能夠促進花青素合成過程中相關基因的表達進而提高花青素的含量[18]。當處理蘿卜苗的蔗糖質量濃度提高時,其誘導花青素合成過程中相關基因表達量增加,進而提高花青素的含量。隨著生長天數的增加,適宜質量濃度的蔗糖持續誘導相關基因的表達,使得其促進花青素合成的效果越顯著。
2.2 蔗糖對蘿卜幼苗葉綠素含量的影響
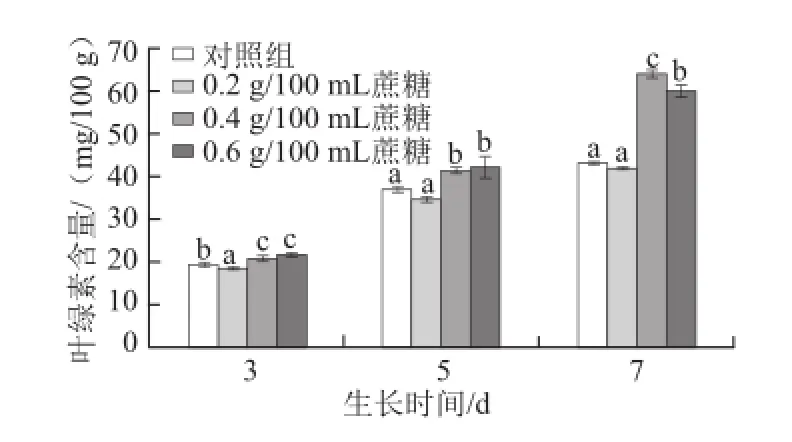
圖2 不同質量濃度蔗糖對不同生長天數的蘿卜苗葉綠素含量的影響Fig.2 Effects of different concentrations of sucrose on chlorophyll content in radish sprouts
葉綠素是植物體進行光合作用必需的元素。如圖2所示,0.4 g/100 mL和0.6 g/100 mL的外源蔗糖處理能顯著提高蘿卜幼苗葉綠素的含量(P<0.05),且隨著生長天數的延長,0.4 g/100 mL蔗糖促進葉綠素積累的效果越明顯(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理組葉綠素含量比對照組分別提高7.2%和11.9%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高12.3%和14.3%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高48.2%和38.9%。有研究報道,葉綠體的形成會受到糖的影響,葡萄糖處理能夠抑制擬南芥幼苗葉綠體的形成[19],而本實驗發現蔗糖處理后蘿卜幼苗葉綠素含量顯著提高,有利于幼苗進行光合作用,與劉麗萍[20]研究結果一致,此差異可能是由于植物種類和糖種類的不同造成的。隨著生長天數的延長,葉片面積逐漸增大,光合作用越明顯,適宜質量濃度蔗糖的促進作用越顯著。
2.3 蔗糖對蘿卜幼苗類胡蘿卜素含量的影響
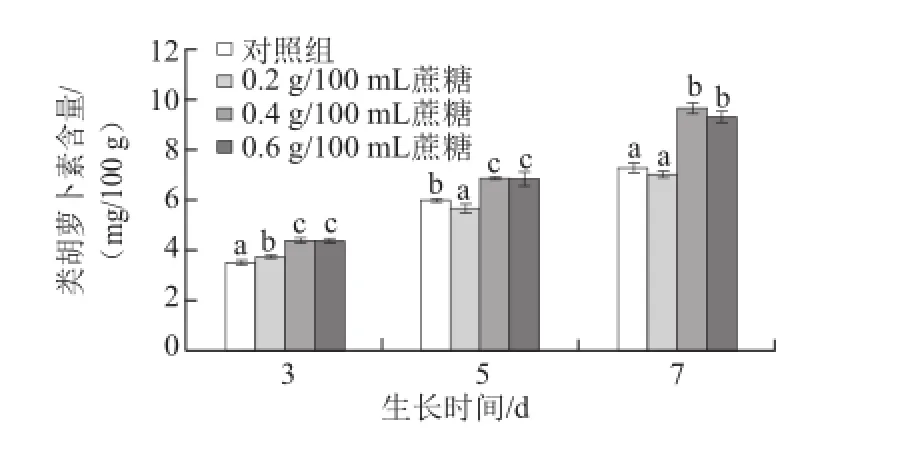
圖3 不同質量濃度蔗糖對不同生長天數的蘿卜苗類胡蘿卜素含量的影響Fig.3 Effects of different concentrations of sucrose on carotenoid content in radish sprouts
類胡蘿卜素是一類對人體有較多生理功能的色素。如圖3所示,隨著蘿卜苗生長天數的增加,其類胡蘿卜素含量逐漸升高;0.4 g/100 mL和0.6 g/100 mL蔗糖處理均能顯著地提高生長3、5 d和7 d的蘿卜苗中類胡蘿卜素的含量(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理組的類胡蘿卜素含量比對照組分別提高24.6%和24.4%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高11.1%和13.7%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高32.4%和27.6%。該結果與葉綠素的變化趨勢一致,原因可能在于這兩種色素的合成途徑較相似[21],且含量呈現正相關關系[22]。
