帽兒山林區紫椴群落物種多樣性、種間關系及對環境因子的響應
2015-12-16 08:11:28張東來
森林工程 2015年6期
關鍵詞:物種
張東來,張 玲
(1.黑龍江省林業科學院,哈爾濱150081;2.黑龍江省林業科學研究所,哈爾濱150081)
紫椴(Tilia amurensis Rupr.)不僅是我國東北地區重要的珍貴闊葉樹種,而且是重要的蜜源、城市綠化以及藥用樹種[1-2]。因為連年過量開采,致使天然紫椴種群無論從數量上還是從質量上都呈逐年下降與衰退的趨勢,面積和蓄積量銳減,現在已被列為國家Ⅱ級保護植物。
紫椴天然更新良好,但研究發現紫椴人工培育困難,主要是受霜凍害的制約,到目前為止,紫椴人工培育過程中幼樹易受霜凍危害的問題始終未得到有效解決,霜凍危害成為制約紫椴人工培育的瓶頸。加強紫椴人工林資源的培育研究,尤其是紫椴混交林的研究已迫在眉睫,這將對保護和擴大紫椴資源,使其永續利用具有深遠歷史意義和更重要的現實意義[3]。
本研究以帽兒山林區紫椴天然種群為研究對象,開展群落的物種多樣性、種間關系及其與環境關系的研究,掌握紫椴群落的組成、結構和功能與環境關系,為今后紫椴人工林培育提供技術支撐。
1 研究區概況
研究地點位于黑龍江省尚志市東北林業大學帽兒山實驗林場內,該林場位于北緯45°21'~45°25',東經 127°31'~127°34',林場平均海拔高度 300 m,最高海拔為805 m,屬長白山系張廣才嶺西部小嶺余脈,植被屬長白植物區系,是東北東部山區較典型的天然次生林區,本區土壤多為典型暗棕色森林土,有白漿化暗棕壤、草甸暗棕壤兩個亞類,土壤下層為巖石。地帶性頂極植被為闊葉紅松林,平均林齡50 a。主要闊葉樹種有紫椴(Tilia amurensis Rupr.)、水曲柳(Fraxinus mandshurica Rupr.)、暴馬丁香(Syringa amurensis Rupr.)春榆(Ulmus japonica(Rehd.)Sarg.)、忍冬(Lonicera naackii(Rupr.)Maxim)、色木(Acer mono Maxim)、衛矛(Euonymus alatus(Thunb.)Sieb.)等 13 種植物。本區屬大陸性氣候,年降雨量723.8 mm,無霜期120~140 d[4]。
2 研究方法
2.1 野外調查
采用典型樣地調查方法,2014年6月末,在帽兒山林場紫椴天然次生林內,沿海拔高度,在研究區域內設10個調查樣方,規格為20 m×20 m,并在每個喬木樣方內隨機設5個灌木樣方,規格為5 m×5 m,每個灌木樣方內隨機設5個草本樣方,規格為1 m×1 m。對樣方內的植物進行常規調查和記錄,并在每個樣地的4個角和中間取等量表土(0~20 cm)混合均勻,去除雜物,帶回、風干,待測[4-5]。
2.2 數據處理
多樣性指數的計算依據為重要值。以各層多樣性指數和為計算整個群落多樣性指數的依據,每種群落的多樣性指數以平均值為準[5]。
喬木重要值IV=(相對多度+相對頻度+相對顯著度)/3。
