乳酸菌基因表達載體及其應用研究進展
2015-12-13 05:41:40崔月倩王菁蕊王艷萍
食品科學 2015年9期
關鍵詞:系統
崔月倩,王菁蕊,王艷萍
(天津科技大學食品工程與生物技術學院,天津 300457)
乳酸菌基因表達載體及其應用研究進展
崔月倩,王菁蕊,王艷萍*>
(天津科技大學食品工程與生物技術學院,天津 300457)
近年來隨著乳酸菌的功能特性和應用研究的深入,利用分子手段研究乳酸菌的功能基因已經成為最新 研究熱點。研究者構建眾多不同特性的基 因表達載體,本文從載體使用的安全性角度對已經報道的載體進行總結,并介紹各類基因表達載體在發酵產胞外多糖、酶制劑、疫苗制備等方面的應用。
乳酸菌;非食品級基因表達載體;食品級基因表達載體;應用
乳酸菌屬革蘭氏陽性、兼性厭氧菌,是一類可以利用發酵碳水化合物產生乳酸的一類菌的總稱,包括腸球菌、乳桿菌、乳球菌、明串珠菌、片球菌、鏈球菌和雙歧桿菌等眾多種屬[1-2],被廣泛地應用于食品發酵、醫藥生產、保健藥物及飼料添加劑等工業。對乳酸菌的研究已經由傳統的菌株篩選、功能特性 及其應用,深入到各種功能的分子機制以及乳酸菌作為食品及生物制品的載體等方面,乳酸菌的分子生物學成為當前的研究熱點之一。早在20世紀80年代初,乳酸菌的遺傳特性開始被國內外專家學者所關注。隨后相繼研發了一系列的基因表達載體和模式菌株。
近年來,研究者從不同的應用角度構建了大量的乳酸菌基因表達載體。就乳酸菌的應用特性而言,乳酸菌表達載體主要分為兩類:非食品級乳酸菌表達載體以及食品級乳酸菌表達載體,這也是乳酸菌非常獨特的區別于其他模式生物的特點之一。本文就乳酸菌中基因表達載體及其應用進行綜述。
1 非食品級基因表達載體
20世紀80年代初,乳酸菌被國內外專家學者關注,展開了乳酸菌遺傳系統的研究,這對乳酸菌的工業化應用以及對菌株的改造有重要意義。在乳酸菌的分子遺傳特性研究過程中,為了選擇合適的轉化子,對特定菌株進行篩選,通常將一個或多個編碼抗性篩選標記的基因與載體進行連接,從而達到篩選重組菌株的目的。其中常見的抗性篩選標記基因有紅霉素、氯霉素等抗性基因。
1.1 帶有紅霉素抗性標記的載體
紅霉素是糖多孢紅霉菌(Saccharopolyspora erythraea)合成的次級代謝產物,目前廣泛應用于臨床及作為分子生物學的篩選標記,其對革蘭氏陽性菌以及革蘭氏陰性菌均有良好的抑菌作用。自從Thompson等[3]克隆出紅霉素抗性基因ermE后,該基因廣泛應用于基因表達載體的構建(表1)。例如:van de Guchte等[4]構建的3.6 kb大小的大腸桿菌-乳酸菌多宿主穿梭載體pMG36e,攜帶來自于質粒pE194的紅霉素抗性基因EryR、大腸桿菌的強啟動子P32、pWV01質粒的復制子以及多克隆位點,以及乳酸乳球菌乳脂亞種蛋白酶基因(prtP)的轉錄終止子,目前是乳酸菌中最常用的載體之一。由Kleerebezem[5]及van de Guchte[4]等構建的乳酸菌-大腸桿菌多宿主穿梭載體pNZ9520、pNZ9530,由低拷貝質粒pAMβ1和pIL253衍生,以ermE作為篩選標記,攜帶來自pAMβ1的rep啟動子以及pIL253的nisR和nisK基因,是近年來乳鏈球菌素調控表達系統(nisin controlled gene expression system,NICE)中乳酸乳球菌基因表達研究的代表。此外,還有一系列攜帶了紅霉素篩選標記的乳酸菌表達載體。如,Bryan等[6]構建的pMSP3535載體,van Kranenburg等[7]構建的pNZ4055表達載體等。