2.4 蔗糖對蘿卜幼苗總酚含量的影響
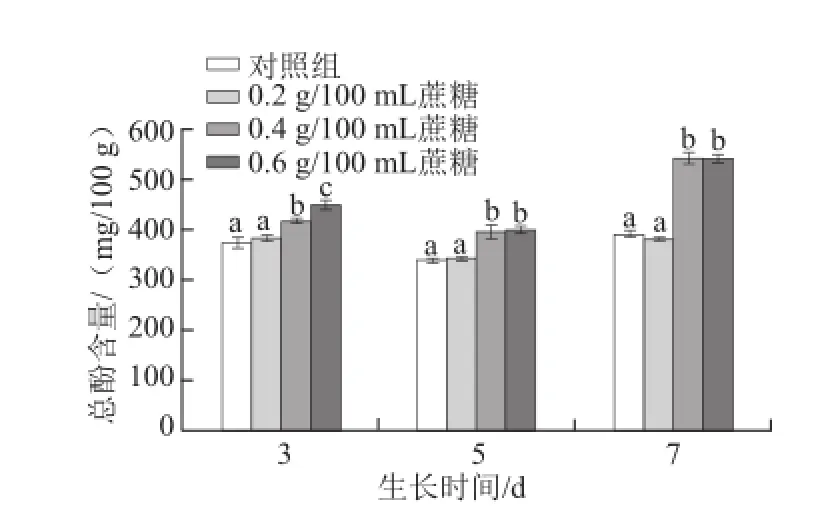
圖4 不同質量濃度蔗糖對不同生長天數的蘿卜苗總酚含量的影響Fig.4 Effects of different concentrations of sucrose on total phenolic content in radish sprouts
酚類化合物在植物抗氧化體系中具有重要作用[17]。酚類化合物的合成或積累受到多種外界環境條件和外源誘導因子的影響[23-25]。如圖4所示,0.4 g/100 mL和0.6 g/100 mL蔗糖處理均能顯著提高生長3、5 d和7 d的蘿卜苗中總酚含量(P<0.05)。隨著天數的延長,0.4 g/100 mL蔗糖促進其合成或積累的效果越明顯(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理組的總酚含量比對照組分別提高11.8%和20.4%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高16.5%和17.4%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高38.2%和38.1%。該發現與Guo Rongfang等[17]研究結果一致,可能是由于蔗糖引起的滲透壓所致。外源蔗糖會提高蘿卜幼苗體內的滲透壓,造成細胞失水,影響其新陳代謝,進而提高總酚含量。隨著生長天數的增加,適宜質量濃度的蔗糖持續作用于蘿卜苗,使得其促進總酚含量提高的效果越明顯。
2.5 蔗糖對蘿卜幼苗芥子油苷含量的影響
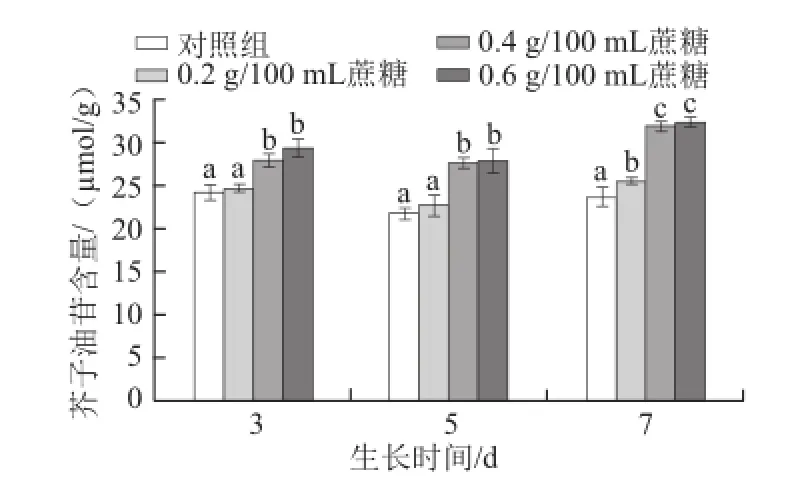
圖5 不同質量濃度蔗糖對不同生長天數的蘿卜苗芥子油苷含量的影響Fig.5 Effects of different concentrations of sucrose on total glucosinolate content in radish sprouts
芥子油苷作為十字花科蔬菜中廣泛存在的次級代謝產物,其對人體有著重要的生理功能,如抗癌和保護細胞等[26]。芥子油苷的合成受到多種因素的調控,如植物激素[25]、光照[27]、采后包裝[28]、水分[29-30]和二氧化碳[31]等。如圖5所示,對于生長3 d和5 d的蘿卜苗,0.4 g/100 mL和0.6 g/100 mL蔗糖能顯著提高其芥子油苷含量(P<0.05),而對于生長7 d的蘿卜苗,0.2、0.4 g/100 mL和0.6 g/100 mL蔗糖均能顯著提高其含量(P<0.05)。隨著生長天數的增加,0.4 g/100 mL和0.6 g/100 mL的蔗糖促進芥子油苷合成或積累的效果越明顯(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理比對照組分別提高15.0%和21.0%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高26.9%和28.2%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高35.1%和37.0%。