灌木和草本植物重要值IV=(相對高度+相對蓋度)/2。
群落多樣性的測度選用豐富度指數(S)、均勻度指數和物種多樣性指數。其計算公式如下:
物種豐富度D=S。
Shannon-Wiener index H'= ∑PilnPi。
Pielou指數:Jsw=(∑PilnPi)/lnS。
式中:Pi為種i的相對重要值;S為中i所在樣方的物種數,即豐富度指數。
2.3 成對物種間關聯的測度
Ochiai指數 OI=a/ [(a+b)(a+c)]1/2。
Dice指數 DI=2a/(2a+b+c)。
Jaccard指數 JI=a/(a+b+c)。
式中:a為2個種都存在的取樣數量;b為B種存在而A種不存在的取樣數量;c為A種存在而B種不存在的取樣數目;d為2個種都不存在取樣多數。
以上3個無中心指數的值域為0到1。其值越趨近于1,則表明該種對的正聯結越緊密[1,6]。
2.4 土壤樣品分析
用風干法測量土壤含水量,pH值用S-3C型酸度計測定,全氮含量用半微量凱氏定氮法測定;有機質采用沙浴外加熱法-重鉻酸鉀氧化容量法,全磷含量用硫酸高氯酸消煮-鉬銻抗比色測定,全鉀含量采用氫氟酸-高氯酸消煮-原子吸收測定。
3 研究結果
3.1 帽兒山林區紫椴群落多樣性指數與均勻度
帽兒山林區紫椴群落草本層物種多樣性豐富,達到19.01±1.29,其次是灌木層,見表1。帽兒山林區紫椴群落喬木層物種豐富度較低,喬木種類少,主要伴生種為山桃稠李(Podus maackiii)、水曲柳、五角槭、黃榆、蒙古櫟,胸徑平均值為22.2 cm,郁閉度為0.7,還有一些空間沒被占據,分布格局相對不均勻。隨著恢復時間逐漸增長,群落結構逐漸呈現出復雜化,均勻度指數也越來越大,反映出該群落越來越穩定,達到頂級群落。灌木層主要伴生種為暴馬丁香、金銀忍冬、五角槭、衛矛(Euonymus alatus)、毛榛子(Corylus heterophylla)、溲疏(Deutzia parviflora)。草本層以鈴蘭(Convallaria keiskei)、蚊子草(Filipendula palmate)、四花苔草(Carex quadriflora)、委陵菜(Potentilla chinensis)、玉竹(Polygonatum odoratum)、木賦(Equisetum hiemale)菊科、莎草科、毛茛科、薔薇科為主。香濃指數則是喬木層和草本層均顯著高于灌木層。
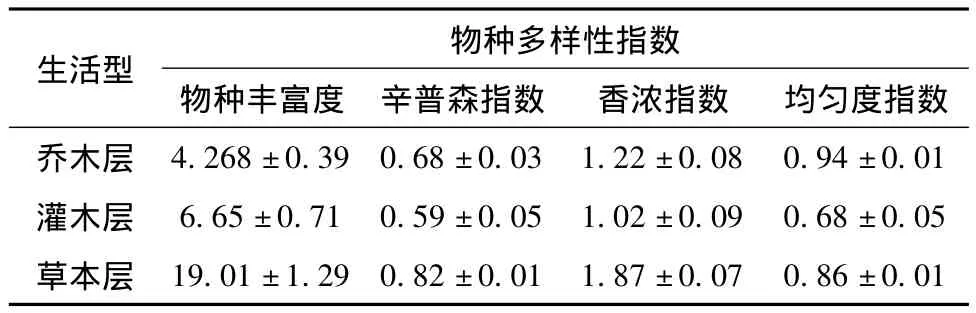
表1 帽兒山林區紫椴群落不同生長型物種多樣性指數和均勻度指數Tab.1 Diversity and evenness indexes ofdifferent growth forms of Tilia amurensis community in Maoer mountain
3.2 種對間關聯度分析