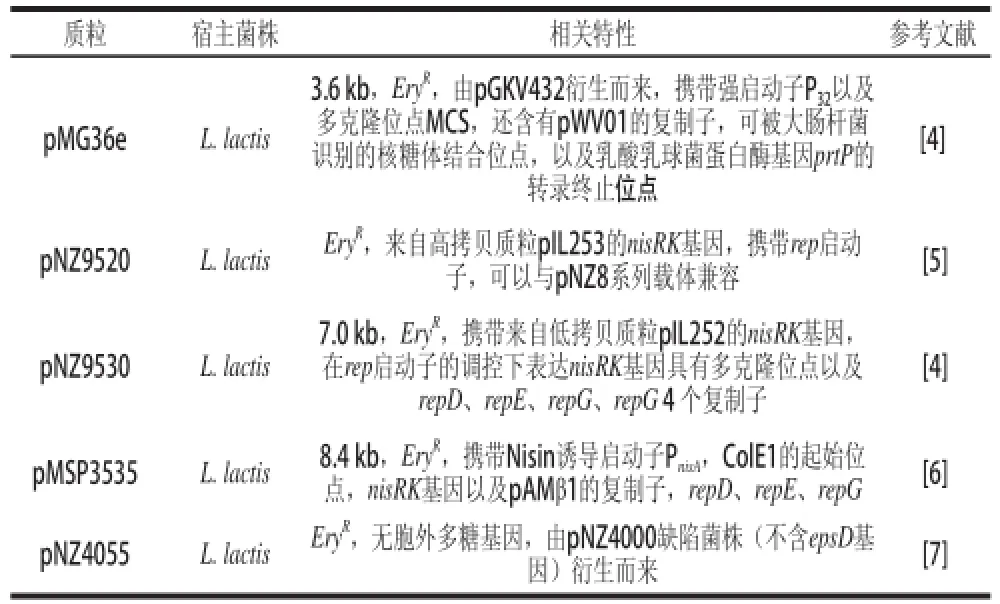
表1 攜帶紅霉素基因EryR篩選標記的常見乳酸菌基因表達載體Table 1 Gene expression withEErryyRin lactic acid bacteriaa
1.2 帶有氯霉素抗性標記的載體
由于氯霉素可以增加低拷貝質粒的拷貝數,提高質粒的得率,因此在大多數構建的乳酸菌表達載體中,均選用了氯霉素作為抗性篩選標記。常用的pNZ系列質粒大都采用了氯霉素抗性篩選標記(表2)。例如:由pNZ123以及pNZ273質粒衍生的以Lactococcus lactis NZ9000/NZ9100為宿主菌株的pNZ8008載體[8],含有CmR抗性基因,攜帶有gusA基因,來源于pSH71質粒的復制子。由此基礎質粒構建的pNZ8037[8]及pNZ8048[9]載體,也是標準的乳酸乳球菌表達載體,編碼CmR抗性基因,攜帶cat基因,復制子來源于pSH71質粒,并攜帶有Nisin誘導的啟動子PnisA,可以在大腸桿菌和乳酸菌中進行表達。以pNZ8048為基礎載體構建的pNZ8148[10]及pNZ8150[10]載體均攜帶了CmR抗性基因,克服了pNZ8048載體低水平 表達的缺點,在乳酸菌內高效表達。除上述載體外,還有其他由基礎載體構建的克隆表達載體,例如:Platteeuw等[11]構建的pNZ2102、pNZ2103載體也攜帶了氯霉素抗性篩選標記,陳家锃等[12]以pPG612為基礎載體構建的SlpA-612載體。
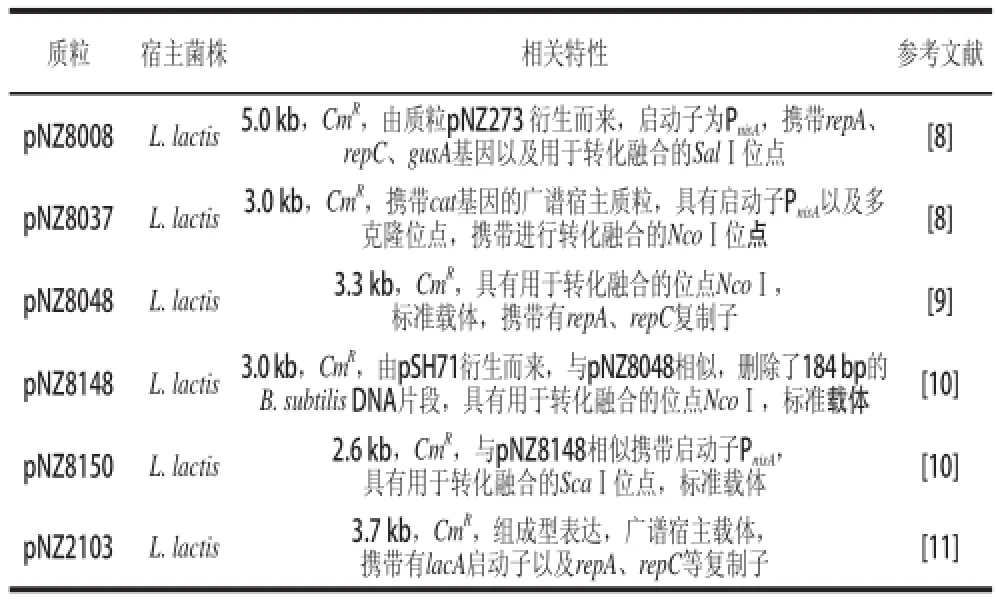
表2 以氯霉素為抗性篩選標記(編碼CmR抗性基因)的常見乳酸菌基因表達載體Table 2 Gene expression with CCmmRin lactic acid bacteriaa
乳酸菌表達載體除了以紅霉素和氯霉素作為篩選標記外,卡那霉素和氨芐青霉素等抗生素也可被用來作為篩選標記。
2 食品級表達載體
目前,分子克隆的載體系統大多是以抗生素抗性作為外源基因穩定表達的選擇標記,而抗性基因容易向環境中漂移擴散,對環境生態系統造成破壞,且不能直接用于人和動物,對生物體易造成不可估量的危害。乳酸菌作為益生菌的主要來源,一般認為是安全的食品級微生物(generally recognized as safe,GRAS),它的應用也是和食品密切相關。隨著人們對食品安全問題日益重視,眾多學者一直致力于高 效、無毒副作用的乳酸菌表達載體的研究。在食品中應用的基因表達載體,需要滿足以下條件:1)轉化載體的宿主菌必須是食品級微生物。食品級微生物,遺傳特性清楚,且能穩定遺傳。乳酸乳球菌和植物乳桿菌等食品級乳酸菌由于其安全無毒的特性,已經在食品工業中得到廣泛的應用。2)乳酸菌基因載體必須是食品級。為了進行篩選實驗,構建的眾多載體均帶有抗性基因標記,在食品中存在抗性基因標記會對人體和環境帶來的一定的安全隱患,所以在食品中必須選用含有食品級篩選標記的載體來進行選擇。3)乳酸菌使用的誘導物必須是食品級。在乳酸菌的克隆表達過程中,需要一定的誘導物。在食品表達系統中,要求誘導物必須是食品級,為人類可食用,例如:乳糖、蔗糖、Nisin等。
2.1 食品級表達系統
在食品級乳酸菌 中已經形成了一系列適合乳酸菌的基因表達系統,例如:糖誘導表達系統,噬菌體Φ31爆發式誘導的表達系統、乳鏈球菌素調控表達系統、pH值調控表達系統[13]等。
2.1.1 糖誘導表達系統