Guo Rongfang等[17]研究發現,用相同質量濃度蔗糖和甘露糖醇處理西蘭花幼苗,均能提高芥子油苷含量,但兩者對其含量的影響差別不大,推斷造成此現象的原因可能在于滲透壓的作用。外源蔗糖會提高蘿卜幼苗體內的滲透壓,造成細胞失水,進而提高芥子油苷的含量。而Guo Rongfang等[6]研究發現,蔗糖處理西蘭花幼苗可提高芥子油苷合成過程中相關基因的表達量,進而提高芥子油苷含量。
2.6 蔗糖對蘿卜幼苗PAL活性的影響
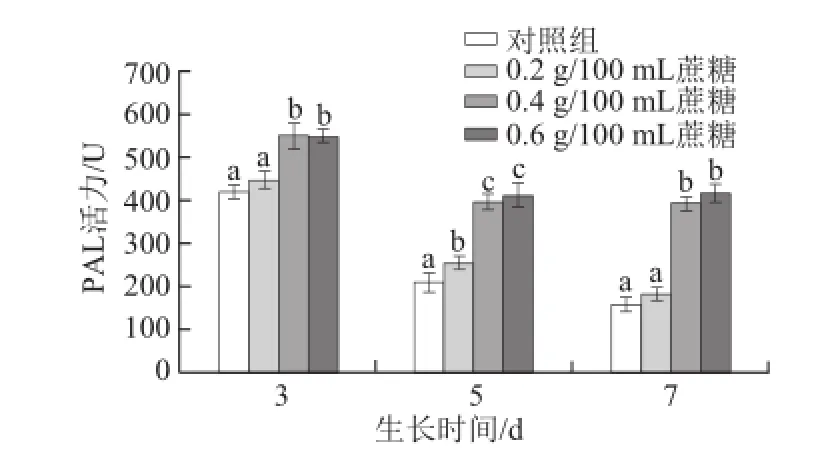
圖6 不同質量濃度蔗糖對不同生長天數的蘿卜苗PAL活性的影響Fig.6 Effects of different concentrations of sucrose on PAL activity in radish sprouts
研究報道稱,苯丙氨酸解氨酶(PAL)是植物體內酚類化合物合成過程中重要的一種酶[32],PAL活性會受到外源茉莉酸甲酯和外界創傷的影響[24-25]。如圖6所示,隨著生長天數的增加,對照組蘿卜苗PAL活性降低;0.4 g/100 mL和0.6 g/100 mL蔗糖處理蘿卜苗均能顯著提高其PAL活性(P<0.05),且生長天數越長其促進效果越明顯(P<0.05)。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理組的PAL酶活力比對照組分別提高30.7%和30.2%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高86.4%和93.7%;7 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高145.8%和159.7%。該結果與總酚含量變化趨勢相一致,其原因在于PAL是酚類化合物合成過程中較為重要的酶,PAL活性提高有利于總酚的合成過程中相關基因的表達而使其含量提高[33]。Guo Rongfang等[17]研究發現,用蔗糖處理西蘭花幼苗,也可以顯著提高其PAL活性。據此可推斷,蔗糖可能通過誘導PAL合成或調控過程中相應基因的表達而導致其活性的增強。隨著生長天數的增加,適宜質量濃度蔗糖持續作用的累積效應增強,使得其促進PAL活性提高的效果更顯著。
2.7 蔗糖對蘿卜幼苗POD活性的影響
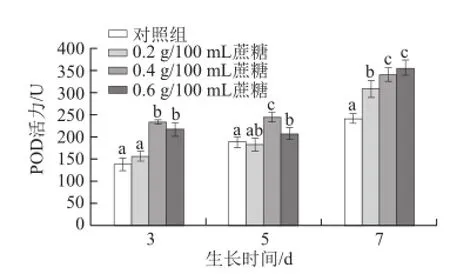
圖7 不同質量濃度蔗糖對不同生長天數的蘿卜苗POD活性的影響Fig.7 Effects of different concentrations of sucrose on POD activity in radish sprouts