帽兒山林區紫椴群落有12個優勢種對,其中有7個種對是正關聯,占總種對數的58%,分別是紫椴-紅松、紫椴-五角槭、紫椴-茶條槭、紫椴-衛矛、紫椴-裂葉榆、紫椴-暴馬丁香、紫椴-糠椴(見表2)。紫椴種群是紅松種群的伴生種群,種群格局隨發育階段呈由強至弱的聚集分布,主要是因為紫椴幼苗期易受霜凍危害,盡管凍后也能萌發,但會造成無明顯主干,因此自然更新受到嚴重影響。

表2 帽兒山紫椴群落優勢種間關聯指數Tab.2 Interspecific relationship of dominant species in Tilia amurensis community
紫椴種對間極顯著正關聯有4對(0.01<P<0.05,ad>bc),紫椴-紅松、紫椴-衛矛、紫椴-暴馬丁香、紫椴-糠椴,顯著正關聯種對有1對(0.01<P<0.05,ad<bc)紫椴-裂葉榆,如圖1所示。說明該樹種對生境的要求和適應具有同一性或相似性,因此,優勢種在相對穩定的群落中可以共同利用群落的非限制性資源,形成顯著的正關聯。
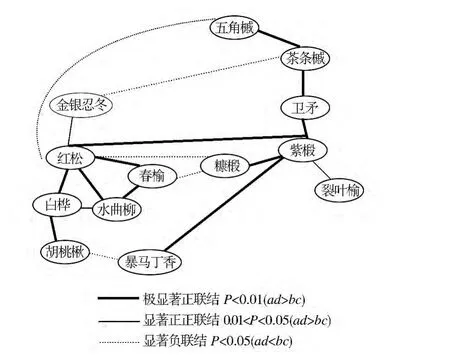
圖1 紫椴群落種群間聯結X2檢驗星座Fig.1 Constellation diagrams of X2-test of interspecific correlation in Tilia amurensis community
Ochiai指數指一個物種看作是另一個物種正常環境的一部分;Dice指數表示一個物種出現而另一物種也存在的概率。從表1可知,紫椴-五角槭、紫椴-衛矛、紫椴-糠椴,它們的OI值、DI值和JI值均在0.5,這是由于這些種群共同適應并占據著相近生境的緣故。OI值、DI值和JI值在0.3區間的種對有6對,分別是紫椴-白樺、紫椴-春榆、紫椴-水曲柳、紫椴-裂葉榆、紫椴-暴馬丁香、紫椴-胡桃楸,說明這些種對的聯結程度稍高,其它種對的聯結程度則較低。
X2值不僅能檢驗聯結的顯著性,而且可用以種群間聯結的測度,它在一定程度上衡量了種群間的相互關聯和植物對環境綜合生態因子反應的差異[7]。由圖1可以看出,如本區紫椴種群出現時,可能有五槭、茶條槭、裂葉榆、暴馬丁香、糖椴出現,這正是紫椴種群表現為頂級群落伴生種群的原因之一。
4 結果與討論
帽兒山林區紫椴群落垂直結構物種多樣性指數為:豐富度指數,草本層>灌木層>喬木層;辛普森指數,草本層>喬木層>灌木層。喬木層物種豐富度較低,草本層相對來說物種豐富并且分布相對較為均勻,灌木層物種盡管較為豐富。灌木層易受人為干擾(紅松球果采摘、火災)的影響,大部分物種個體分布不均勻,因此辛普森指數較低。該群落地帶性植被為闊葉紅松林,經過長期破壞和大肆開發利用(剩余的原始植被不足10%),導致原生植被已不復存在[8]。紫椴群落平均海拔325 m,郁閉度為0.7,坡向為南坡,土壤肥沃,更有利于草本生長發育。帽兒山林區是原始闊葉紅松林分布區,原始闊葉紅松林遭破壞后,種植人工林,由于種間競爭和更新能力不同形成不同的森林類型,立地質量的差異導致紫椴群落結構的差異[9]。