乳酸菌基因組中含有糖代謝相關基因簇,在以各種糖類為碳源的培養基上生長。在乳酸菌的糖誘導表達系統中,主要應用的是糖類選擇標記。糖類作為食品級誘導物,可以作為非抗生素抗性的選擇標記進行應用。大多數表達系統使用乳糖作為選擇標記來誘導基因表達。在乳糖選擇系統中,將糖類篩選標記進行克隆,構建食品級載體(表3)。例如:以L. lactis NZ3900為宿主菌的食品級乳酸菌表達載體pNZ8149[8],大小為2.5 kb,在NcoⅠ位點后攜帶有Nisin誘導的啟動子PnisA,并含有lacF基因,以乳糖為選擇標記,通過乳糖進行篩選。以L. casei為宿主菌的食品級質粒pLEB590[14]和pLEB600[15]均由L. lactis DNA構成,攜帶pSH71復制子以及repA基因,分別攜帶P45以及PpepR啟動子,且含有lacG基因,利用乳糖選擇標記進行篩選,此外,pLEB590還攜帶Nisin結構基因-nisI基因。
此外,還有木糖、蔗糖等選擇標記。以D-木糖作為選擇性標記的大腸桿菌-乳酸桿菌穿梭載體pLP3537[16]進行了戊糖乳桿菌染色體上的木糖還原酶基因簇xyl的克隆與表達。此乳酸菌表達載體由L. pentosus MD353中1.7 kb和2.3 kb的質粒構成,攜帶有XylR、XylA以及XylB以及來自于pUC19的復制子。Leenho uts等[17]利用乳酸菌的整合構建了以蔗糖為選擇標記的食品級表達系統,將編碼蔗糖和蔗糖水解酶的基因scrA/scrB克隆進乳球菌質粒pWV01中,利用此質粒的拷貝(Ori+)以及L. lactis DNA在L. lactis LL108及L. lactis LL102菌種中進行構建,成功構建了以蔗糖為篩選標記的乳酸菌表達載體pINT124、pINT125。Mahmoud等[18]利用此表達載體在乳酸乳球菌中表達了細菌素基因pctA。
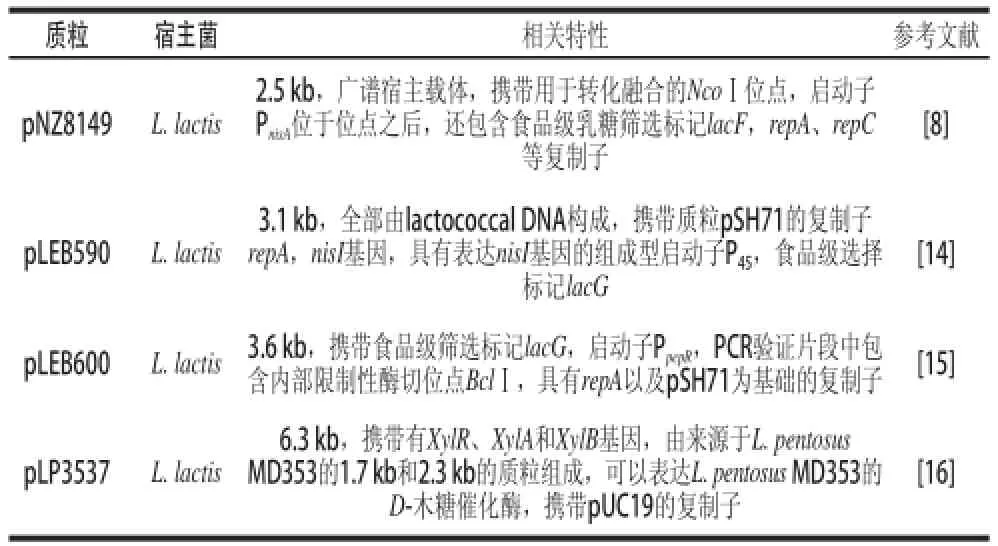
表3 糖誘導表達系統應用的常見乳酸菌基因表達載體Table 3 Gene expression vectors in sugar inducible expression system
2.1.2 乳鏈球菌素調控表達系統
在乳鏈球菌素調控 表達系統中,應用了細菌素抗性選擇標記。細菌素是細菌在代謝過程中產生的一種具有抗菌特性的蛋白質,菌體本身對細菌素具有免疫性[19]。常用的食品級細菌素為乳鏈球菌素,又名乳鏈菌肽(Nisin),此食品級誘導物控制的基因表達系統為NICE基因表達系統,是由de Ruyter等[8]構建的。它是以Nisin生物合成基因簇(包括結構基因nisA)的啟動子PnisA和雙組分調節系統基因nisRK為基礎,由Nisin誘導而自我調節的系統。Nisin可以在含有nisRK基因的宿主菌中高效誘導表達,在PnisA后插入外源基因,研究證明利用Nisin控制的基因表達(nisin controlled gene expression system,NICE)系統在乳酸菌中表達異源蛋白,誘導效率可超過1 000 倍以上[9]。許多乳酸菌菌株因含有Nisin抗性基因,構建基因載體便可以此基因為抗性選擇標記。目前,已構建了大量含有Nisin抗性基因的載體(見表2、3)。van de Guchte等[4]構建的含有EryR的多宿主菌株表達載體pNZ9530運載nisR和nisK基因,在乳鏈菌肽誘導下表達雙質粒系統。此外,由pNZ123、 pNZ273和pSH71等基礎載體演變而來的含有CmR的乳酸菌表達載體pNZ8008[8]、pNZ8037[8]、pNZ8048[10]、pNZ8148[10]和pNZ8150[10]等均含有nisA基因,含有Nisin誘導的啟動子PnisA,在Nisin的誘導下進行表達。