過氧化物酶(POD)是植物體一種重要的組織抗氧化酶。如圖7所示,隨著生長天數的增加,POD活性逐漸增強。對于生長3 d和5 d的蘿卜苗,0.4 g/100 mL和0.6 g/100 mL蔗糖處理均能顯著提高其活性(P<0.05),而0.2 g/100 mL蔗糖處理效果不明顯(P>0.05)。對于生長7 d的蘿卜苗,0.2、0.4 g/100 mL和0.6 g/100 mL蔗糖處理均能顯著提高其活性(P<0.05)。對于生長3 d的蘿卜苗,蔗糖對其促進效果最好。3 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理組比對照組分別提高68.4%和56.4%;5 d苗:0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高29.6%和10.1%;7 d苗:0.2、0.4 g/100 mL和0.6 g/100 mL蔗糖處理分別提高27.7%、41.2%和47.2%。李小剛等[34]研究發現,用1 mmol/L的蔗糖處理在140 mmol/L NO3—脅迫下的黃瓜幼苗可以提高其POD活性,而當蔗糖濃度達5 mmol/L時,其抗氧化酶活性開始降低。賈曉琳[35]實驗結果表明,外源糖可以提高POD相關基因表達,進而提高其活性。與此相反,劉麗萍[20]研究發現,外源蔗糖處理蕎麥幼苗,可減低POD活性,造成此差異的原因可能在于植物種類及糖質量濃度的不同。
3 結 論
通過分析不同質量濃度蔗糖對不同生長天數的蘿卜幼苗營養物質含量的影響可知:對于生長3、5、7 d的蘿卜幼苗,0.4、0.6 g/100 mL蔗糖處理均能夠顯著提高其花青素、葉綠素、類胡蘿卜素、總酚、芥子油苷的含量,而0.2 g/100 mL蔗糖處理對其營養物質含量的影響并不顯著。
通過分析不同質量濃度蔗糖對不同生長天數的蘿卜幼苗抗氧化酶活性的影響可知:對于生長3、5、7 d的蘿卜幼苗,0.4、0.6 g/100 mL蔗糖處理均能夠顯著提高POD和PAL活力,而0.2 g/100 mL蔗糖處理對其活性影響并不顯著。
綜上所述,用不同質量濃度蔗糖對不同生長天數的蘿卜幼苗,0.4、0.6 g/100 mL蔗糖處理均能顯著提高其綜合營養價值,且隨著生長天數的增加,0.4 g/100 mL蔗糖處理的效果最好。因此,在農業生產中,可采用質量濃度為0.4 g/100 mL的蔗糖噴灑處理蘿卜幼苗來提高其營養價值。本研究提供了一種簡便、經濟的改善蘿卜幼苗營養品質的方法。隨著人們生活水平的提高和對飲食營養的重視,其在食品領域具有廣闊的應用前景。
[1] ZHOU Chenguang, ZHU Yi, LUO Yunbo. Effects of sulfur fertilization on the accumulation of health-promoting phytochemicals in radish sprouts[J]. Journal of Agricultural and Food Chemistry, 2013,61(31): 7552-7559.
[2] BAENAS N, MORENO D A, GARCIA-VIGUERA C. Selecting sprouts of brassicaceae for optimum phytochemical composition[J]. Journal of Agricultural and Food Chemistry, 2012, 60(45): 11409-11420.