植物群落是土壤基本屬性的綜合指標,而特定的土壤、氣候和地形條件孕育了不同的植物群落[10-12]。紫椴群落香濃指數表現為:草本層>喬木層>灌木層。由于人為經營活動和火燒干擾,灌木層物種個體均勻性降低,因此,灌木層香濃指數最小。
土壤是植物賴以生存和植被恢復的重要物質基礎,因此土壤理化性質的變化直接影響植被的變化,相反,植被的演替進化可以影響著土壤的發育[13-14]。表3是多樣性指數與環境因子關系,物種多樣性指數中物種豐富度指數與土壤pH值明顯相關,辛普森指數與海拔高度和土壤pH顯著負相關,反映物種多度和物種數量的均勻程度的綜合指數;香濃指數與海拔高度、有機質含量有明顯相關,紫椴對土壤條件要求相對嚴格,多分布于山中、下腹的砂質壤或壤土上,特別適宜生長在土層深厚,排水良好的砂壤土上。紫椴群落均勻度指數與海拔、有機質含量、土壤總氮明顯相關。
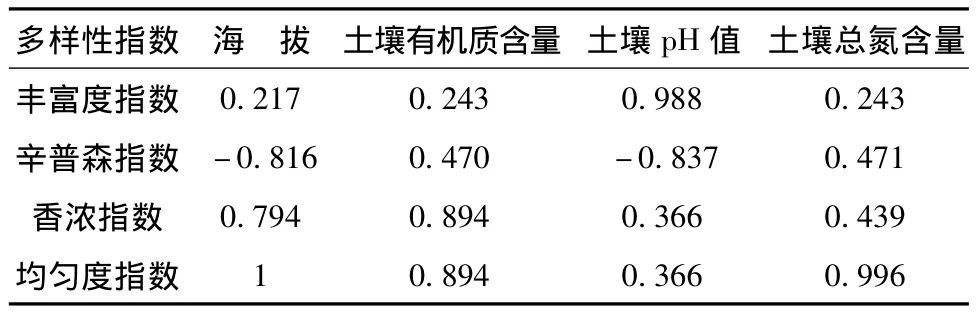
表3 物種多樣性指數與環境因子的相關系數rTab.3 Correlation between species diversity and environmental factors
紫椴是紅松的主要伴生樹種,在不同生長階段往往表現出不同的空間格局[15],Ochiai指數是一個物種看作是另一個物種正常環境的一部分。Dice表示一個物種出現而另一物種也存在的概率。相鄰植物之間的競爭作用影響到植物本身生長發育、種群動態、群落演替和物種多樣性,物種間相互作用與環境差異對植物分布的影響通過種間聯結來反映[16]。帽兒山林區紫椴與五角槭、茶條槭、裂葉榆、暴馬丁香、糠椴之間為正聯結關系,這是它們與所分布地區的生態環境條件長期適應、形成的共憩促進關系。帽兒山紫椴群落Ochaia指數和Dice指數與土壤總氮含相關系數達0.938,0.940,說明紫椴種群分布與土壤含氮狀況息息相關,見表4。
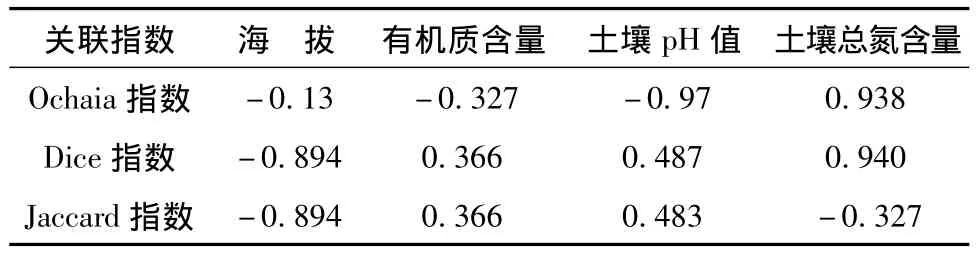
表4 種間相關指數與環境因子的相關系數Tab.4 Correlation between correlation index and environmental factors
群落中植物的生長、發育的各個階段、都不同程度地受到環境和遺傳基因的制約與支配,因而會產生不同程度的聚集和相關關聯,所以測定不同個體在空間聯結的程度,對于研究群落組成與動態及種群間的相互作用都具有重要意義[17-20]。帽兒山林區以人工林為主,人工林天然化發育的理想狀態是目的樹種生長正常,能夠自我更新,物種多樣性豐富,病蟲危害少。處于不同發育階段的紫椴種群與其它地帶性植被存在相似性,紫椴群落物種多樣性指數與種間關聯指數相關性達到了較高水平,這是因為經過前期發育過程中的激烈競爭與遴選,一些物種被淘汰,余下的物種也分別找到了各自合適的位置,彼此間互惠共存,達到了相對穩定階段。
[1]崔 磊,穆立薔.黑龍江省紫椴典型分布區外生菌根形態與解剖特征[J].生態學雜志.2014,22(9):2490-2500.