de Ruyter等[8]構建的大小為2.5 kb的pNZ8149食品級表達載體以Lactococcus lactis NZ3900為宿主菌,在NcoⅠ位點含有一個nisA基因,以及由Nisin誘導的PnisA,并運載 repA、repC基因及lacF基因。L. lactisNZ3900/ pNZ8149系統的誘導劑、篩選物和宿主都是食品級,符合美國食品藥品監督管理局(U.S. Food and Drug Administration,FDA)安全標準,該系統可作為理想的異源蛋白表達系統,此系統已經表達了靈芝免疫調節蛋白編碼基因LZ-8[20]。此外,Takala等[14]構建的以乳酸乳球菌MG1614為宿主菌的pLEB590食品級表達載體,運載nisI基因,由乳球菌DNA,pSH71復制子和用于nisI表達的組成型啟動子P45組成。上述載體均已經成功的應用在NICE基因表達系統中。
2.1.3 噬菌體Φ31爆發式誘導的表達系統
在乳酸菌的噬菌體誘導表達系統中,通過Φ31侵染后進行誘導表達。此系統中應用的表達載體pTRK391,是由LacZ基因以及Φ31啟動子P8625構成,構建載體后侵染乳酸菌菌株,構建噬菌體誘導系統,啟動蛋白的高效表達,但是會促進乳酸菌宿主菌的裂解,因此噬菌體表達系統并沒有被廣泛應用[16-21]。
2.1.4 pH值調控表達系統
Madsen等[22]通過研究發現了受pH值 調節的啟動子P170,構建了表達載體pAMJ529、pAMJ536和pAMJ547,轉 化后發現,當pH值為5.5時,菌株能夠生長,當上調至7.0時表達受到抑制,停止生長。通過調節不同的pH值,進行調控系統的誘導表達。
2.2 營養缺陷篩選標記
此種選擇標記是將某些表型基因進行突變或缺失,整合到質粒中,再整合入互補的表型,進行篩選(表4)。Dickely等[23]構建的乳酸菌表達載體pFG1,以赭石型突變抑制基因supB作為選擇標記,進行克隆表達。Sorensen等[24]等用琥珀型突變抑制基因supD構建了可抑制嘧啶營養缺陷的載體pFG200載體。此外,王春鳳[25]構建的以胸苷酸合成酶基因thyA作為篩選標記的非抗性重組質粒表達載體pW425t,以及孫強正等[26]構建的食品級分泌型表達載體pSQZ,以thyA為選擇標記,表達出有保護活性的蛋白。質粒pW425t和pSQZ是由以乳酸乳球菌MBP71為宿主菌的以thyA為選擇標記的質粒pSH91構建而來。

表4 以營養缺陷作為篩選標記的常見的乳酸菌基因表達載體Table 4 Gene expression with nutrient deficiency in lactic acid bacteria
3 乳酸菌表達載體的應用
乳酸菌是人體內必不可少且具有重要生理功能的菌群,廣泛存在于人體腸道中,調節著人體的腸道健康,與人們的健康長壽有著直接關系。乳酸菌作為安全級食品菌株,通過對其深入研究,應用乳酸菌基因表達載體構建重組菌株,及其產生的大量酶、多肽、中間代謝物等表達產物,可以應用于食品、醫藥、保健業和工業等領域,有巨大的應用前景和潛在的商業價值。乳酸菌表達載體的廣泛應用,提高了酶制劑、胞外多糖等代謝物質的產量,對食品、醫藥和工業等領域的發展具有一定的促進作用。
3.1 在疫苗制備方面的應用