[3] SINGH J, UPADHYAY A K, PRASAD K, et al. Variability of carotenes, vitamin C, E and phenolics in Brassica vegetables[J]. Journal of Food Composition and Analysis, 2007, 20(2): 106-112.
[4] KOPSELL D E, KOPSELL D A, RANDLE W A, et al. Kale carotenoids remain stable while flavor compounds respond to changes in sulfur fertility[J]. Journal of Agricultural and Food Chemistry, 2003,51(18): 5319-5325.
[5] PEREZ-BALIBREA S, MORENO D A, GARCIA-VIGUERA C. Improving the phytochemical composition of broccoli sprouts by elicitation[J]. Food Chemistry, 2011, 129(1): 35-44.
[6] GUO Rongfang, YUAN Gaofeng, WANG Qiaomei. Sucrose enhances the accumulation of anthocyanins and glucosinolates in Broccoli sprouts[J]. Food Chemistry, 2011, 129(3): 1080-1087.
[7] YUAN Gaofeng, WANG Xiaoping, GUO Rongfang, et al. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts[J]. Food Chemistry, 2010,121(4): 1014-1019.
[8] PEREZ-BALIBREA S, MORENO D A, GARCIA-VIGUERA C. Glucosinolates in Broccoli sprouts (Brassica oleracea var. italica) as conditioned by sulphate supply during germination[J]. Journal of Food Science, 2010, 75(8): C673-C677.
[9] SCHONHOF I, BLANKENBURG D, MULLER S, et al. Sulfur and nitrogen supply influence growth, product appearance, and glucosinolate concentration of broccoli[J]. Journal of Plant Nutrition and Soil Science, 2007, 170(1): 65-72.
[10] TENG S, KEURENTJES J, BENTSINK L, et al. Sucrose-specific induction of anthocyanin biosynthesis in Arabidopsis requires the MYB75/PAP1 gene[J]. Plant Physiology, 2005, 139(4): 1840-1852.
[11] 王學奎. 植物生理生化實驗原理和技術[M]. 2版. 北京: 高等教育出版社, 2006: 134-136.
[12] AINSWORTH E A, GILLESPIE K M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent[J]. Nature Protocols, 2007, 2(4): 875-877.
[13] 劉念, 蒙大慶, 湯天澤, 等. 油菜硫代葡萄糖苷檢測技術研究進展[J].湖北農業科學, 2011, 50(7): 1301-1304.
[14] 李培武, 周海燕. 油菜硫代葡萄糖苷檢測技術研究進展[J]. 中國油料作物學報, 2008, 30(1): 127-131.
[15] 曹建康, 姜微波, 趙玉梅. 果蔬采后生理生化試驗指導[M]. 北京: 中國輕工業出版社, 2011: 142-144.
[16] 張愛琴, 朱鐘. 植物生理學實驗[M]. 北京: 北京大學出版社, 1990.
[17] GUO Rongfang, YUAN Gaofeng, WANG Qiaomei. Effect of sucrose and mannitol on the accumulation of health-promoting compounds and the activity of metabolic enzymes in broccoli sprouts[J]. Scientia Horticulturae, 2011, 128(3): 159-165.
[18] SOLFANELLI C, POGGI A, LORETI E, et al. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis[J]. Plant Physiology, 2006, 140(2): 637-646.
[19] TO J P C, REITER W D, GIBSON S I. Chloroplast biogenesis by Arabidopsis seedlings is impaired in the presence of exogenous glucose[J]. Physiologia Plantarum, 2003, 118(3): 456-463.
[20] 劉麗萍. 外源蔗糖對蕎麥種子萌發、幼苗生長和黃酮積累及根系耐鹽性的影響[D]. 南京: 南京農業大學, 2011.
[21] KOPSELL D A, BARICKMAN T C, SAMS C E, et al. Influence of nitrogen and sulfur on biomass production and carotenoid and glucosinolate concentrations in watercress (Nasturtium offi cinale R. Br.)[J]. Journal of Agricultural and Food Chemistry, 2007, 55: 10628-10634.
[22] REIF C, ARRIGONI E, SCHARER H, et al. Carotenoid database of commonly eaten Swiss vegetables and their estimated contribution to carotenoid intake[J]. Journal of Food Composition and Analysis, 2013,29(1): 64-72.
[23] GIORGI A, MINGOZZI M, MADEO M, et al. Effect of nitrogen starvation on the phenolic metabolism and antioxidant properties of yarrow (Achillea collina Becker ex Rchb.)[J]. Food Chemistry, 2009,114(1): 204-211.