[2]聶紹荃.紫椴種群生態學研究[M].哈爾濱:東北林業大學出版社,1992.
[3]郭忠玲,馬元丹,鄭金萍.長白山落葉闊葉混交林的物種多樣性、種群空間分布格局及種間關聯性研究[J].應用生態學報,2004,15(11):2013-2018.
[4]張 玲,袁曉穎,張東來.帽兒山落葉松群落主要樹木種群間聯結關系的研究[J].北京林業大學學報,2008,30(4):141-145.
[5]張 玲,袁曉穎,張東來.大、小興安嶺植物區及交錯區物種多樣性比較研究[J].植物研究,2007,27(3):236-360.
[6]王伯蓀.植物群落學[M].北京:高等教育出版社,1987.
[7]鄭超超,伊力塔,張 超,等.浙江江山公益林物種種間關系及CCA 排序[J].生態學報,2015,35(22):1-11.
[8]鄭雪峰,侯照云.帽兒山林區次生林物種多樣性分析[J].林業科技情報.2010,42(4):6-7.
[9]丁 壯.東北林業大學帽兒山實驗林場原始紅松林的破壞與恢復的雛儀[J].植物研究,2013,33(3):379-384.
[10]郭逍宇,張金屯,宮輝力,等.安太堡礦區復墾地植被種間關系及土壤因子分析[J].生物多樣性,2007,15(1):46-52.
[11]宋創業,郭 柯.渾善達克沙地中部丘間低地植物群落分布與土壤環境關系[J].植物生態學報,2007,31(1):40-49.
[12]王順忠,陳桂琛,柏玉平,等.青海湖鳥島地區植物群落物種多樣性與土壤環境因子的關系[J].應用生態學報,2005,16(1):186-188.
[13]趙景學,曲廣鵬,多吉頓珠,等.藏北高寒植被群落物種多樣性與土壤環境因子的關系[J].干旱區資源與環境,2011,25(6):105-108.
[14]張 玲,王承義.大小興安嶺過渡區闊葉紅松林次生演替階段群落多樣性指數與環境因子關系[J].森林工程,2014,30(5):1-5.
[15]吳初平,張 駿,沈愛華,等.千島湖次生林不同演替階段林分空間結構研究[J].植物研究,2015,35(1):16-21.
[16]史作民,劉世榮,程瑞海,等.寶天曼落葉闊葉林種間聯結性研究[J].林業科學,2001,37(2):29-35.
[17]賈呈鑫卓,李帥鋒,蘇建榮,等.擇伐對思茅松天然林喬木種間與種內關系的影響[J].植物生態學報,2014,38(12):1296-1306.
[18]嚴令斌,余德會,李 鶴,等.赤水河清香木群落主要樹種種間關系與生態種組[J].西部林業科學,2015,44(1):69-75.
[19]高金輝,張厚良,陳海波,等.小興安嶺天然闊葉混交林群落多樣性特征[J].林業科技,2014,39(5):30-33.
[20]許林紅,李思廣,蔣云東,等.西南樺、山桂花、肉桂人工林林下物種多樣性和生物量對比研究[J].四川林業科技,2014,35(6):51-55.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