近年來,對疫苗傳遞系統的研究過程中,人們期待找到一種能夠在體內較長時間存在,而對機體本身既安全又能產生持續免疫力的傳遞載體。乳酸菌廣泛存在于人類與動物腸道中,促使機體產生特異性或非特異性的免疫應答反應。近年來隨著乳酸菌質粒載體系統、電穿孔轉化技術、食品級載體的發展,以乳酸菌作為研制載體疫苗成為可能[27]。目前已經有利用乳酸菌研究研制疫苗的報道。Kajikawa等[28]利用表達載體pIGM2J在Lactobacillus casei中進行轉化,成功誘發細胞免疫因子的分泌,促進了乳酸菌疫苗的成功應用。Sung等[29]將構建的載體pKV-Pald-PgsA-Amylase在乳酸菌中表達人乳頭瘤病毒(human papilloma virus,HPV)抗原蛋白,構建重 組菌株,為制備子宮頸癌疫苗,治愈子宮頸癌提供了可能。此外,曲英敏等[30]利用大腸桿菌-乳酸菌穿梭載體pW425et與腦膜炎奈瑟氏菌NspA基因進行重組,構建表達載體,進行蛋白表達,表達蛋白能夠與腦膜炎奈瑟氏菌患者體內的IgG特異性結合,推動了研制抵抗腦膜炎奈瑟氏菌的乳酸菌免疫作用疫苗的發展。郭衍冰等[31]也利用此載體對新城疫病毒基因(HN)進行了表達,為制備病毒口服疫苗提供了技術保障。夫廈玲[32]利用pKV-Pald-PgsA-E7(Rb)表達載體在乳酸菌內表達,乳酸菌在其表面上表達HPV抗原蛋白E7(Rb),然后將其作為疫苗直接使用,通過在小鼠模型實驗,產生黏膜免疫應答,對患有宮頸癌的小鼠具有一定的效果。
3.2 在酶制劑制備中的應用
近年來,通過將乳酸菌酶基因與質粒連接構建表達載體,在乳酸菌或外源菌株中進行大量表達,成功制備各類酶制劑。van de Guchte等[4]在pMG36e載體構建成功后,表達了溶菌酶基因,對溶菌酶的大量表達與制備提供了可行性。寇田田[33]在此質粒的基礎上進行改造,構建了以紅色熒光蛋白基因為標記的乳酸菌融合表達系統pMG36e-dsred2,對α-淀粉酶基因 進行了表達,能夠產生淀粉酶從而分解淀粉。Wang Lamei等[34]利用載體pNZ8149對硬脂酰-輔酶A脫氫酶基因scd1進行表達,對功能性酶制劑的制備提供了美好前景。Sorensen等[24]構建的食品級表達載體pFG200,在干酪乳桿菌中進行了β-葡萄糖苷酸酶基因(gusA)的表達,以及進行了蛋白酶基因pepN在工業乳酸乳球菌的克隆表達。此外,唐麗杰等[35]還利用了pPG612載體表達了豬乳鐵蛋白基因,為重組乳酸菌抗菌制劑的制備奠定了基礎。Li Bin等[36]利用載體pNZ8148將膽堿水解酶(bile salt hydrolase,BSH)基因轉入乳球菌標準菌株NZ9000中,為制備降膽固醇的酶制劑提供了實驗和理論基礎。
3.3 在代謝產物調控方面的應用
有不同的發酵特性及代謝產物,例如產胞外多糖。由于乳酸菌的安全性,因此其生產的胞外多糖可以應用于食品工業領域,且胞外多糖還具有抗腫瘤,免疫作用,對生物體具有良好的益生作用[37]。由于乳酸菌菌株產的胞外多糖產量較小,通過分子生物學手段使其高產胞外多糖,滿足食品工業的需求。Haywood等[38]將多糖代謝關鍵酶——α-磷酸葡萄糖變位酶基因pgm利用pT1NX表達載體電轉化到Lactobacillus casei BL310中,成功使胞外多糖產量增加了172%。van Kranenburg等[7]構建的pNZ4055表達載體在產胞外多糖乳酸球菌中過量地表達eps-D基因(代謝關鍵酶——引導糖基轉移酶),使胞外多糖產量提高15%。除胞外多糖外,還有乳酸以及類黃酮等代謝物的生產。寇田田[33]在質粒pMG36e的基礎上構建的pMG36e-dsred2基因工程菌,實現了一步法生產乳酸,提高了乳酸的產量。Schümann等[39]利用表達載體pSIP409將蘋果酸乳酸酶基因(mle)克隆到Lactobacillus plantarum WCFS1中,發現重組菌加速了乳酸的發酵。Liu Hongyu等[40]利用載體pNZ8149將大豆查爾酮異構酶基因(CHI1A)進行克隆表達,為生產類黃酮奠定基礎。