[24] KIM H J, CHEN Feng, WANG Xi, et al. Effect of methyl jasmonate on phenolics, isothiocyanate, and metabolic enzymes in radish sprout (Raphanus sativus L.)[J]. Journal of Agricultural and Food Chemistry,2006, 54(19): 7263-7269.
[25] REYES L F, CISNEROS-ZEVALLOS L. Wounding stress increases the phenolic content and antioxidant capacity of purple-flesh potatoes (Solanum tuberosum L.)[J]. Journal of Agricultural and Food Chemistry, 2003, 51(18): 5296-5300.
[26] GUERRERO-BELTRAN C E, CALDERON-OLIVER M, PEDRAZACHAVERRI J, et al. Protective effect of sulforaphane against oxidative stress: recent advances[J]. Exper imental and Toxicologic Pathology,2012, 64(5): 503-508.
[27] BODNARYK R P. Effects of wounding on glucosinolates in the cotyledons of oilseed rape and mustard[J]. Phytochemistry, 1992,31(8): 2671-2677.
[28] JIA Chengguo, XU Chaojiong, WEI Jia, et al. Effect of modified atmosphere packaging on visual quality and glucosinolates of broccoli florets[J]. Food Chemistry, 2009, 114(1): 28-37.
[29] RASK L, ANDREASSON E, EKBOM B, et al. Myrosinase: gene family evolution and herbivore defense in Brassicaceae[J]. Plant Molecular Biology, 2000, 42(1): 93-113.
[30] CHAMPOLIVIER L, MERRIEN A. Effect s of water stress applied at different growth stages to Brassica napus L. var. oleifera on yield,yield components and seed quality[J]. European Journal of Agronomy,199 6, 5(3/4): 153-160.
[31] SCHONHOF I, KLARING H P, KRUMBEIN A, et al. Interaction between atmospheric CO2and glucosinolates in broccoli[J]. Journal of Chemical Ecology, 2007, 33(1): 105-114.
[32] YUAN Youxi, CHIU Liwei, LI Li. Transcriptional regulation of anthocyanin biosynthesis in red cabbage[J]. Planta, 2009, 230(6): 1141-1153.
[33] JAHANGIR M, ABDEL-FARID I B, KIM H K, et al. Healthy an d unhealthy plants: the effect of stress on the meta bolism of Brassicaceae[J]. Environmental and Experimental Botany, 2009,67(1): 23-33.
[34] 李小剛, 宋云鵬, 王靜, 等. 外源蔗糖對NO3—脅迫下黃瓜幼苗生長及抗氧化酶活性的影響[J]. 山東農業科學, 2011(7): 30-34.
[35] 賈曉琳. 外源糖在心里美蘿卜幼苗花青素代謝中的作用初探[D]. 新鄉: 河南師范大學, 2013.
Effect of Exogenous Sucrose on Quality and Metabolic Enzyme Activities of Radish Sprouts
ZHAO Xiaoguo, ZHU Yi*, LUO Yunbo
(College of Food Science and Nutritional Engineering, China Agricultural University, Beijing 100083, China)
Radish (cv. Mantanghong) sprouts were adopted as the experimental material and sprayed with sucrose at different concentrations. The variations of physiological indexes (anthocyanin, chlorophyll, carotenoid, total phenolic,total glucosinolate, phenylalanine ammonia-lyase activity and peroxidase activity) of 3-, 5- and 7-day-old sprouts were investigated. Results showed that the contents of anthocyanin, chlorophyll, carotenoid, total phenolic and total glucosinolate were significantly (P < 0.05) increased by 0.4 and 0.6 g/100 mL sucrose treatment for 3-, 5- and 7-day-old sprouts, but not influenced (P > 0.05) by 0.2 g/100 mL sucrose. There was no difference between 0.4 and 0.6 g/100 mL sucrose treatment. Therefore, the nutritional value of radish sprouts could be increased by spraying 0.4 g/100 mL sucrose.
radish; sprouts; sucrose; quality; metabolic enzyme
TS255.1
A
1002-6630(2015)09-0007-05
10.7506/spkx1002-6630-201509002
2014-06-30
國家自然科學基金青年科學基金項目(31101263)
趙曉幗(1989—),女,碩士研究生,研究方向為食品科學。E-mail:zhaoxg_runner@163.com
*通信作者:朱毅(1973—),女,副教授,博士,研究方向為果蔬采后保鮮與生物活性物質功效。E-mail:zhuyi@cau.edu.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