4 結 語
乳酸菌作為益生菌,其應用前景巨大,早已涵蓋食品、醫療、養殖、環保、新材等多個領域,對有效抵抗腸道感染、抗腹瀉、調節胃腸道功能,減輕腹脹、腹瀉、便秘等起到了重要作用。但因其遺傳背景不清楚,應用受到限制,通過抗性篩選載體對乳酸菌功能基因進行研究,進一步清晰其調控機理,在乳酸菌遺傳機制的調控以及對乳酸菌的改造方面具有推動作用。乳酸菌抗性篩選載體構建了大量的基因工程菌株,促進了乳酸菌工業化的應用進展,通過基因的超表達,多肽、酶、多糖、細菌素等代謝物質的產量提高,為制備酶制劑、乳酸菌口服疫苗提供了理論基礎和實驗依據。
然而,乳酸菌抗性篩選載體在應用的同時,因其攜帶的抗性篩選基因,對環境以及人體有害,雖然達到了篩選的目的,但在食品、疫苗等生產方面的應用受到了一定程度的限制。因此,需要利用食品級、安全性高的篩選標記代替抗性標記,構建食品級乳酸菌載體。食品級乳酸菌載體可以有效地應用于乳酸菌食品、口服疫苗生產中,其安全、穩定的特性為人們所重視,為人們所應用。
綜上,乳酸菌表達載體的研究及應用,對推動乳酸菌的分子生物學研究起到關鍵作用。在乳酸菌構建的表達載體中,含有抗性篩選標記的表達載體雖不能應用在食品中,但對于研究乳酸菌的遺傳背景做出了巨大的貢獻。食品級的乳酸菌表達載體無毒副作用,對利用分子手段改造乳酸菌菌株發揮了安全保證作用。乳酸菌基因表達載體已經成功的應用于酶、多肽、中間代謝物等表達產物的生產,對食品、醫藥、保健業和工業等領域大發展起到極大的推動作用。在乳酸菌中還有許多隱蔽性質粒存在,其具有巨大的應用前景和潛在的應用價值,需要進一步的深入研究。隨著乳酸菌分子生物學的發展,對乳酸菌基因工程菌株的利用與研發將對乳酸菌的深入研究和應用發揮重要作用。
[1] 劉斌, 黃少磊, 劉彥民. 分子標記技術應用于乳酸菌分類鑒定的研究進展[J]. 中國微生態學雜志, 2012, 24(7): 670-673.
[2] KHALISANNI K. An overview of lactic acid bacteria[J]. International Journal of Biosciences, 2011, 1(3): 1-13.
[3] THOMPSON C J, KIESER T, WARD J M, et al. Physical analysis of antibiotic-resistance genes from Streptomyces and their use in vector construction[J]. Gene, 1982, 20(1): 51-62.
[4] van de GUCHTE M, van der VOSSEN J, KOK J, et al. Construction of a lactococcal expression vector: expression of hen egg white lysozyme in Lactococcus lactis subsp. lactis[J]. Applied and Environmental Microbiology, 1989, 55(1): 224-228.
[5] KLEEREBEZEM M, BEERTHUYZEN M, VAUGHAN E E, et al. Controlled gene expression systems for lactic acid bacteria transferable nisin-inducible expression cassettes for Lactococcus, Leuconostoc, and Lactobacillus spp.[J]. Applied and Environmental Microbiology, 1997,63(11): 4581-4584.
[6] BRYAN E M, BAE T, KLEEREBEZEM M. Improved vectors for nisin-controlled expression in gram-positive bacteria[J]. Plasmid,2000, 42(2): 183-190.
[7] van KRANENBURG R, vos HARMJAN R, van SWAMI I, et al. Functional analysis of glycosyl transferase genes from Lactococcus lactis and other gram-positive cocci: complementation, expression,and diversity[J]. Journal of Bacteriology, 1999, 181(20): 6347-6353.
[8] de RUYTER P G, KUIPERS O P, de vos WILLEM M. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin[J]. Applied and Environmental Microbiology, 1996,62(10): 3662-3667.
[9] KUIPERS O P, de RUYTER P G, KLEEREBEZEM M, et al. Quorum sensing-controlled gene expression in lactic acid bacteria[J]. Journal of Bacteriology, 1998, 64(1): 15-21.
[10] MIERAU I, KLEEREBEZEM M. 10 years of the nisin-controlled gene expression system (NICE) in Lactococcus lactis[J]. Applied Microbiology and Biotechnology, 2005, 68(6): 705-717.
[11] PLATTEEUW C, van ALEN-BOERRIGTER I, van SCHALKWIJK S. Food-grade cloning and expression system for La ctococcus lactis[J]. Applied and Environmental Microbiology, 1996, 62(3): 1008-1013.
[12] 陳家锃, 崔紅玉, 田志軍. 乳酸桿菌組成型啟動子探測載體pgateway-612的構建[J]. 中國預防獸醫學報, 2011, 33(11): 849-853.
[13] 王瑩, 胡桂學. 食品級乳酸菌表達系統的研究進展[J]. 中國畜牧獸醫, 2010, 37(7): 80-83.
[14] TAKALA T M, SARIS P E J. A food-grade cloning vector for lactic acid bacteria based on the nisin immunity gene nisI[J]. Applied Microbiology and Bi otechnology, 2002, 59(4): 467-471.
[15] TAKALA T M. Nisin immunity and food-grade transformation in lactic acid bacteria[D]. Finland: University of Helsinki, 2005.
[16] POSNO M, HEUVELMANS P T, van GIEZEN M J, et al. Complementation of the inability of Lactobacillus stains to utilize D-xylose with D-xylose catabolism-encoding genes of Lactobacillus pentosus[J]. Applied and Environmental Microbiology, 1991, 57(9): 2764-2766.
[17] LEENHOUTS K, BOLHUIS A, VENEMA G, et al. Construction of a food-grade multiple-copy integration system for Lactococcus lactis[J]. App l ied Microbiology and Biotechnology, 1998, 49(1): 417-423.
[18] MAHMOUD K T, SAMEH E M. Heterologous expression of pctA gene expressing propionicin T1 by some lactic acid bacterial strains using pINT125[R]. Netherlands: Alexandria University, 2011.
[19] 張小美, 樓秀玉, 顧青. 1 株 產 細菌素乳酸菌的鑒定和細菌素的分離純化[J]. 中國食品學報, 2013, 13(12): 181-187.
[20] 曲寧寧, 陳萍, 孫鑫澤. 靈芝lz-8基因乳酸菌表達載體的構建與表達及免疫特性[J]. 吉林農業大學學報, 2013, 35(4): 411-415.
[21] WALKER S A, KLAENHAMMER T R. Molecular characterization of a phage inducible middle promoter and its transcriptional activator from the lactococcal bacteriophage Φ31[J]. Journal of Bacteriology,1998, 180(4): 921-930.
[22] MADSEN S M, ARNAU J, VRANG A, et al. Molecular chara cterization of the pH-inducible and growth phase-dependent promoter P170 of Lactococcus lactis[J]. Molecular Microbiology,1999, 32(1): 75-87.
[23] DICKELY F, NILSSON D, HANSEN B E, et al. Isolation of Lactococcus lactis nonsense suppressors and construction of a food-grade cloning vector[J]. Mole cular Microbiology, 1995, 15(5): 839-847.
[24] SORENSEN K I, LARSEN R, KIBENICH A, et al. A food-grade cloning system for industrial strains of Lactococcus lactis[J]. Applied and Environmental Microbiology, 2000, 66(4): 1253-1258.
[25] 王春鳳. 共生乳酸桿菌非抗性表達載體的構建及柔嫩艾美耳球蟲SO7基因的表達[D]. 北京: 中國農業大學, 2001.
[26] 孫強正, 熊衍文, 葉長蕓. 食品級分泌表達載體的構建及報告蛋白在乳酸乳球菌中的表達[J]. 微生物學報, 2008, 48(3): 293-298.
[27] 孟慶峰, 徐展, 王偉利. 活載體疫苗的研究進展[J]. 黑龍江畜牧獸醫,2013(10): 28-31.
[28] KAJIKAWA A, MASUDA K, KATOH M, et al. Adjuvant effects for oral immunization provided by recombinant Lactobacillus casei secreting biologically active murine interleukin-1βΔ[J]. American Society for Micro biology, 2010, 17(1): 43-48.
[29] SUNG M H, POO H Y, LEE I H, et al. Stable constitutively high expression vector for preparing HPV vaccine and recombinant lactic acid bacteria transformed thereby: US, 8685721[P]. 2014-04-01.
[30] 曲英敏, 李艷艷, 李景梅. 腦膜炎奈瑟氏菌NspA基因重組乳酸菌表達載體的構建及原核表達[J]. 吉林農業大學學報, 2011, 33(4): 439-442.
[31] 郭衍冰, 胡靜濤, 于丹. 重組NDVHN 基因乳酸菌穿梭表達載體的構建及在大腸桿菌中的表達[J]. 吉林農業大學學報, 2013, 35(2): 184-187.
[32] 夫廈玲. 用于制備HPV疫苗和由其轉化的重組乳酸菌的穩定組成型高表達載體: 中國, 000126[P]. 2010-01-08.
[33] 寇田田. 乳酸菌融合表達載體的構建及α-淀粉酶基因的克隆與表達[D].保定: 河北農業大學, 2013.
[34] WANG Lamei, LI Shili, GOU Kemian, et al. Heterologous expression of stearoyl-CoA desaturase-1 in Lactococcus lactis NZ3900[J]. Chinese Journal of Biotechnology, 2012, 28(9): 1106-1117.
[35] 唐麗杰, 哈卓, 趙麗麗. 豬乳鐵蛋白基因的克隆及其重組乳酸菌表達系統構建[J]. 東北農業大學學報, 2010, 41(3): 79-84.
[36] LI Bin, JIANG Yujun. Cloning of bile salt hydrolase gene and its expression in lactic acid b a cteria[J]. Journal of Northeast Agricultural University(English Edition), 2011, 18(2): 48-53.
[37] 王艷萍, 李超, AHMED Z. 一株馬乳酒樣乳桿菌胞外多糖的理化性質[J]. 食品與發酵工業, 2011, 37(9): 59-63.
[38] SANF?LIX-HAYWOOD N, COLL-MARQU?S J M, YEBRA M J. Role of α-phosphoglucomutase and phosphoglucose isomerase activities at the branching point between sugar catabolism and anabolism in Lactobacillus casei[J]. Journal of Applied Microbiology,2011, 111(2): 433-442.
[39] SCH?MANN C, MICHLMAYR H, EDER R, et al. Heterologous expression of Oenococcus oeni malola ctic enzyme in Lactobacillus plantarum for improved malolactic fermentation[J]. AMB Express,2012, 2(1): 19. doi: 10.1186/2191-0855-2-19.
[40] LIU Hongyu, WANG Piyu, FU Yongping. Cloning of tap Ⅱ chalcone i somerases (CHI1A) gene and construction of Lactococcus lactis expression vector[J]. Journal of Agricultural Science and Technology,2010, 11(4): 44-46.
A Review of Research on Lactic Acid Bacteria Vectors for Gene Expression and Their Applications
CUI Yueqian, WANG Jingrui, WANG Yanping*
(College of Food Engineering and Biotechnology, Tianjin University of Science and Technology, Tianjin 300457, China)
In recent years, with the further functional study on the characteristics and applications of lactic acid bacteria, molecular biology of lactic acid bacteria is also of concern. Cloning and functional expression of lactic acid bacteria using molecular tools have become the latest hotspot. Researchers have found or built many gene expression vectors wit h different characteristics which lay the foundation for gene cloning and expression of lactic acid bacteria. Based on the characteristics of lactic acid bacteria, this paper summarizes the vectors which have been reported from the perspective of safety. And we also review the applications of gene expression vectors for the production of polysaccharides, enzyme and vaccine preparation, etc.
lactic acid bacteria; non-food-grade genetic expression vector; food-grade genetic expression vector; application
Q782
A
1002-6630(2015)09-0224-06
10.7506/spkx1002-6630-201509042
2014-08-10
國家自然科學基金面上項目(31171629);國家自然科學基金青年科學基金項目(31101218);中國博士后科學基金面上項目(2014M551029)
崔月倩(1989—),女,碩士研究生,研究方向為食品生物技術。E-mail:cuiyueqian@163.com
*通信作者:王艷萍(1962—),女,教授,博士,研究方向為食品生物技術。E-mail:ypwang@tust.edu.cn
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
